
N.E. Abdelmeguid
Department of Biological and Environmental Sciences, Faculty of Science, Beirut Arab University, Lebanon
M.H. Mostafa
Department of Biological and Environmental Sciences, Faculty of Science, Beirut Arab University, Lebanon
A.M. Abdel-Moneim
Department of Zoology, Faculty of Science, Alexandria University, Alexandria, Egypt
A.F. Badawi
Department of Biological and Environmental Sciences, Faculty of Science, Beirut Arab University, Lebanon
N.S. Abou Zeinab
Department of Biological and Environmental Sciences, Faculty of Science, Beirut Arab University, Lebanon
International Journal of Cancer Research
Year: 2008 | Volume: 4 | Issue: 3 | Page No.: 81-91







ABSTRACT
The mammary gland cells of Sprague Dawley rats treated with melatonin and/or tamoxifen were analyzed for apoptosis and ultrastructural damage using transmission electron microscopy. The alterations of p53 expression were determined by western blotting. The experiment lasted for 8 weeks where animals were subcutaneously injected with melatonin (0.5 mg/rat daily) and/or tamoxifen (2.5 mg/rat twice a week). It is of considerable interest that the doses used were equivalent to human therapeutic doses reported previously. Obvious features of apoptosis due to melatonin treatment were noticed. These included irregularities of nuclear shapes, clumping and margination of nuclear chromatin and dilatation of the nuclear envelope forming nuclear blebs. Electron micrographs of tamoxifen treated mammary gland showed that there were occasional irregularities in nuclear shape and chromatin distribution. Melatonin/tamoxifen administration resulted in normal cellular and nuclear morphology with normal chromatin distribution. In addition, pyknotic cells increased in frequency, yet this value was less compared to melatonin treated group as well as tamoxifen treated group. On the other hand, melatonin resulted in insignificant increase of p53 expression compared to control while tamoxifen treatment revealed p53 expression similar to control group. In melatonin/tamoxifen treated group, p53 expression was almost confounded greatly to control group. It could be concluded that the synergetic interaction between melatonin and tamoxifen on the expression of p53 is weakly evident on normal mammary gland tissue.
PDF Abstract XML References Citation
How to cite this article
N.E. Abdelmeguid, M.H. Mostafa, A.M. Abdel-Moneim, A.F. Badawi and N.S. Abou Zeinab, 2008. Tamoxifen and Melatonin Differentially Influence Apoptosis of Normal Mammary Gland Cells: Ultrastructural Evidence and p53 Expression. International Journal of Cancer Research, 4: 81-91.
DOI: 10.3923/ijcr.2008.81.91
URL: https://scialert.net/abstract/?doi=ijcr.2008.81.91
DOI: 10.3923/ijcr.2008.81.91
URL: https://scialert.net/abstract/?doi=ijcr.2008.81.91
INTRODUCTION
Selective Estrogen Receptor Modulators (SERM), is a class of breast cancer preventive agents that has been proved to be promising as anticancer agents (DiSaria et al., 1996). SERM exerts its action by binding to the estrogen receptors, thus blocking the proliferative action of estrogen on mammary gland epithelia and can inhibit cancer development (Martindale, 2001). A well characterized member of SERM is tamoxifen, which has been widely used in chemotherapy since it has proved to be the effective front-line of endocrine treatment of breast cancer (Jordan, 1990; Nayfield et al., 1991). Although tamoxifen acts as antiestrogen in breast tissue; yet, the maintenance therapy may be precluded by cumulative toxicity. Like most antineoplastic agents, tamoxifen appears to possess oncogenic potential in laboratory animals (Greaves et al., 1993; Hard et al., 1993) and may be also carcinogenic in man (Fisher et al., 1994; Kedar et al., 1994). Thus the potential undesirable effect of tamoxifen requires modification of its application alone or in combination with other chemopreventive substances which would enable the decrease of its dose and/or its side effects. Thus different developing strategies for the breast cancer chemoprophylaxis, where low non-genotoxic but effective doses of tamoxifen along with other oncostatic agents might yield desired results (Kothari et al., 1997).The pineal indole, melatonin is an oncostatic neurohormone that has been reported by numerous investigators (Blask et al., 1991; Molis et al., 1994; Sanchez-Barcelo et al., 2003; Srinivasan et al., 2008) to be effective in in vivo and in vitro experimental models for human breast cancer. Thus, the combination of melatonin and tamoxifen is indeed very promising.
Cell cycles progression in mammalian cells is regulated by the interactions of cyclins, CDK and CDK inhibitors (Jang et al., 2000). Moreover, gene rearrangement, overexpression and amplification of cyclin D1 and cyclin E have been reported in a number of human as well as of rat mammary tumors (Keyomarsi et al., 1994; Zhang et al., 1994). Whereas, p21 gene has been shown to mediate p53 functions (Wu and Levine, 1997), in such a way overexpression of p21 suppressed the proliferation of tumor cells in vitro as well as tumorigenicity in vivo (Chen et al., 1995). However, the expression in some human cancer cell lines and transplantable murine tumors, tamoxifen (Sionov and Haupt, 1999; Truchet et al., 2000) as well as melatonin antiproliferative effect results from an inhibition of cell cycle kinetics (Shiu et al., 1999; Cos and Sanchez-Barcelo, 2000a; Blask et al., 2002) including the expression of cyclin D1, cyclin E, p21 and p53.
The proposed plan of this research is aimed to assess the apoptic effect both melatonin and tamoxifen alone or in combination on normal mammary gland tissue of female Sprague Dawley rats from utrastructural and molecular point views.
MATERIALS AND METHODS
Animal Selection and Care
At the age of 23 days, 40 prepubertal female Sprague Dawley rats Rattus norvegicus (n = 40), each weighing 35-50 g were used in the present study. Throughout the experiment, all of the rats were adapted to controlled environmental conditions with temperature of 23±2°C, relative humidity 60-70% and artificial regimen of light: dark-10:14 h (light on 7:00 am). Rats were housed in separate large cages (4 rats/cage) where they are provided with standard food diet and drinking tap water ad libitum. The cages were cleaned every alternate day.
Chemicals Used
Melatonin, 5-methoxy-N-acetyltryptamine was obtained as off-white powder, purchased from Fluka Chemic GmbH CH-9471 Buchs, 081/7552511. Sigma-Aldrich, Pf. D-89552 Steinheim, 07329/970. Tamoxifen was purchased from SIGMA-Aldrich (Steinheim, Germany). CAS Registry No. 105450-29-1.
Experimental and Control Groups
The animals were divided into two major groups:
Group 1: Control group (G 1): Animals of this group were further subdivided into two subgroups as follows:
| • | G 1a (n = 8): Female rats of this group did not receive any kind of chemical treatment thus they were used as controls. |
| • | G 1b (n = 8): Female rats of this group received daily subcutaneous injections of the first vehicle (30% of ethanol) at a dose of 0.1 mL and another subcutaneous injection of the second vehicle (sesame oil/absolute ethanol; 9:1 ratio) at a dose of 0.1 mL twice a week. Such animals were also used as controls. |
Group 2: Experimental group (G 2): Animals of this group were further subdivided into three subgroups as follows:
| • | G 2a (melatonin treated group) (n = 8): Each rat of this group was injected subcutaneously in the dorsal interscapular area with freshly prepared melatonin at a dose of 0.5 mg, dissolved in 0.1 mL of 30% ethanol. Such injections received by the rats were given daily in the late-afternoon, (i.e., between 5:00 pm and 8:00 am) (Blask et al., 1991). |
| • | G 2b (tamoxifen treated group) (n = 8): Each rat of this group was injected subcutaneously in the dorsal interscapular area with tamoxifen at a dose of 2.5 mg per rat where tamoxifen was dissolved in 0.1 mL of (1:9) absolute ethanol/sesame oil, twice a week (Kubatka et al., 2001). |
| • | G 2c (melatonin/tamoxifen treated group) (n = 8): Each rat of this group was injected subcutaneously in the dorsal interscapular area with melatonin and tamoxifen at dose levels similar to those previously mentioned in group (2a) and group (2b). |
It is of considerable interest that doses used were equivalent to human therapeutic doses reported previously. Three animals were sacrificed after 5 weeks of treatment (i.e., such animals were still in their pre-maturation stage). The remaining animals were sacrificed at the age of 83 days i.e., after 8 weeks of treatment, where such animals have already reached post-maturation stage of their development. Mammary glands from both experimental as well as control groups were dissected. Small pieces gland tissues of postmature rats were fixed in 4F1G for ultrastructural studies while pieces of mammary tissues of both pre- and post-mature animals were immediately frozen in liquid nitrogen and stored at -70°C for subsequent determination of p53 protein expression.
Preparation of Materials for Electron Microscopy
Small blocks from the mammary gland tissues about 1 mm3 of different groups were removed and dropped as soon as possible into 4F1G fixative, buffered with 0.1 M phosphate buffer, pH = 7.4 at 4°C for around 1 h. After two rinses into the buffer (for a period of 4 h or overnight), fixed samples were post-fixed in 1% buffered OsO4 for 1-2 h at 4°C. Then the tissue pieces were washed twice in buffer for 30 min. The specimens were then dehydrated in ascending grades of ethanol concentration. Tissues were then treated with propylene oxide and embedded in a mixture of 1:1 of Epon-Araldite. Specimens were embedded in pre-dried gelatin capsule (dried in the oven at 37°C for 1 h before use). Polymerization was done in the oven at 65°C for 24 h. Ultrathin sections obtained from such blocks were cut on a LKB ultratome with a glass knife, mounted on copper grids, double stained with uranyl acetate and lead citrate and investigated on a JEOL 100CX transmission electron microscope.
Protein Extraction for Electrophoresis and Western Blotting Studies
Total protein concentration was assayed by the method of Bradford (1976), which utilizes the change in absorbance of Coomassie Brilliant Blue G-250 at 595 nm in acidic solution when bound to protein. The isolated and extracted proteins from all treatments under study were analyzed by SDS-polyacrylamide gel electrophoresis (SDS-PAGE) according to Laemmili (1970). Western blotting was carried out according to Towbin et al. (1979). Isolated proteins were subjected to SDS-PAGE (10%), using EC 120 Mini Vertical Gel System (EC 250-90). After electrophoresis, the proteins in the gel were transferred by electroblotting, using Western blotting transfer buffer (100 volts, 1 h, 4°C) to a PVDF membrane pretreated with methanol. After brief rinse in 1x TBS, the membranes were then blocked using Western blocking buffer 5% non-fat milk in 1x TBS for 1-2 h at room temperature with gentle continuous agitation. The primary antibody (CHEMICON International Company) was diluted in 0.5% blocking buffer (1:1000) and incubated at room temperature for 3-12 h, following which the blot was washed with TBST buffer (4x15 min). After the last wash, the membrane was rinsed briefly for 5 min in 1x TBS after which the membrane was incubated with the conjugate antibody (Horse Raddish Peroxidase labelled) at a dilution of 1:2000 in 0.5% blocking buffer for 1 h at room temperature followed by washing with TBST (4x15 min) and for 5 min with 1x TBS buffer. Excess TBS was blotted off immediately prior to detection by TMB/M peroxidase substrate for blotting, CHEMICON International Company, Cat.No.ES002-500ML. The band intensity of p53 protein expression among experimental groups as well as control groups was measured using Scion Image Frame Grabber Status Program.
Statistical Analysis
The immunoblots were independently repeated three times. All blots presented are representative of at least three separate samples of each group. All quantitative data are expressed as mean±SD. Comparisons between two groups were analyzed using Student`s t-test and values of p<0.05 were considered statistically significant.
RESULTS
TEM Observations
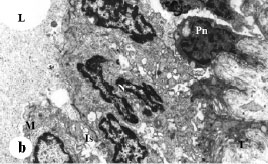
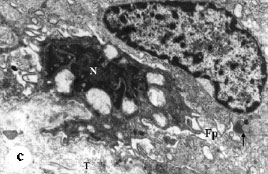
The mammary gland cells in the control group and in animals treated with melatonin and/or tamoxifen were analyzed for apoptosis and ultrastructural damage in postmature animals. Mammary gland of control animals had normal cell architecture; some cells were observed occasionally in apoptosis (Fig. 1). Obvious features of apoptosis due to melatonin treatment were commonly noticed. These included irregularities of nuclear shapes, clumping and margination of nuclear chromatin and dilatation of the nuclear envelope forming nuclear blebs (Fig. 2). Increased pyknotic frequency was also noticed. Electron micrographs of tamoxifen treated mammary gland showed that there were occasional irregularities in nuclear shape and chromatin distribution (Fig. 3). Nuclear envelope was almost normal in some cells while it was irregular in few others forming evident nuclear blebs. Pyknotic frequency increased, yet this value reduced in comparison to melatonin treated group. Melatonin/tamoxifen administration resulted in normal cellular and nuclear morphology with normal chromatin distribution (Fig. 4a). Few cells exhibited pleomorphism of their shape. Nuclear abnormalities including bizarre nuclear profiles, Sezary`s nuclei, nuclear pockets, margination of the heterochromatin, terminalization of nucleoli and blebbing of the nuclear envelope were also occasionally observed (Fig. 4a-c). Pyknotic cells increased in frequency, yet this value was lessened compared to melatonin treated group as well as tamoxifen treated group. Electron microscopical preparations of control group (G1b) (received vehicle only) did not show great abnormalities from those of control group (G1a).
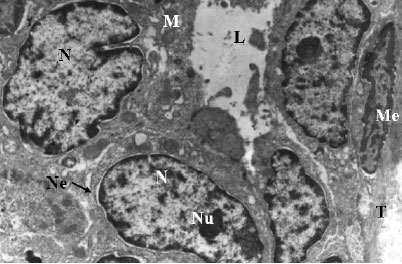 | |
| Fig. 1: | Electron micrograph of control rat`s mammary gland showing nuclei (N) with marginated heterochromatin appearing as thin dense line along the nuclear envelope (Ne). Nucleoli (Nu) are obvious in some nuclei. Note also few spherical mitochondria (M), rough endoplasmic reticulum dispersed within the cytoplasm. Oval shape myoepithelial cells (Me) with heterochromatic nuclei and moderately electron dense cytoplasm can be seen with thick bundles of tonofilaments (T). Short blunt microvilli project toward the lumen (L). Notice the narrow intercellular spaces. X3000 |
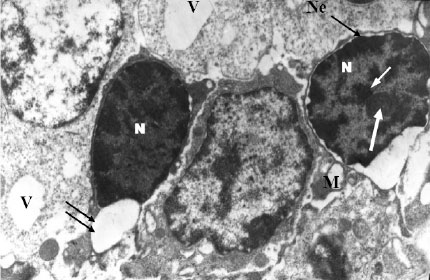 | |
| Fig. 2: | Electron micrograph of melatonin treated rat`s mammary gland showing nuclei (N) greatly endowed with heterochromatin aggregated in the form of great clumps along the margin of nuclear envelope (Ne). Note the intranuclear amorphous dense body (arrow) associated with heterochromatin and pseudonucleoli (or karyosomes) (thick arrow). Nuclear envelope (Ne) shows dilatation at certain sites forming large blebs (double arrow). Few vacuolated mitochondria (M) are scattered within the cytoplasm. Large vacuoles (V) are found within the cytoplasm. X7500 |
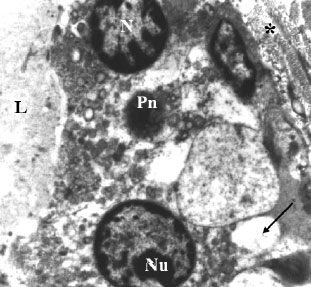 | |
| Fig. 3: | Electron micrograph of tamoxifen treated rat`s mammary gland. Normal acinar architecture is poorly differentiated. The lumen (L) lacks microvilli. Lateral borders are obscure. Nuclei (N) of most cells are of different sizes and highly heterochromatic. Heterochromatin accumulated at certain sites along the nuclear envelope and scattered within the nuclear matrix. Note some nuclei appear poorly endowed with heterochromatin within their nucleoplasm. Nucleolus (Nu) is prominent and terminalized. Pyknotic nucleus (Pn) with obscure nucleolus can be seen. Highly vacuolized cytoplasm of cells occurs with high frequency (arrow). High degree of fibrosis (*) is observed at basement membrane. X5000 |
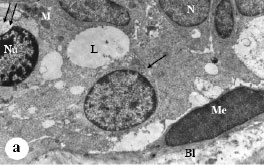
| Fig. 4: | Electron micrograph of melatonin/tamoxifen treated rat`s mammary gland, (a) Almost normal acinar architecture can be seen. Lumen (L) lacks microvilli. Lateral cellular borders are closely packed (arrow). Nuclei (N) appear highly euchromatic spherical or oval. Few densely stained mitochondria (M) appear in the cytoplasm. Note also, pleomorphic heterochromatic nucleus with terminalized nucleolus (Nu), dilated nuclear envelope forming large bleb (double arrow). Spindle shape myoepithelial cell (Me) attached to the basal lamina (Bl) by hemidesmosomes can also be observed. X5000, (b) Note the irregular shape of mammary gland cells with highly heterochromatic nuclei, N: Nucleus bisected by deep invagination giving bizarre profile, Pn: pyknotic nucleus, M: mitochondria (M) with evident vacuolated matrices, T: tonofilaments, Is: intercelluar space. X5000, (c) Notice the ceribriform nucleus known as Sezary`s nucleus (N). The cytoplasm possesses numerous vacuoles. Secondary lysosomes are frequent within the cytoplasm (arrow). Filopodia (Fp) and tonofilaments (T) with increased fibrosis are common within intercellular spaces. X7500 |
Detection of p53 Protein Expression Level by Immunoblotting
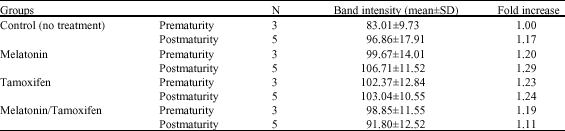
In the present study, Western blot analysis was carried out to detect the level of p53 protein expression using p53 monclonal antibody. It was revealed that p53 protein levels in rats` normal (control group) mammary glands at their premature and postmature stages indicated normal p53 protein expressed in normal mammary gland tissue (Table 1, Fig. 5 Lanes 2-3). However, the expression of p53 protein levels increased in melatonin treated rat mammary glands at their postmature with respect to their corresponding at the premature stages. Yet, such increase of p53 expression, whether in animals at their premature or postmature stages, was found to be insignificant (p>0.05) when compared to their corresponding control group (Table 1, Fig. 5 Lanes 4-5). On the other hand, p53 protein level in tamoxifen treated rat mammary glands exhibited slight increase that was almost negligible when compared to control group. Taking into consideration that p53 protein expression among animals of this group, did not show any changes whether in their premature or postmature stages (Table 1, Fig. 5 Lanes 6-7). In addition, further Western blot analysis of p53 protein level demonstrated that in melatonin/tamoxifen treated group, p53 protein expression decreased insignificantly (p>0.05) in comparison to their corresponding in melatonin as well as in tamoxifen treated groups. However, p53 protein levels among animals of this group, whether in their premature and postmature stages, illustrated almost similar expression to those recorded in control group (Table 1, Fig. 5 Lanes 8-9).
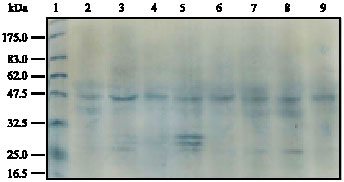 | |
| Fig. 5: | Immunoblotting analysis of p53 proteins extracted from rats` 4th abdominal mammary gland. Lane 1 represents standard protein molecular weight. Lanes 2 and 3 represent protein of control mammary glands. Lanes 4 and 5 represents protein of melatonin treated mammary gland. Lanes 6 and 7 represents protein of tamoxifen treated mammary gland. Lanes 8 and 9 represents protein of melatonin/tamoxifen treated mammary gland |
| Table 1: | Differences in band intensity of immunoblot staining of p53 proteins among different experimental and control groups |
 | |
Insignificant changes of p53 protein expression in all groups | |
DISCUSSION
During the course of investigation, a dose of 28 mg of melatonin consumed at an average daily 0.5 mg per rat failed to result in any significant variation or changes in the expression of p53 protein in the mammary gland of animals belonging to this group, when compared to control group. Many researchers have conducted experiments in evaluating the antiproliferative effect of melatonin on mammary tumor cells more than normal cells and more specifically its action on the expression of p53 protein level. In their study, Cos and Sanchez-Barcelo (2000a, b) showed that physiological melatonin-induced increases in p53 expression which can trigger apoptosis in cancer cells. Worth mentioning that relative increase expression of p53 protein could be correlated to an insignificant increase in the cellular proliferation manifested by insignificant (p>0.05) increase in the epithelial layer thickness of mammary gland duct (unpublished data). However, most of the cells were altered and exhibited features of apoptosis, in such a way that the normal mammary gland architecture exhibited sort of variation. Yet, it is worthy to mention that pyknotic cells were observed in ultrastructural preparations of melatonin treated mammary gland sections. Pyknosis may lead to apoptosis that is mediated by the action of p53 that induce cellular arrest. Thus, our findings tend to show that melatonin could exert hazardous effect on normal mammary gland tissue that is not cancerous in nature. She et al. (2004) suggested that H22 mouse hepatoma cells receiving 1x10(-6) mol L-1 of melatonin treatment exhibited significant elevation of p53 protein expression. Thus, they concluded that melatonin exerted its antiproliferative effect by inhibiting the proliferation of H22 cells by arrest and apoptosis and the mechanism interfere with increasing p53 protein expression. Another experiment was conducted by Mediavilla et al. (1999) who demonstrated that the antiproliferative effect of 1 nM of melatonin administrated to human MCF-7 cells was evaluated as it induces the expression of some proteins involved in the control of cell cycle including p53 protein. In addition, MCF-7 proliferation was inhibited due to the action of melatonin administration that induces cell cycle arrest dependent on the increased expression of p53 protein. Cos and Sanchez-Barcelo (2000a) demonstrated that melatonin upregulates the expression of p53 and p21/Waf1 proteins in MCF-7 cells in vitro. They correlated this to an important mechanism by which melatonin causes a cell cycle transition delay at the G1-S interface in this cell line. Moreover, Bast et al. (2000) suggested that melatonin-treated MCF-7 cells resulted in accumulation of cells in G1 which provided a greater opportunity for them to enter the non-dividing state (G0) which is typical of terminally-differentiated cells. Kothari et al. (1997) suggested that melatonin`s antiproliferative effects on tumor cells may be due in part to its ability to shift the balance from proliferation to differentiation.
On the other hand, it was found that total intake of approximately 40 mg of tamoxifen during 56 days (2.5 mg per rat twice a week) was insufficient to produce significant elevation or reduction in the expression of p53 protein level in rat mammary gland cells when compared to the corresponding control group. Such results were in great correlation with the ultrastructural observation of tamoxifen treated mammary gland that illustrates almost normalization of the mammary gland architecture with increased cellular proliferation (unpublished data). Most proliferated cells attained almost normal morphology. Thus, apoptotic features were evident to a lesser extent; when compared to those of melatonin-treated group. Pyknotic cell frequency increased yet such value was less than those detected in melatonin treated groups. Thus, we may refer this pyknosis to the side effects of tamoxifen treatment on normal mammary gland cells especially by inducing apoptosis to a lesser extent. Present results are not consistent to those reported by Wright et al. (2005) who suggested that 500 ng mL-1 of tamoxifen administrated to human ovarian surface epithelial cells (OSE) inhibited cellular proliferation where p53 protein level was elevated 2-3 fold in its expression. Similar studies were conducted by Kulesz-Martin et al. (1994) and Ljungman et al. (2001) who suggested the anticancerous effect of tamoxifen on mammary gland tumor cells that was evaluated clearly by the significant elevation of p53 protein expression. Fisher et al. (1994) and Thiantanawat et al. (2003) documented that the mechanisms and signaling pathways regulating cell cycle and induction of apoptosis upon treating breast cancer cells with aromatase inhibitors in comparison with antiestrogens. Zhang et al. (1999) and Hengartner (2000) showed that tamoxifen-induced apoptosis in MCF-7 cells involves down-regulation of Bcl-2 but not Bax. They suggested that aromatase inhibitors and antiestrogens such as tamoxifen (Tam) inhibit the proliferation of MCF-7Ca breast cancer cells by inducing cell cycle arrest in the G0-G1 phase and this is coupled with increased apoptosis.
During the course of investigation, the combined treatment of melatonin and tamoxifen did not show also any significant changes in p53 protein expression. Yet, it is worthy to mention that p53 protein expression levels were almost confounded greatly to control group. Such results could be appreciated at the ultrastructural level where combined treatment revealed obvious signs of protection manifested by the great normalization of the mammary gland architecture when compared to their corresponding in control groups. Contradicting our results, Kothari et al. (1997) demonstrated that the combined treatment of melatonin and tamoxifen resulted in reducing the tumor incidence in MNU-treated rat mammary gland. They believed that melatonin synergistically enhances the antiproliferative effect of tamoxifen. Similarly, Massion et al. (1995) suggested that both tamoxifen and melatonin treatment synergistically appear to affect the same cell cycle phase and specifically cause G1 to S phase transition delay due to the synergetic increase of p53 expression that induced cellular arrest in MNU-induced rat mammary gland carcinomas. However, in the present study, it could be concluded that the synergetic interaction between melatonin and tamoxifen on the expression of p53 is weakly evident on normal mammary gland tissue.
REFERENCES
- Blask, D.E., D.B. Pelletier and S.M. Hill, 1991. Pineal melatonin inhibition of tumor promotion in the N-nitroso-N-methylurea model of mammary carcinogenesis: Potential involvement of antiestrogenic mechanisms in vivo. J. Cancer Res. Clin. Oncol., 117: 526-532.
PubMed - Blask, D.E., R.T. Dauchy, L.A. Sauer, J.A. Krause and G.C. Brainard, 2002. Light during darkness, melatonin suppression and cancer progression. Neuro. Endocrinol. Lett., 2: 52-56.
PubMed - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Chen, Y.Q., S.C. Cipriano, J.M. Arenkiel and F.R. Miller, 1995. Tumor suppression by p21WAF1. Cancer Res., 55: 4536-4539.
PubMed - Cos, S. and E.J. Sanchez-Barcelo, 2000. Melatonin, experimental basis for a possible application in breast cancer prevention and treatment. Histol. Histopathol., 15: 637-647.
PubMed - DiSaria, P.J., E.A. Grosen and T. Kurosaki, 1996. Hormone replacement therapy in breast cancer survivors: A cohort study. Am. J. Obstet. Gynecol., 174: 1494-1498.
PubMedDirect Link - Fisher, B., J.P. Costantino, C.K. Redmond, E.R. Fisher, D.L. Wickerham and W.M. Cronin, 1994. Endometrial cancer in tamoxifen-treated breast cancer patients: Findings from the National Surgical Adjuvant Breast and Bowel Project (NSABP) B-14. J. Natl. Cancer Inst., 86: 527-537.
CrossRefPubMedDirect Link - Greaves, P., R. Goonetilleke, G. Nunn, J. Topham and T. Orton, 1993. Two years carcinogenicity study of tamoxifen in Alderly Park Wistar derived rats. Cancer Res., 53: 3919-3924.
PubMed - Hard, G.C., M.J. Iatropoulos, K. Jordan, L. Radil, O.P. Kaltenberg, A.R. Imondi and G.M. Williams, 1993. Major differences in the hepatocarcinogenicity and DNA adduct forming ability between tormefene and tamoxifen in female CnCD(BR) rat. Cancer Res., 53: 4534-4541.
PubMed - Hengartner, M.O., 2000. The biochemistry of apoptosis. Nature, 407: 770-776.
CrossRefPubMedDirect Link - Jordan, V.C., 1990. Long-term adjuvant tamoxifen therapy for breast cancer. Breast Cancer Res. Treat., 15: 125-136.
PubMed - Jang, T.J., M.S. Kang, H. Kim, D.H. Kim, J.I. Lee and J.R. Kim, 2000. Increased expression of Cyclin D1, Cyclin E and p21Cip1 associated with decreased expression of p27Kip1 in chemically induced rat mammary carcinogenesis. Jap. J. Cancer Res., 91: 1222-1232.
PubMed - Keyomarsi, K., N. O’Leary, G. Molnar, E. Lees, H.J. Fingert and A.B. Pardee, 1994. Cyclin E, a potential prognostic marker for breast cancer. Cancer Res., 54: 380-385.
PubMed - Kubatka, P., B. Bojkova and K. Mocikova-Kalicka, 2001. Effect of tamoxifen and melatonin on mammary gland cancer induced by N-methyl-N-nitrosourea and by 7,12-dimethylbenz(a)-anthracene, respectively, in female Sprague-Dawley rats. Folia. Biol. (Praha), 47: 5-10.
PubMed - Kulesz-Martin, M.F., B. Lisafeld, H. Huang, N.D. Kisiel and L. Lee, 1994. Endogenous p53 protein generated from wild-type alternatively spliced p53 RNA in mouse epidermal cells. Mol. Cell Biol., 14: 1698-1708.
PubMed - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Ljungman, M., H.M. O’Hagan and M.T. Paulsen, 2001. Induction of Ser15 and Lys382 modifications of p53 by blockage of transcription elongation. Oncogene, 20: 5964-5971.
PubMed - Molis, T.M., L.L. Spriggs and S.M. Hill, 1994. Modulation of estrogen receptor mRNA expression by melatonin in MCF-7 human breast cancer cells. Mol. Endocrinol., 8: 1681-1690.
PubMedDirect Link - Nayfield, S.G., J.E. Karp, L.G. Ford, F.F. Dorr and B.S. Kramer, 1991. Potential role of tamoxifen in prevention of breast cancer. J. Natl. Cancer Inst., 83: 1450-1459.
PubMed - Sanchez-Barcelo, E.J., S. Cos, R. Fernandez and M.D. Mediavilla, 2003. Melatonin and mammary cancer: A short review. Endocr. Relat. Cancer, 10: 153-159.
PubMed - She, M.H., B.B. Chen, X.M. Wang and S.S. He, 2004. P53-dependent antiproliferation and apoptosis of H22 cell induced by melatonin. Ai Zheng., 23: 803-807.
PubMed - Shiu, S.Y.W., L. Li, J.N. Xu, C.S. Pang, J.T.Y. Wong and S.F. Pang, 1999. Melatonin-induced inhibition of proliferation and G1/S cell cycle transition delay of human choriocarcioma JAr Cells: Possible Involvement of MT2 (MEL1b) Receptor. J. Pineal Res., 27: 183-192.
PubMed - Sionov, R.V. and Y. Haupt, 1999. The cellular response to p53: The decision between life and death. Oncogene, 18: 6145-6157.
PubMed - Srinivasan, V., D.W. Spence, S.R. Pandi-Perumal, I. Trakht, A.I. Esquifino, D.P. Cardinali and G.J. Maestroni, 2008. Melatonin, environmental light, and breast cancer. Breast Cancer Res. Treat., 108: 339-350.
PubMed - Thiantanawat, A., B.J. Long and A.M. Brodie, 2003. Signaling pathways of apoptosis activated by aromatase inhibitors and antiestrogens. Cancer Res., 63: 8037-8050.
PubMed - Towbin, H., T. Staehelin and J. Gordon, 1979. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA., 76: 4350-4354.
PubMedDirect Link - Wright, J.W., R.L. Stouffer and K.D. Rodland, 2005. High-dose estrogen and clinical selective estrogen receptor modulators induce growth arrest, p21 and p53 in primate ovarian surface epithelial cells. J. Clin. Endocrinol. Metab., 90: 3688-3695.
PubMed - Wu, L. and A.J. Levine, 1997. Differential regulation of the p21/WAF-1 and mdm2 genes after high-dose UV irradiation: p53-dependent and p53-independent regulation of the mdm2 gene. Mol. Med., 3: 441-451.
PubMed - Zhang, S.Y., J. Caamano, F. Cooper, X. Guo and A.J. Klein-Szanto, 1994. Immunohistochemistry of cyclin D1 in human breast cancer. Am. J. Clin. Pathol., 102: 695-698.
PubMed - Zhang, S., D.J. Hunter, S.E. Hankinson, E.L. Giovannucci, B.A. Rosner, G.A. Colditz, F.E. Speizer and W.C. Willet, 1999. A prospective study of folate intake, alcohol and risk of breast cancer. JAMA, 281: 1632-1637.
PubMed