
K. Nirmala
Food and Drug Toxicology Research Centre, National Institute of Nutrition (ICMR) PO-Jamai Osmania, Hyderabad-500 007, India
T. Prasanna Krishna
Food and Drug Toxicology Research Centre, National Institute of Nutrition (ICMR) PO-Jamai Osmania, Hyderabad-500 007, India
K. Polasa
Food and Drug Toxicology Research Centre, National Institute of Nutrition (ICMR) PO-Jamai Osmania, Hyderabad-500 007, India
International Journal of Cancer Research
Year: 2007 | Volume: 3 | Issue: 3 | Page No.: 134-142







ABSTRACT
A link between diet and health has been appreciated by many cultures for centuries; however, an association between diet and cancer has been documented only in recent decades. The diet is likely the major source of human exposure to environmental carcinogens/mutagens and anticarcinogens/antimutagens. Spices and condiments that are important components of our diet are also used in traditional medicine to treat variety of ailments. Ginger has been used in Ayurveda to alleviate inflammation, pain associated with rheumatoid arthritis, osteoarthritis and muscular discomfort. Literature evidence suggests that some of the active constituents present in ginger show anticancer activity by inhibiting tumorigenicity in animal model following topical application/feeding. Agents having genotoxic property exhibit antimutagenic, antioxidant and anti-inflammatory properties. It was shown in an in vitro experiment using single cell gel electrophoresis that ginger could inhibit benzo(a)pyrene, a well known carcinogen, induced cell damage, suggesting that ginger has antimutagenic potential. In order to investigate if this effect could be present even under in vivo conditions, a study was planned to investigate antimutagenic potential of ginger. Ginger powder was incorporated in the diet and fed to NIN male Wistar rats. At the end of one month of ginger feeding the rats were injected with 5 mg of benzo(a)pyrene by intraperitoneal route(ip). Twenty-four hours urine was collected before and after carcinogen exposure. The urine samples were analyzed for urinary mutagens by Ames test. The results indicated the antimutagenic potential of ginger under in vivo conditions using the strains TA 98 and TA 100 strongly suggesting that regular intake of ginger through diet is likely to confer protective effect against carcinogen induced mutation events. Since dietary components are subjected to cooking before consumption, an in vitro experiment was designed to assess the antimutagenic potential of ginger subjected to heat treatment. The antimutagenic activity of ginger was retained even after boiling and frying indicating that ginger consumed through diet can confer protection against mutational events.
PDF Abstract XML References Citation
How to cite this article
K. Nirmala, T. Prasanna Krishna and K. Polasa, 2007. In vivo Antimutagenic Potential of Ginger on Formation and Excretion of Urinary Mutagens in Rats. International Journal of Cancer Research, 3: 134-142.
DOI: 10.3923/ijcr.2007.134.142
URL: https://scialert.net/abstract/?doi=ijcr.2007.134.142
DOI: 10.3923/ijcr.2007.134.142
URL: https://scialert.net/abstract/?doi=ijcr.2007.134.142
INTRODUCTION
The association between the nutrients and non-nutrients of diet and cancer was reported initially by Doll and Peto (1981). It has been suggested that intake of antimutagens and anticarcinogens, particularly those present as natural components of diet, can be one of the effective way of preventing human cancer and other mutation based degenerative diseases (Wattenberg, 1992) Antimutagenic substances occur naturally in all food categories.
Thus the association between occurrence of cancer and diet is strong. Cancer chemopreventive potential of naturally occurring phytochemicals is gaining lot of importance and interest. Although it may be impossible to avoid exposure to xenobiotics and their toxic effects, it may be possible to modify their effects by consuming diet rich in protective phytonutrients that will enhance the cellular protective mechanism and reverse the effects of xenobiotics. Antigenotoxicants have antimutagenic property and inhibit or deactivate the mutagens/carcinogens. It has been suggested that chemoprevention should be considered as an inexpensive, easily applicable approach to cancer control. Different classes of antigenotoxicants have various mechanisms of action. Antigenotoxic activities of various phytochemicals have great potential in the fight against human cancer (Park and Pezzuto, 2002).
Spices and condiments, which are part of the Indian diet, have chemical constituents that have antioxidant, antimutagenic and anticarcinogenic properties (Krishnaswamy and Polasa, 2001). There are several short-term tests for mutagenicity that are suitable for studying mutagenic or antimutagenic potential of specific components. In an in vitro experiment on ginger it was shown that ginger has potential to inhibit carcinogen induced cell damage by benzo(a)pyrene as measured by comet assay (Nirmala et al., 2007).
This study was planned to evaluate the in vivo antimutagenic potential of ginger by quantifying the urinary mutagens of benzo(a)pyrene exposed rats using the salmonella microsome assay. In a study conducted on alliums and onion, when subjected to heat treatment, it was observed that both raw and cooked form of onion and garlic retained their antimutagenic potential (Polasa et al., 2006). Simultaneously an in vitro experiment was also done with dried and fresh forms of ginger after subjecting them to normal cooking conditions to assess if the antimutagenic potential of ginger is retained even after thermal processing. Benzo(a)pyrene is an ubiquitously present compound in the environment and belongs to polycyclic aromatic hydrocarbon. It is metabolized mainly by the Cytochrome P450 (Cyt P450) dependent AHH into various derivatives including electrophilic epoxides (Gelboin, 1980). These epoxides are thought to play a major role in determining the mutagenic and carcinogenic effect of the parent compound in various species and tissues (Brauze et al., 1991). The testing of urine from treated animals can provide information on the in vivo metabolism and therefore is considered to be good biomonitoring method. The test has also been shown to be very useful in correlating in vitro bacterial mutagenesis and in vivo carcinogenesis in animals (Deflora, 1998; Ikkeny et al., 1999). Therefore, ginger was tested for its in vivo antimutagenic potential in this study. Ginger at 0.5, 1 and 5% were incorporated in diet and fed to rats for one month. Antimutagenic potential of ginger feeding was tested by using urine concentrates belonging to B(a)P exposed and not exposed group by Ames test.
MATERIALS AND METHODS
Bacterial Cultures
Salmonella typhimurium strains TA98 and TA100 were procured from Prof. B.N. Ames, University of California. They were sub cultured in nutrient broth, were dispensed in small vials and frozen in the presence of dimethyl sulfoxide (DMSO) and kept at -80°C. The bacterial cultures were incubated in fresh nutrient broth and grown for 16 h at 37°C before the experiment.
Chemicals
Nutrient broth and agar were purchased from Himedia, Hyderabad and benzo(a)pyrene from Sigma Chemical Company. All other chemicals were of analytical reagent quality.
Preparation of Experimental Diets
Ginger was purchased from local market, peeled, washed, coarsely minced, air dried and pulverized with a blender to a fine powder. This was added to the stock diet and thoroughly mixed so as to get diets of three compositions containing 0.5, 1 and 5% ginger. The stock diet contained wheat flour 15%, roasted Bengal gram flour 58%, groundnut flour 10%, skimmed milk powder 5%, casein 4%, refined oil 4%, salt mixture 4% and vitamin mixture 0.2%. The nutritive value of the diets fed to control and experimental groups were identical.
In vivo Experiment
Study Design
The procurement of animals for experiment was undertaken by following the Institutes Animal Ethical Clearance Committee. Inbred male NIN wistar rats aged about 8-10 weeks were taken. They were randomly divided into four groups containing 6 animals in each group. The first group was taken as control and received standard stock diet and the other three groups were fed with 0.5, 1 and 5% ginger through diet, respectively (Table 1). The rats were maintained at 22±1.2°C with 50-55% relative humidity and a 12 h light/dark cycle. Food and water were given adlib. The weekly food intake of the animals were recorded at the beginning and end of the experiment.
After one month of ginger feeding, the animals were given intraperitoneal injection of 5 mg B(a)P as a single dose in ground nut oil. Twenty-four hour urine was collected from each rat, using metabolic cage on two occasions from the entire animals prior and after carcinogen exposure.
Concentration of Urinary Mutagens
There was no change in pH of the urine (6.5) at the time when samples were deported for concentration. The urine samples were filtered passed through XAD-2 column and the mutagens were eluted with acetone. The eluent was evaporated and stored in the refrigerator at 4°C till analysis.1.5 mL of DMSO was added to each sample before setting up the antimutagenicity assay.
In vitro Experiment
Ginger paste and powder was used in the experiment and the number of groups categorized were 1) unboiled 2) boiled 3) unfried and 4) fried
Preparation of Boiled/unboiled Fresh Ginger Paste and Powder
Three grams of ginger paste and one gram of ginger powder was taken in 2 sets of tubes containing water and vortexed for 5 mts. One set of tubes was subjected to boiling for 5 mts and cooled. All the tubes were centrifuged at 4000 RPM for 20 mts. The supernatants were used for the Ames test. To study the antimutagenicity of ginger, ginger paste was used at 1, 2 and 3 mg concentration and ginger powder was used at 0.5, 1 and 1.5 mg concentration for boiled/unboiled/fried/unfried, respectively.
Preparation of Frled/unfried Fresh Ginger Paste and Powder
Three grams of ginger paste and one gram of ginger powder was taken in two sets of tubes containing 10 mL of corn oil. Five mL of DMSO was added and vortexed for 5 mts. The DMSO layer was used for the Ames test.
| Table 1: | Ginger (G) intake in different groups |
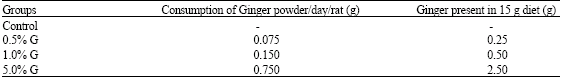 | |
| Rats belonging to Control and Experimental groups consumed 15 g of diet per day/animal | |
The Ames Test
The main objective of this study was to evaluate the in vivo antimutagenic potential of ginger by studying its effect on two histidine requiring strains of Salmonella typhimurium namely TA 98 and TA 100 in the absence and presence of a liver metabolizing system (Maron and Ames, 1983). Since dietary components are subjected to cooking before consumption, an in vitro study was planned to assess the antimutagenic potential of fresh and dried forms of ginger under normal cooking conditions using both the tester strains mentioned above
Benzo(a)pyrene concentration of 2 and 5 μg was used as the mutagen. Ginger paste was used at concentrations of 1, 2 and 3 mg, respectively and ginger powder was used at concentrations of 0.5, 1.0 and 1.5 mg, respectively.
Basic Procedure
Nutrient broth was inoculated with Salmonella strains of TA 98 and TA 100 and incubated overnight. Bacterial culture, urine extracts and S9 mix were mixed with top agar and then added to minimal agar plates. The plates were incubated, kept inverted in incubator at 37°C for 48 h. The number of revertant colonies was counted and the means were calculated for the individual plate counts for each dose. Statistical analyses of the counts were carried out and the results for mutagenicity were assessed.
Statistical Analysis
The data on in vivo experiment was analyzed by the analysis of variance (ANOVA). Mean values were tested for significance by Duncan’s multiple range test. The analysis of data for the in vitro experiment was performed by Descriptive statistics (i.e., means, standard deviation) and the calculation was done for each category of the groups. Between group differences were tested by non-parametric Kruskal Wallis test in view of heterogeneity of variance. SPSS 14.5 window version was used (Middle Brooks, 1977).
RESULTS
In vivo Experiment
There were no significant differences in weight gain between control and experimental animals at the end of the feeding period indicating that ginger-containing diet did not alter the intake of diet (Table 2).
The TA98 and TA100 reversion frequency in our lab ranged from 15-25 (-S9), 20-30 (+S9) for TA98. For TA100 it was 110-120(-S9) and 120-150 (+S9). These values were in acceptable range as described by Maron and Ames (1983). The frequency of revertants in the basal samples in the absence of B(a)P, with both the two tester strains TA98 and TA100 was found to be similar in control and ginger fed groups, both with or without S9 mixture. Ginger feeding by itself did not produce any adverse effect. Elevated No. of revertants (p<0.001) were observed in both TA98 and TA100 strains of salmonella typhimurium in urine extracts of carcinogen exposed rats. Similar observations were made in the presence of metabolic activation system (S9) (Table 3 and 4). The urine of rats prior fed with ginger for a month and treated with B(a)P, a significant reduction (p<0.001) in the mutagenicity was observed as indicated by decreased No. of revertants and with both the tester strains namely TA98 and TA100. This observation was consistent with all the levels of ginger feeding as well as in the presence and absence of S9 mixture. The percentage of mutagenicity inhibition with tester strain TA98 without S9 was 25.2, 43.4 and 67.5 and with S9 was 23.8, 39.3 and 43.7 in the rats given ginger at the levels of 0.5, 1 and 5%, respectively (Table 5). In the absence of S9 the percentage inhibition with tester strain TA100 was found to be 17.3, 35.8 and 46.3 and in the presence of S9 was 27.6, 40.5 and 46.7 in the rats fed with ginger through diet at the levels of 0.5, 1 and 5%, respectively (Table 5).
| Table 2: | Initial and final body weights of rats fed diet with ginger for one month |
 | |
| Values are Mean±SD of 6 rats per group | |
| Table 3: | Inhibition of urinary mutagen levels in ginger fed rats |
 | |
| Values are mean±SE of 6 rats per group. Vertical comparisons: Values bearing different superscripts are significant at p<0.001 by ANOVA. Each sample was tested in duplicate plate assay | |
| Table 4: | Inhibition of urinary mutagen levels in ginger fed rats |
 | |
| Values are mean±SE of 6 rats per group Vertical comparisons: Values bearing different superscripts are significant at p<0.001 by ANOVA. Each sample was tested in duplicate plate assay | |
| Table 5: | Percent inhibition of urinary mutagens in ginger fed rats treated withbenzo(a)pyrene in TA 98 and TA100 |
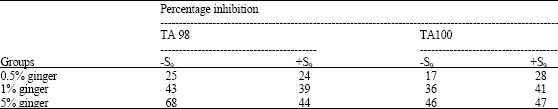 | |
| Table 6: | Antimutagenic potential of unboiled and boiled ginger paste-TA 98 (-S9) No. of histidine revertant colonies |
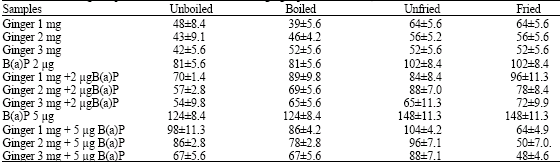 | |
| Spontaneous frequency of revertants (-S9) = 17.8±2.2 Values are mean±SD. Each sample was tested in duplicate plate assay. Between group differences were seen by means of Kruskal Wallis test | |
The dose response curves indicated a decrease in mutagenic load with increasing doses of ginger with the two tester strains TA98 and TA100.
| Table 7: | Antimutagenic potential of unboiled and boiled ginger powder-TA 98 (-S9) No. of histidine revertant colonies |
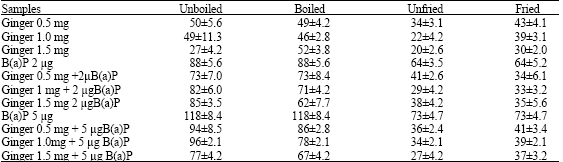 | |
| Spontaneous frequency of revertants (-S9) = 12.2±3.6. Values are mean±SD. Each sample was tested in duplicate plate assay. Between group differences were seen by means of Kruskal Wallis test | |
| Table 8: | Antimutagenic potential of unboiled and boiled ginger paste-TA 100 (-S9) No. of histidine revertant colonies |
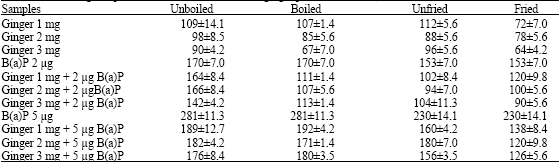 | |
| Spontaneous frequency of revertants (-S9) = 54±4.3. Values are mean±S.D. Each sample was tested in duplicate plate assay. Between group differences were seen by means of Kruskal Wallis test | |
| Table 9: | Antimutagenic potential of unboiled and boiled ginger powder-TA 100 (-S9) No. of histidine revertant colonies |
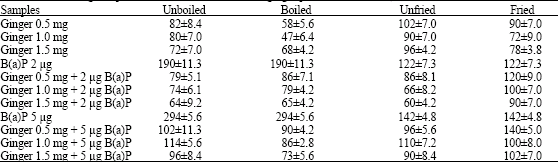 | |
| Spontaneous frequency of revertants (-S9) = 37.8±5.2. Values are mean±SD. Each sample was tested in duplicate plate assay. Between group differences were seen by means of Kruskal Wallis test | |
Effect of Unprocessed/Processed Ginger Paste and Powder on the Antimutagenic Potential Induced by B(a)P
Raw/powder forms of ginger irrespective of treatment (unboiled/boiled/fried/unfried) did not alter the antimutagenic potential of ginger when tested with 2 μg (2 μL/plate) and 5 μg (5 μL/plate) concentrations of B(a)P at the concentrations of 1, 2 and 3 mg of ginger paste and 0.5,1.0 and 1.5 mg ginger powder with both tester strains TA 98 (Table 6 and 7) and TA100 (Table 8 and 9). The antimutagenic potential of boiled/fried ginger was comparable to raw ginger.
The results clearly demonstrated that ginger has antimutagenic potential under in vivo and in vitro conditions. Cooking procedures involving boiling, frying is unlikely to alter its beneficial property.
DISCUSSION
Non-nutrient component of diet are reported to possess health-promoting properties. The discovery and exploration of chemical compounds with antimutagenic and antigenotoxic potency is of importance in view of the increased incidence of mutation related diseases like cancer. Phytochemicals present in plants are gaining wide recognition as possible antigenotoxicants (Krishnaswamy and Polasa, 2001).
Nutrients, which show modulatory effects in experimental cancers, include macronutrients such as fat, carbohydrates, proteins, fiber and micronutrients such as vitamins-folic acid, riboflavin, beta-carotene, retinol, α-tocopherol, vitamin B12 and minerals such as selenium, zinc, magnesium and calcium. Presently, however, the focus has shifted to a number of non-nutritional components in our diet that possess anticarcinogenic and antimutagenic properties (Wargovich, 1999).
In traditional medicine, spices and herbs have been extensively used to treat various ailments. Commonly consumed spices/vegetables like turmeric, mustard, garlic, onions were found to be antimutagenic. In principle, the antimutagenic substances can either inhibit the metabolic conversion or can deactivate directly the xenobiotics or enhance the cellular levels of carcinogen deactivating enzymes (Krishnaswamy and Polasa, 2001).
In this study, benzo(a)pyrene a ubiquitously present genotoxicant was used to treat experimental rats that were fed with ginger for a period of one month. The carcinogen that is present in high concentrations in environmental tobacco smoke is known to produce genotoxic effects (Van Zeeland, 1999).
The polycyclic aromatic hydrocarbons are metabolised to their epoxide derivatives, which can bind to DNA (Jeffrey and Straub, 1997). Alterations in DNA in the form of point mutations or macrolesions like adduct formations are key events in the process of carcinogenesis (Ames et al., 1995). Natural plant products such as phenols, indoles and flavonoids have been shown to have strong antimutagenic and anticarcinogenic effects against direct and indirect acting mutagens (Surh, 1999). Many tests employing prokaryotes to eukaryotes can be used to study antimutagenesis (Deflora and Sobels, 1992). In vitro test using two Salmonella strains TA 98 and TA 100 is one of the most commonly used and highly validated technique. In this technique, S9 fraction is employed to obtain preliminary information of an agent in order to stimulate the in vivo metabolism, as screening in animals for every chemopreventive substance will be cumbersome. This test system might aid in detecting antimutagens/ antigenotoxicants of relevance when used as part of a structural testing sequence. It has been proposed that mutagenesis testing of body fluids of treated animals could be an indirect in vivo screening procedure for potential carcinogens (Smith et al., 1996).
Increased urinary mutagenicity is noted following exposure to occupational, dietary or other environmental mutagens, while a decrease due to administration of putative antigenotoxic agent indicates a protective effect. In this study prior feeding of ginger for a period of one month could reduce the formation and excretion of urinary mutagens in benzo(a)pyrene treated rats. The mutagenic activity of the urine is indicated by the frequency of reversion in the bacterial strain Salmonella typhimurium TA 98 and TA100. Polycyclic aromatic hydrocarbon (PAHs) of which B(a)P is most commonly studied is formed by the incomplete combustion of organic matter. They are widely distributed in environment and human exposure is unavoidable. A number of them, such as B(a)P are carcinogenic and mutagenic and are believed to contribute substantially to the over all burden of cancer in humans. The PAHs’are converted to diol epoxides, which form adducts with DNA (Jeffrey and Straub, 1997). It is possible that ginger may exert its antimutagenic effect by counteracting the oxidative metabolism of carcinogen through induction of Phase II drug metabolism enzymes in host or by directly deactivating the carcinogen and its metabolites (Polasa and Nirmala, 2003). Many plant products are known to stimulate the cellular detoxifying enzymes (Lampe, 2003).
Cooking processes can generate PAHs (Phillips, 1999). Since dietary components undergo changes as a result of heat treatment during cooking, it was necessary to know if ginger could exert antimutagenic potential after heat treatment. The in vitro effect with different levels of ginger against two concentrations of benzo(a)pyrene suggested that the antimutagenicity of ginger was unaltered following heat treatment. This would be attributed to the antioxidant potential of ginger. In a study on garlic it was reported that the antimutagenic property was not destroyed on heat treatment during cooking processes (Polasa et al., 2006). Antioxidants in diet play a major role in counteracting the mutagenic substances that may be present in the diet itself or in environment (Demarini, 1996). Ginger is known to possess antioxidant and anti-inflammatory properties (Srivastava, 1986; Kikusaki and Nakatani, 1993). Phenolic compounds derived from Zingiberacea (ginger family) plants have been shown to induce apoptosis in human cancer cell lines and this has been attributed to their anti-inflammatory activity (Lee and Surh, 1998).
The correlation between bacterial mutagenicity and DNA adduct formation due to heterocyclic amines has been reported by Malfatti et al. (1995). The decreased mutagenic response in ginger treated rats as observed in this study suggests that reaction of carcinogen with DNA could be modified in the presence of ginger and or its active ingredients. Turmeric, which belongs to the family Zingiberaceae like ginger, has been shown to be antimutagenic (Polasa et al., 1991) and is now being considered as potential chemopreventer. Phytochemicals present in fruits, vegetables and spices possess several other beneficial properties. Identifying antigenotoxic potential of these foods will enable in modifying diet so as to protect against toxic effect of xenobiotics present in the environment (Surh, 1999).
Effectiveness and safety are the crucial requisites of candidate antigenotoxic agents for human use. Understanding of the mechanisms involved is essential for rational recommendation of their intakes through diet. Naturally occurring dietary components are unknown to produce toxic effects even when consumed in large proportions particularly in their natural form. Many of them are found in a variety of foods consumed daily. The beneficial effects of chemopreventers in preventing not only cancer but also chronic diseases, their easy availability, low toxicity and low cost have promoted an increased interest in these chemicals. This interest has been developed not only for their presence in the diet, but also in isolating them and fortifying other foods with them.
Diet survey conducted by Thimmayamma et al. (1983) reported that an intake of 2.6 g/consumption unit/day is being consumed in Indian population. Our suggestion of 3-5 g/day is within the normal intake that is existing in the population and hence daily intake of ginger may be recommended to ward off tissue/cell damage associated with degenerative diseases. The results of this study indicate that ginger per se exerts antimutagenic action against carcinogens in vivo and in vitro. These observations are important in view of the widespread exposure to environmental genotoxicants.
ACKNOWLEDGMENTS
The authors wish to thank Spices Board, Cochin, India for supporting in part by grants.
REFERENCES
- Ames, B.N., L.S. Gold and W.C. Willet, 1995. The causes and prevention of cancer. Proc. Natl. Acad. Sci. USA., 92: 5258-5265.
PubMedDirect Link - Deflora, S. and F. Sobels, 1992. Assessment and antimutagenicity and anticarcinogenicity. End points and systems. Mut. Res., 267: 153-295.
Direct Link - Doll, R. and R. Peto, 1981. The causes of cancer: quantitative estimates of avoidable risks of cancer in the United States today. J. Natl. Cancer Inst., 66: 1191-1308.
PubMed - Ikkeny, Y., P. Morales, A. Martinez, M.L. Marin, A.I. Haza and M. Cambero, 1999. Antimutagenic effect of fruit and vegetable ethanolic extracts against N-nitrosamines evaluated by the Ames test. J. Agric. Fd. Chem., 47: 3257-3264.
Direct Link - Johanna, W.L., 2003. Spicing up a vegetarian diet: Chemopreventive effects of phytochemicals. Am. J. Clin. Nutr., 78: 579S-583S.
Direct Link - Lee, E. and Y.J. Surh, 1998. Induction of apoptosis in HL-60 cells by pungent vanilloids, [6]-gingerol and [6]-paradol. Cancer Lett., 134: 163-168.
CrossRefPubMedDirect Link - Maron, D.M. and B.N. Ames, 1983. Revised methods for the Salmonella mutagenicity test. Mutat. Res./Environ. Mutagen. Relat. Subj., 113: 173-215.
CrossRefDirect Link - Nirmala, K., T.P. Krishna and K. Polasa, 2007. Protective effect of ginger against benzo(a)pyrene induced DNA damage. Int. J. Cancer Res., 3: 13-24.
CrossRefDirect Link - Park, E.J. and J.M. Pezzuto, 2002. Botanicals in cancer chemoprevention. Cancer Metastasis Rev., 21: 231-255.
CrossRefDirect Link - Phillips, D.H., 1999. Polycyclic aromatic hydrocarbons in the diet. Mutat. Res./Genet. Toxicol. Environ. Mutagen., 443: 139-147.
CrossRefDirect Link - Polasa, K., B. Sesikeran, T.P. Krishna and K. Krishnaswamy, 1991. Turmeric (Curcuma longa) induced reduction in urinary mutagens. Food Chem. Toxicol., 29: 699-706.
Direct Link - Smith, T.J., S.C. McKarns, R.A. Davis, S.D. Livingston and B.R. Bombick et al., 1996. Human urine mutagenicity study comparing cigarettes which burn or primarily heat tobacco. Mut. Res., 361: 1-9.
Direct Link - Surh, Y.J., 1999. Molecular mechanisms of chemopreventive effects of selected dietary and medicinal phenolic substances. Mutat. Res., 428: 305-327.
Direct Link - Van Zeeland, A.A., A.J.L. Degroot, J. Hall and J. Donato, 1999. 8-hydroxyguanosine in DNA from leukocyte of healthy adults; relationship with cigarette smoking, environmental tobacco smoke, alcohol and coffee consumption. Mut. Res., 439: 249-257.
Direct Link - Wargovich, M.J., 1999. Nutrition and cancer: The herbal revolution. Curr. Opin. Clin. Nutr. Metabolic Care, 2: 421-424.
Direct Link - Wattenberg, L.W., 1992. Inhibition of carcinogenesis by minor dietary constituents. Cancer Res., 52: 2085S-2091S.
Direct Link - Kikuzaki, H. and N. Nakatani, 1993. Antioxidant effects of some ginger constituents. J. Food Sci., 58: 1407-1410.
CrossRefDirect Link