
Totiya Ishchi
Department of Biology, Education Faculty, Jawzjan University, Sheberghan, Afghanistan
G. Sibi
Department of Biotechnology, Indian Academy Degree College-Autonomous, Bengaluru, India
LiveDNA: 91.5767
International Journal of Biological Chemistry
Year: 2020 | Volume: 14 | Issue: 1 | Page No.: 1-7







ABSTRACT
Background and Objective: Azo dyes are most widely used in various industries due to their ease of synthesis and chemical stability. But many azo dyes and their breakdown derivatives have been suggested to be toxic to life. Biological treatments of dyes with microorganisms are relatively cost effective and eco-friendly due to lower sludge production. In this study, the microalgae Chlorella vulgaris was examined for its degrading ability of azo dyes viz., Reactive Black 5, Direct Blue 71 and Disperse Red 1. Materials and Methods: The experiments included dye decolorization experiments and factors affecting decolorization such as initial dye concentration, pH and temperature. Further, azoreductase enzyme specific activity and its kinetics were determined. Results: The results revealed that azo degradation based on the initial dye concentration varied among the dyes tested and the same was reflected for pH. In specific, 200 mg L–1 was found optimum for Reactive Black 71 and Direct Blue 71 whereas 300 mg L–1 was for Disperse Red 1. The optimum pH for decolorization of RB5, DB71 and DR1 was 5, 8 and 8, respectively. The optimum temperature for degradation of all the dyes was found as 40°C. Maximum azoreductase activity of 0.127 U mg–1 protein was observed for DR1 and the enzyme activity was not pH dependent but substrate specific. Conclusion: The results revealed the azo dye degrading potential of Chlorella vulgaris and the decolorizing conditions were optimized for the microalgae.
PDF Abstract XML References Citation
Received: September 19, 2019;
Accepted: October 30, 2019;
Published: January 02, 2020
How to cite this article
Totiya Ishchi and G. Sibi, 2020. Azo Dye Degradation by Chlorella vulgaris: Optimization and Kinetics. International Journal of Biological Chemistry, 14: 1-7.
URL: https://scialert.net/abstract/?doi=ijbc.2020.1.7
URL: https://scialert.net/abstract/?doi=ijbc.2020.1.7
INTRODUCTION
Synthetic dyes could cause serious pollution to the environment and ecology, because the complex aromatic molecular structures of these dyes are difficult to degrade1. Dyes are emitted into wastewaters from various industrial branches, mainly from the dye manufacturing and textile finishing and also from food colouring, cosmetics, paper and carpet industries2,3. Large water bodies contaminated with low concentrations of dyes leads to reduced light penetration and photosynthesis.
Dyes having chromophore group with a pair of conjugated aryl moieties linked by an azo (-N = N-) bond are called azo dyes. Azo dyes are the most widely used and large amounts are produced annually around the world and used extensively in textile, printing, cosmetic, plastic, food, pharmaceutical and many other industries due to their ease of synthesis and chemical stability4. Effluents containing azo dyes must be treated before discharge into environment not only because they may affect the transparency and aesthetics of water bodies but also because many azo dyes and their breakdown derivatives have been suggested to be toxic/mutagenic to life5,6. Generally, dye containing waste water is decolorized using physicochemical process such as adsorption, oxidation, photocatalysis. However, high cost and formation of secondary pollutants associated with these methods restrict their application7-9.
Biological treatments of dyes with microorganisms are relatively cost effective and eco-friendly due to lower sludge production. Azo dyes are initially reduced/decolorized by various microbes under anaerobic or anoxic conditions to corresponding aromatic amines, some of which are then further mineralized aerobically10,11. Among the dye degrading microorganisms, microalgae have been involved in degrading textile dyes and the reported elsewhere12-19.
In this study, the microalgae Chlorella vulgaris was examined for its degrading ability of azo dyes viz., Reactive Black 5, Direct Blue 71 and Disperse Red 1.
MATERIALS AND METHODS
Azo dyes used: The synthetic azo dyes Reactive Black 5, Direct Blue 71 and Disperse Red 1 (Fig. 1a-c) were purchased from Sigma-Aldrich, India.
Determination of absorption maxima (λmax) and plotting of standard curve: The absorption of light by each dye and mixture of dyes within the visible range (400-700 nm) was determined to calculate the adsorption maxima (λmax) using UV-Vis spectrophotometer (Systronics, India).
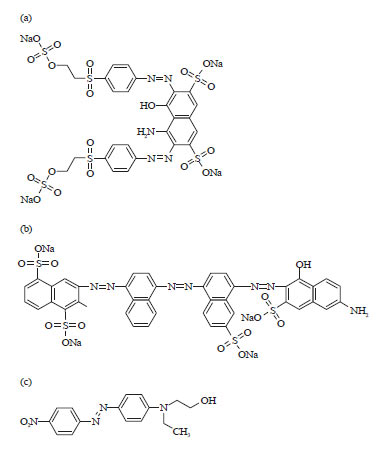 | |
| Fig. 1(a-c): | (a) Chemical structure of Reactive Black 5, (b) Direct Blue 71 and (c) Disperse Red 1 |
Accordingly, the wavelength for Reactive Black 5, Direct Blue 71 and Disperse Red 1 was determined as 597, 595 and 505 nm, respectively. Known concentrations (0, 0.1, 1, 10 and 100 mg L–1) of each dye from decolorizing medium was prepared to plot the standard curve. Using scatter diagram, trend line was added and then the R2 value was calculated. The percentage (%) of dye decolorization was calculated using the same equation.
Decolorization study and spectroscopic analysis: The experiments were conducted in sterilized 250 mL Erlenmeyer flask containing 120 mL of the sterile mineral salt media (MSM) with 30 mL of algal culture and the azo dyes at a concentration of 100 mg L–1. All of the treated flasks were incubated for 12 days at 30°C with constant fluorescent illumination. Control experiments were conducted in light and dark conditions in the absence of microalgae to evaluate the effect of photodegradation. The degradation ratio was determined after 14 days of incubation by measuring the absorbance of the cell free supernatant of the sample at the corresponding λmax of each dye solution as mentioned earlier by a double beam UV-Visible spectrophotometer. The percentage of decolorization was calculated by using the equation according to Telke et al.20:
![]()
Optimization of environmental factors for efficient decolorization
Effect of initial dye concentration on decolourization: Optimization of decolourization based on initial dye concentration was carried out at by adding different dye concentrations ranging from 100-500 mg L–1 to mineral salt media (MSM) and incubated at ambient temperature in static conditions.
Effect of pH and temperature on decolourization: Effect of pH on decolourization was studied at different initial pH values and incubation temperatures by following the same procedure as described earlier. To study the effects of pH, flasks were maintained at ambient temperature and the pH of the medium was adjusted to values between 4 and 10 by adding 1 moL–1 HCl or 1 moL–1 NaOH. The optimal concentration of each dye was used for the experiments. To examine the effect of incubation temperature, the flasks were incubated in temperature which varied from 20-50°C. Optimal initial dye concentration and pH of each dye were used to ascertain the experimental conditions.
Crude enzyme extraction: The algal cells were grown in MSM media under optimized conditions for each dye tested were investigated for the production of azo-reductase enzyme. Cells from experimental and control flasks were harvested by centrifugation at 12,000 rpm for 10 min. Phosphate buffer (100 mM, pH 7.4) was used to wash the pellet and the cells were suspended in the same buffer. This was followed by sonicating the cells at 200 amp, giving 99 stroke, each of 3 sec with 7 sec interval at 4°C to lyse the cells. In the final step, the lysed cell suspension was centrifuged at 5000 rpm for 30 min at 4°C and the supernatant was then used for further analysis21.
Azo-reductase specific activity: Azo-reductase activity was determined by the reduction of azo dyes (Reactive Black 5, Direct blue 71, Disperse Red 1) as the main substrate and oxidation of NADH. The assay mixture consisted of 600μL phosphate buffer (50mM, pH 7.5), 100μL dye (50mM), 100μL the crude enzyme and 200 μL NADH (200μM). One unit of azo-reductase activity was defined as 1μg of dye reduced/mg protein/min22. All enzyme assays were run in triplicate.
Azo reductase kinetics: Different concentrations of dyes (ranged from 20-3800 mM) were prepared to determine the azo reductase kinetics. All other constituents of reaction mixture were constant. Kinetic values such as Michaelis-Menten constant (Kmax) and maximal velocity (Vmax) were calculated from Lineweaver-Burk double reciprocal plots23.
RESULTS AND DISCUSSION
Decolorization experiments involving 100 mg L–1 concentration of dye in growth media was carried out for a period of 12 days. In all the experiments, more than 90% of the dye was decolorized by C. vulgaris (Fig. 2) within 10 days. Based on the time taken for higher decolorization percentage, 10 days cultivation period was chosen for optimizing the dye degradation.
Results of initial dye decolorization experiments revealed that 80% of the dye was decolorized at 200 mg L–1 concentration of Reactive Black 5 (RB5), 78% decolorization of Direct Blue 71 (DB71) at 300 mg L–1 concentration and 84% decolorization of Disperse Red 1 (DR1) at 200 mg L–1 concentration (Fig. 3). Dye removal by the algal species is dependent on concentration of dye and algal biomass24. However, Yang et al.25 found that there was a negative correlation between dye removal and initial dye concentration when using Shewanella oneidensis to decolorize acid yellow 199. Dye concentrations were selected based on highest decolorization percentage and used to optimize the pH and temperature conditions for effective decolorization. Concentration of 200 mg L–1 for RB5 and DR1and 300 mg L–1 for DB71 was used to optimize the decolorization based on pH values of the growth medium. The pH in the range between 4 and 10 were selected and the highest decolorization of RB 5 was found at pH 5. Further increase in pH beyond 6 and decrease in pH below 5 resulted in decreased percentage of dye decolorization of RB 5. Decolorization percentage of 84 and 79% were obtained for DB71 and DR1 at pH 8. Increase in pH beyond 8 and decrease in pH below 7 resulted in decreased percentage of dye decolorization for both the dyes (Fig. 4).
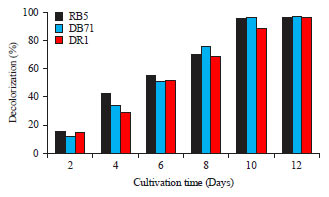 | |
| Fig. 2: | Decolorization (%) by C. vulgaris at 100 mg L–1 dye concentration |
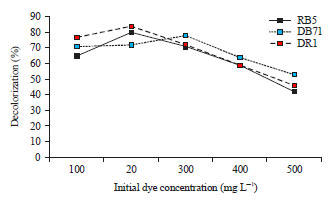 | |
| Fig. 3: | Effect of initial dye concentration on decolorization (%) by C. vulgaris |
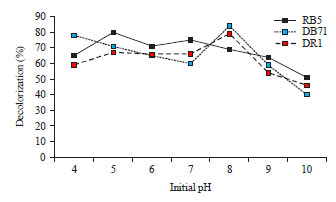 | |
| Fig. 4: | Effect of initial pH on dye decolorization (%) by C. vulgaris |
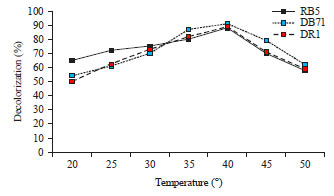 | |
| Fig. 5: | Effect of temperature on dye decolorization (%) by C. vulgaris |
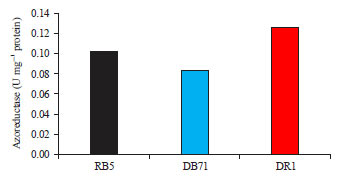 | |
| Fig. 6: | Azoreductase specific activity (U mg–1 protein) of C. vulgaris on different substrates at 40°C, 50 mM substrate concentration and 200 mM NADH |
Cells of microbes get ruptured under alkaline conditions which expose additional functional groups and solubilize certain cell constituents26. This enhances the dye removal by the organisms from the solutions. The optimum pH was found to be at 8 for maximal removal of dyes (DB71 and DR1). The pH has a major effect on the efficiency of dye decolorization and the optimal pH for color removal is often between 6.0 and 10.0 for most of the dyes27. However, the optimum pH for highest decolorization of RB1 was found as pH 5. Similarly, Ozer et al.28 found that acidic pH as optimum for the degradation of acid blue dyes by Spirogyra rhizopus.
To examine the effect of incubation temperature, the flasks were incubated in temperature which varied from 20-50°C (Fig. 5). The initial dye concentration, pH for RB 1 and DB71 was 200 mg L–1, pH 5 and 300 mg L–1, pH 8, respectively whereas, it was 200 mg L–1 and pH 8 for DR1. Among different temperature tested, 40°C was found to be an optimum on dye decolorization of RB 1, DB71 and DR1. It is noted that the percentage decolorization of dye increased with an increase in temperature from 20-40°C. The percentage decolorization of dye decreased with further increase in temperature up to 50°C.
Similar results were reported by Cetin et al.10 where the decolourization activity was significantly suppressed above 45°C. The reason for this may be due to the loss of cell viability or deactivation of the enzymes responsible for decolourization at higher temperatures. Ozer et al.28 reported that mesophilic temperature was optimum for the degradation of acid blue by Spirogyra rhizopus.
Azoreductase specific activity against specific substrate was determined at 50 mM substrate concentration, 200 mM NADH and optimized pH and temperature of each dye tested. Highest azoreductase activity of 0.127 U mg–1 proteins was observed for DR1 followed by 0.103 U mg–1 protein for RY22. Lowest activity of 0.084 U mg–1 proteins was determined for DB71 (Fig. 6).
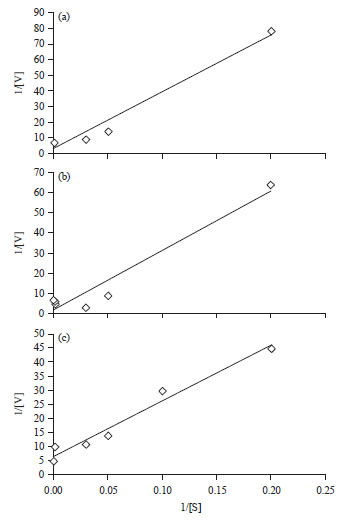 | |
| Fig. 7(a-c): | A lineweaver-Burk plot for determination the kinetic parameters of azoreductase, (a) Reactive Black 5, (b) Direct Blue 71 and (c) Disperse Red 1 |
El-Sheekh et al.18 had measured the azoreductase activity of Microcystis aeruginosa and Pseudanabaena sp. while degrading Disp.orange 2RL, Reactive yellow 3RN, Reactive Black NN and Tracid Red BS. Treatment of M. aeruginosa with Disp.orange 2RL induced the azo reductase enzyme by 68.04% and Pseudanabaena sp. with Reactive yellow 3RN by 55.64% after 7 days of incubation.
Azo reductases have been shown to be very specific enzymes thus cleaving only the azo bonds of azo dyes and the azoreductase reaction should take place at optimum condition. The most important condition affecting on the activity of the enzymes is pH and temperature. Hence, it is important to determine the optimal conditions for the operation of enzymes. In this study, maximum azoreductase activity of 0.127 U mg–1 proteins was observed for DR1 which has used pH 8. It was also noted that the enzyme activity was not pH dependent but substrate specific. In other words, the second highest azoreductase activity was observed for RB5 at pH 5. The lowest enzyme activity of 0.084 U mg–1 proteins was observed for DB71 though it was studied at pH 8. Similar experiments were carried out earlier to optimize the azoreducatase production by other microorganisms29,30. Other conditions which affect dye decolorization by microalgae also reported. Azo dye reduction under saline conditions by Shewanella strains were investigated by Liu et al.31. Under anaerobic conditions, both strains could effectively decolorize real textile effluents. Shewanella algae reached over 87.8% decolorization in 48 h, whereas, Shewanella marisflavi removed nearly 80% color of the effluents in 72 h. Guolan et al.12 reported that immobilized algae is better than free algae in degradation of Better brown.
Using a different concentration of the substrate (RB5, DB71, DR1) is a main factor to determine the enzyme kinetics. The kinetics parameters like Vmax (maximum velocity) and Km (Michaelis constant) were estimated and the velocity of azoreductase in C. vulgaris was 1.67, 0.72 and 1.49 μM min–1 mg–1 for RB5, DB71 and DR1, respectively (Fig. 7a-c). However, the value Km were 2069, 247 and 2468 μM.
The experiments were carried out for a period of 10 days and the optimal conditions of initial dye concentration, pH and temperature were determined. The initial dye concentration varied among the dyes tested and the same was reflected for pH. In specific, 200 mg L–1 was found optimum for Reactive Black 71 and Direct Blue 71 whereas, 300 mg L–1 was for Disperse Red 1. Similarly, the optimum pH for decolorization of RB5, DB71 and DR1 was 5, 8 and 8, respectively. In a study by Baldev et al.16, the optimum operating conditions for degradation of Rhodamine by Coelastrella were found to be 100 mg L−1 initial dye concentration, 30°C with 10% of inoculum at a pH of 8 within 20 days. In another study, 30 days was used to screen the cyanobacterial degradation of textile dyes and Gloeocapsa pleurocapsoides was able to remove 90% of FF Sky blue from the medium13. In this work, C. vulgaris have exhibited sufficient dye degradation potential of dyes commonly used in textile industries. Depending on the final results in this study, it can be concluded that C. vulgaris can be used for removing these dyes. Further, knowledge from present work may be employed on large scale with in situ applications.
CONCLUSION
This study revealed the azo dye degrading potential of Chlorella vulgaris. The optimum pH was found to be at 8 for maximal removal of Direct Blue 71 and Disperse Red 1. It is noted that the percentage decolorization of dye increased with an increase in temperature from 20-40°C. Highest azoreductase activity of 0.127 U mg–1 proteins was observed for Disperse Red 1 and the enzyme activity was not pH dependent but substrate specific. The results revealed that azo dye degrading potential of C. vulgaris was independent on pH, temperature and substrate concentration.
SIGNIFICANCE STATEMENT
This study discovers the azo dye degrading potential of C. vulgaris that can be beneficial for treating textile industry effluents. Implementing the optimized parameters will uncover the critical areas of associated with in situ biodegradation of textile dyes. A new theory on determination of enzyme kinetics is necessary for the effective azo dye degradation by microalgae may be arrived at.
REFERENCES
- Tomei, M.C., D.M. Angelucci and A.J. Daugulis, 2016. Sequential anaerobic-aerobic decolourization of a real textile wastewater in a two-phase partitioning bioreactor. Sci. Total Environ., 573: 585-593.
CrossRefDirect Link - Mall, I.D., V.C. Srivastava, N.K. Agarwal and I.M. Mishra, 2005. Removal of congo red from aqueous solution by bagasse fly ash and activated carbon: kinetic study and equilibrium isotherm analyses. Chemosphere, 61: 492-501.
CrossRefDirect Link - Namasivayam, C. and D. Kavitha, 2002. Removal of Congo Red from water by adsorption onto activated carbon prepared from coir pith, an agricultural solid waste. Dye Pigments, 54: 47-58.
CrossRefDirect Link - Stolz, A., 2001. Basic and applied aspects in the microbial degradation of azo dyes. Applied Microbiol. Biotechnol., 56: 69-80.
CrossRefDirect Link - Brown, M.A. and S.C. De Vito, 1993. Predicting azo dye toxicity. Criti. Rev. Environ. Sci. Technol., 23: 249-324.
CrossRefDirect Link - Xu, H., T.M. Heinze, S. Chen, C.E. Cerniglia and H. Chen, 2007. Anaerobic metabolism of 1-amino-2-naphthol-based azo dyes (Sudan dyes) by human intestinal microflora. Applied Environ. Microbiol., 73: 7759-7762.
CrossRefDirect Link - Robinson, T., G. McMullan, R. Marchant and P. Nigam, 2001. Remediation of dyes in textile effluent: A critical review on current treatment technologies with a proposed alternative. Bioresour. Technol., 77: 247-255.
CrossRefDirect Link - Pearce, C.I., J.R. Lloyd and J.T. Guthrie, 2003. The removal of colour from textile wastewater using whole bacterial cells: A review. Dyes Pigments, 58: 179-196.
CrossRef - Chen, K.C., J.Y. Wu, D.J. Liou and S.C.J. Hwang, 2003. Decolorization of the textile dyes by newly isolated bacterial strains. J. Biotechnol., 101: 57-68.
CrossRefDirect Link - Cetin, D. and G. Donmez, 2006. Decolorization of reactive dyes by mixed cultures isolated from textile effluent under anaerobic conditions. Enzyme Microb. Technol., 38: 926-930.
CrossRefDirect Link - Dos Santos, A.B., F.J. Cervantes and J.B. van Lier, 2007. Review paper on current technologies for decolourisation of textile wastewaters: Perspectives for anaerobic biotechnology. Bioresour. Technol., 98: 2369-2385.
CrossRefDirect Link - Guolan, H., S. Hongwen and C.L. Li, 2000. Study on the physiology and degradation of dye with immobilized algae. Artif. Cells Blood Substitutes Biotechnol., 28: 347-363.
CrossRefDirect Link - Parikh, A. and D. Madamwar, 2005. Textile dye decolorization using cyanobacteria. Biotechnol. Lett., 27: 323-326.
CrossRefDirect Link - Daneshvar, N., M. Ayazloo, A.R. Khataee and M. Pourhassan, 2007. Biological decolorization of dye solution containing Malachite Green by microalgae Cosmarium sp. Bioresour. Technol., 98: 1176-1182.
CrossRefDirect Link - Vijayakumar, S. and C. Manoharan, 2012. Treatment of dye industry effluent using free and immobilized cyanobacteria. J. Bioremediat. Biodegrad., Vol. 3.
CrossRefDirect Link - Baldev, E., D. MubarakAli, A. Ilavarasi, D. Pandiaraj, K.A.S.S. Ishack and N. Thajuddin, 2013. Degradation of synthetic dye, Rhodamine B to environmentally non-toxic products using microalgae. Colloids Surf. B: Biointerfaces, 105: 207-214.
CrossRefPubMedDirect Link - El-Kassas, H.Y. and L.A. Mohamed, 2014. Bioremediation of the textile waste effluent by Chlorella vulgaris. Egypt. J. Aquat. Res., 40: 301-308.
CrossRefDirect Link - El-Sheekh, M., W.G. Abou-El-Souod and H.A. Al Asrag, 2017. Biodegradation of some dyes by the cyanobacteria species Pseudoanabaena sp. and Microcystis aeruginosa Kützing. Egypt. J. Exp. Biol. (Bot.), 13: 233-243.
Direct Link - Dellamatrice, P.M., M.E. Silva-Stenico, L.A.B. de Moraes, M.F. Fiore and R.T.R. Monteiro, 2017. Degradation of textile dyes by cyanobacteria. Braz. J. Microbiol., 48: 25-31.
CrossRefDirect Link - Telke, A.A., S.M. Joshi, S.U. Jadhav, D.P. Tamboli and S.P. Govindwar, 2010. Decolorization and detoxification of Congo red and textile industry effluent by an isolated bacterium Pseudomonas sp. SU-EBT. Bidegradation, 21: 283-296.
CrossRefDirect Link - Zhao, M., P.F. Sun, L.N. Du, G. Wang, X.M. Jia and Y.H. Zhao, 2014. Biodegradation of methyl red by Bacillus sp. strain UN2: Decolorization capacity, metabolites characterization and enzyme analysis. Environ. Sci. Pollut. Res., 21: 6136-6145.
CrossRefDirect Link - Du, L.N., S. Wang, G. Li, B. Wang, X.M. Jia, Y.H. Zhao and Y.L. Chen, 2011. Biodegradation of malachite green by Pseudomonas sp. strain DY1 under aerobic condition: Characteristics, degradation products, enzyme analysis and phytotoxicity. Ecotoxicology, 20: 438-446.
CrossRefDirect Link - Wilkinson, G.N., 1961. Statistical estimations in enzyme kinetics. Biochem. J., 80: 324-332.
CrossRefPubMedDirect Link - Mohan, S.V., C.N. Roa, K.K. Brasad and J. Karthikeyan, 2002. Treatment of simulated reactive yellow 22 (Azo) dye effluents using Spirogyra species. Waste Manage., 22: 575-582.
CrossRefDirect Link - Yang, Y.Y., L.N. Du, G. Wang, X.M. Jia and Y.H. Zhao, 2011. The decolorisation capacity and mechanism of Shewanella oneidensis MR-1 for methyl orange and acid yellow 199 under microaerophilic conditions. Water Sci. Technol., 63: 956-963.
CrossRefDirect Link - Nagase, H., D. Inthorn, Y. Isaji, A. Oda, K. Hirata and K. Miyamoto, 1997. Selective cadmium removal from hard water using NaOH-treated cells of the cyanobacterium Tolypothrix tenuis. J. Ferment. Bioeng., 84: 151-154.
CrossRefDirect Link - Chen, K.C., W.T. Huang, J.Y. Wu and J.Y. Houng, 1999. Microbial decolorization of azo dyes by Proteus mirabilis. J. Ind. Microbiol. Biotechnol., 23: 686-690.
CrossRefPubMedDirect Link - Ozer, A., G. Akkaya and M. Turabik, 2006. Biosorption of acid blue 290 (AB 290) and acid blue 324 (AB 324) dyes on Spirogyra rhizopus. J. Hazard. Mater., 135: 355-364.
CrossRefPubMedDirect Link - Supaka, N., K. Juntongjin, S. Damronglerd, M.L. Delia and P. Strehaiano, 2004. Microbial decolorization of reactive azo dyes in a sequential anaerobic-aerobic system. Chem. Eng. J., 99: 169-176.
CrossRefDirect Link - Shah, M.P., K.A. Patel, S.S. Nair, A.M. Darji and S. Maharaul, 2013. Microbial degradation of azo dye by Pseudomonas spp. Mps-2 by an application of sequential microaerophilic and aerobic process. Am. J. Microbiol. Res., 1: 105-112.
CrossRefDirect Link - Liu, G., J. Zhou, X. Meng, S.Q. Fu, J. Wang, R. Jin and H. Lv, 2013. Decolorization of azo dyes by marine Shewanella strains under saline conditions. Applied Microbiol. Biotechnol., 97: 4187-4197.
CrossRefDirect Link