
M.N.M. Esmaail
Department of Genetics and Cytology, Molecular Cytogenetics Group, Genetic Engineering and Biotechnology Division, National Research Center, Dokki, Giza, Egypt
E.A.A. Abd-Elhady
Department of Genetics and Cytology, Molecular Cytogenetics Group, Genetic Engineering and Biotechnology Division, National Research Center, Dokki, Giza, Egypt
S.A. Edris
Department of Genetics, Faculty of Agriculture, Ain Shams University, Egypt
F.M. El Domyati
Department of Genetics, Faculty of Agriculture, Ain Shams University, Egypt
N.R. Abdelhamid
Department of Genetics and Cytology, Molecular Cytogenetics Group, Genetic Engineering and Biotechnology Division, National Research Center, Dokki, Giza, Egypt
A.H.M. Hassan
Department of Genetics and Cytology, Molecular Cytogenetics Group, Genetic Engineering and Biotechnology Division, National Research Center, Dokki, Giza, Egypt
A.A. El Seoudy
Department of Genetics, Faculty of Agriculture, Ain Shams University, Egypt
International Journal of Agricultural Research
Year: 2012 | Volume: 7 | Issue: 3 | Page No.: 107-120







ABSTRACT
The present study was carried out to amplify glutenin genes to follow up any variations among different species of wheat genomes. Four designed specific primers of HMW-GS (HMW SX and HMW SY) and LMW-GS (LMW 7,8 and LMW S1) were used. In respect to the HMW-GS, the Subunit y gave higher polymorphism than subunit x, which failed to amplify gene patterns with diploid source of genome A and tetraploid species. While the polymorphism of subunit x appeared in variable size. In contrast, primers LMW 7, 8 and LMW S1 showed variations among the wheat species. The qualitative analysis of gene expression of HMW genes was studied by the SDS-PAGE, which was supported for the molecular studies and more informative approach to detect variations between and within species. Generally, the results of SDS-PAGE showed that, the absence or presence of GluA1 allele seemed to be responsible for clustering the cultivars and the species.
PDF Abstract XML References Citation
Received: January 17, 2012;
Accepted: March 01, 2012;
Published: June 02, 2012
How to cite this article
M.N.M. Esmaail, E.A.A. Abd-Elhady, S.A. Edris, F.M. El Domyati, N.R. Abdelhamid, A.H.M. Hassan and A.A. El Seoudy, 2012. Discriminative Features of Some High and Low Molecular Weight Glutenin Genes in Different Wheat Species. International Journal of Agricultural Research, 7: 107-120.
DOI: 10.3923/ijar.2012.107.120
URL: https://scialert.net/abstract/?doi=ijar.2012.107.120
DOI: 10.3923/ijar.2012.107.120
URL: https://scialert.net/abstract/?doi=ijar.2012.107.120
INTRODUCTION
Wheat is one of the major cereal crops in the world. It is a monocotyledonous that belongs to the grass family Poaceae, which encompasses approximately 8,700 species from which about 650 belong to triticeae tribe (Judd et al., 2002). There are two important species of the genus Triticum; Triticum durum; 2n = 2x = 28, AABB (durum wheat) and Triticum aestivum (bread wheat) which are the most common species grown and distributed widely in Egypt. Triticum durum is an allotetraploid originated through intergeneric hybridization and polyploidization involving two diploid grass species; Triticum urartu and Triticum monococcum 2n = 2x = 14, AA genome and B diploid genome related to A. speltoides 2n = 2x = 14, SS genome. T. aestivum, 2n = 2x = 42, AABBDD which is allohexaploid includes A, B and D genomes resulted from the hybridization between Triticum turgidum (AABB) and Aegilops tauschii (DD).
The major seed storage proteins of wheat are called prolamins because of their high proline and glutamine content. Wheat prolamins are classified into two major groups; gliadins and glutenins. Gliadins are monomeric proteins that form intra-molecular disulphide bonds, whereas glutenins are polymeric proteins, which are held together by inter-molecular disulphide bonds thereby forming the glutenin polymer (Shewry and Tatham, 1997). Wheat gluten contains both high molecular Weight (HMW-GS) and low molecular weight (LMW-GS) glutenin subunits. The effect of HMW-GS on dough properties (strength and elasticity) may be additive or synergistic with significant interactions with LMW-GS subunits. The HMW-GS are encoded by polymorphic genes at the Glu-1 loci that are present on the long arm of group 1 chromosomes. The HMW glutenin subunits are encoded by Glu-1 loci on chromosomes 1A, 1B and 1D in bread wheat, each locus encoding one x-type and one y-type subunits (Payne, 1987). Due to gene silencing and allelic variation, the composition of HMW glutenin subunits usually differs among hexaploid wheat varieties (Wan et al., 2002).
Although, the role of HMW-GS in bread-making quality is better understood, LMW-GS also plays a significant role in the formation of large polymers. Some allelic forms of LMW-GS showed greater effects on these properties than HMW-GS (Gupta et al., 1989).
The genes coding for subunits 1Bx, 1Dx and 1Dy are always expressed, while genes coding for subunits 1Ax and 1By are expressed in some cultivars (Payne, 1987). 1Ay subunit is occasionally present in hexaploid wheat (Johansson et al., 1995) and more widely in A-genome diploids (Wan et al., 2002; Jiang et al., 2009). Many genes of HMW glutenin subunits have been isolated from bread wheat, where the related wild species and the coding region sequences are highly homologous (Wan et al., 2002; Yan et al., 2008; Liu et al., 2007). This study aimed to investigate high and low molecular weight of glutenin protein in different species of wheat.
MATERIALS AND METHODS
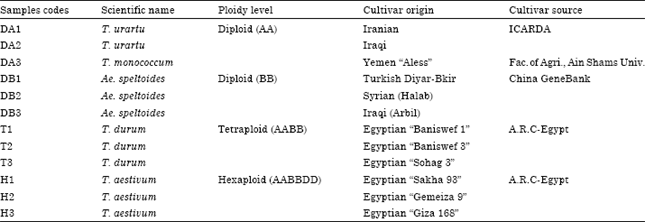
Twelve samples in total were used, classified into four groups, each of them was represented by three cultivars (Table 1); the first group refers to the diploid species that are the source for genome A (T. urartu (DA1, DA2) and T. monococcum (DA3). The second group represented the diploid species coming from B genome (DB1, DB2, DB3) which belong to Ae. speltoides. The third one represents the tetraploid cultivars T. durum (T1, T2, T3). The fourth group is the hexaploid cultivars T. aestivum (H1, H2, H3).
Seeds of tetraploid and hexaploid cultivars were supplied by the Agriculture Research Center (ARC), Egypt. While the diploid (AA) cultivars were supported from ICARDA and cultivars of the diploid source (BB) were supported by the GeneBank of china, each species represented by three cultivars. One seed was used to extract the DNA and protein content; half for DNA extraction and the other one for the protein extraction and three replicates of each cultivar were used to test the homogeneity of the samples as a pre-step.
| Table 1: | Wheat species, code of names, scientific cultivar names, their ploidy models, which genome they represent and sample sources |
 | |
Glutenin gene amplification
Extraction of genomic DNA (Template): The DNA was extracted from the seeds according to the protocol of Biospin plant genomic DNA extraction Kit (BioFlux). Agarose Gel electrophoresis was performed for genomic DNA with 0.8% agarose gel (Axygen) and stained by ethidium bromide stain (Sigma).
Allele specific PCR: Allele specific PCR or (AS-PCR) is a technique to amplify a gene product specifically from one allele (Saiki et al., 1985). Selectivity is achieved using specific primers, which match only one of the alleles of the gene.
Glutenin primers: This study aimed to detect some glutenin genes by designing some primers, some of which might detect HMW-GS and others detect LMW-GS. Primers names, forward and reverse sequences are presented in Table 2.
PCR amplification conditions: Amplification of glutenin genes were done according to (Yan et al., 2002), AS-PCR approach was performed to detect some glutenin genes in 30 μL, using 15 μL GC-buffer I, 8.8 μL ddH2O, 3 μL dNTPS, 1 μL for each primer, 1 μL DNA template from the extracted DNA and 0.2 μL LA Taq polymerase (all of them were produced from TaKaRa, Japan).
The following PCR program was used in a DNA Mastercycler, Eppendorf, Germany. Initial denaturation at 94°C for 4 min followed by 35 cycles of 94°C for 45 Sec. 57 to 58°C for 1.10 min for HMW and 1 min for LMW (according to the primer’s annealing temperature). Agarose Gel electrophoresis was performed with 1.2% agarose gel (Axygen) and 1 kb Gene RulerTM (TaKaRa), then stained by ethidium bromide stain.
Gene expression evaluation
Glutenin protein subunits extraction: The high molecular weight protein content was extracted according to the sequential procedure of Branlard and Bancel (2006), while the extraction of LMW-GS was done according to Ikeda et al. (2005). Four hexaploid Chinese cultivars seeds were extracted in addition to our samples, their glutenin proteins were used as reference for accessing the electrophoretic mobility of the HMW glutenin subunits (Yan et al., 2006), their known glutenin subunits was indicated as following:
| • | Chinese Spring CS (Null A, 1Bx7+1By8, 1Dx2+1Dy12) |
| • | CB 037 (1Ax1, 1Bx17+1By18, 1Dx2+1Dy12) |
| • | Jing 771 (1Ax1, 1Bx17+1By18, 1Dx5+1Dy10) |
| • | Jimai 20 (1Ax1, 1Bx13+1By16, 1Dx1+1Dy12) |
SDS-PAGE profile: Glutenin protein extracts (including the HMW and LMW) were separated according to the method of Singh et al. (1991) with some modifications. The concentration of separating gel was 15.0%. One-dimensional sodium dedocyl sulphate polyacrylamide gel electrophoresis (1D-SDS-PAGE) method was used according to Laemmli (1970) and was modified to get better resolution suitable to the protein patterns of glutenin subunits by Yan et al. (2002). The analysis of protein gels was done according to the known patterns of glutenin subunits of the hexaploid Chinese cultivars and the molecular weight of Protein Ruller II. The measurements were done by computer software program called Gel ANALYZER3, while the calculation of the similarity matrix and the designing of dendrogram were developed by SPSS software, version 10.
| Table 2: | The forward and reverse sequences of the specific glutenin primer pairs and their annealing temperature that used to amplify the HMW-GS and LMW-GS |
 | |
RESULTS AND DISCUSSION
The glutenin genes: Early investigations reported that each type of glutenin subunits are controlled by pairs of tightly linked loci; the HMW-GS encoded by (Glu-A1, Giu-B1, Glu-D1) loci and are located on the long arm of group 1 chromosomes (Payne et al., 1980), while the LMW-GS loci (Glu-A3, Giu-B3, Glu-D3) are found on the short arm of these chromosomes (Gupta and Shepherd, 1990; Jackson et al., 1983). No genetic recombination was observed between the HMW-GS and LMW-GS in the crosses. Some previous studies referred to that all the HMW glutenin subunits had a single locus in the three genomes (Payne and Lawrence, 1983). The subunit Mr-x is higher than Mr-y of the HMW-GS. The order of glutenin genes and their loci were illustrated in Fig. 1. On the other hand, LMW-GSs are classified into three types, LMW -i, -m and-s, based on the amino acid at their N-terminal end, which corresponded to isoleucine, methionine and serine residues, respectively (Cloutier et al., 2001; Lew et al., 1992). Types -m and s of LMW-GS were identified as protein products (Lew et al., 1992; Masci et al., 1998). Whereas, the LMW-i type was also identified as a protein product by Maruyama et al. (2004). Payne and Lawrence (1983) described the allelic variations at the HMW glutenin loci (Fig. 2).
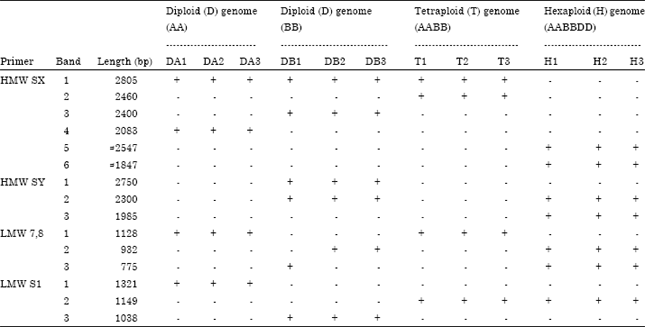
The PCR amplification patterns of the HMW and LMW alleles are shown in Fig. 3a-d. The presence, absence and the molecular sizes (bp) of detected alleles are illustrated in Table 3. Fig. 3a shows the pattern of primer HMW SX. In general, all the samples showed two alleles, but the allelic variation was detected at the size of the alleles. This variation presented differences; within the same species which exhibited complete homology, while between different species, the variations at the length were seen. The first allele has the same length (2805 bp) in 2xgenomes (AA, BB) and 4xgenome (AABB) while genome DD showed this allele with length of ≈2547 bp. The thickness of the bands of D genome was slightly higher than other bands, whereas, the second allele appeared in different length among all species but was the same within each species. The length of this allele was 2460, 2400, 2083 and ≈1847 bp in AA, BB, AABB and AABBDD genomes, respectively. Variations in the size of alleles may indicate that gene evolution caused by nucleotides polymorphisms (SNP) which cause changes in the homology situation from orthologous to paralogous genes.
The amplification of primer HMW-Y reflected the existence of subunits y of the HMW glutenin genes (Fig. 3b). The amplified alleles disappeared (silent) in all the cultivars that represented diploid (AA genome) and tetraploid species (AABB genome), while it successes to produce two alleles with the other diploid genome (BB) at length of 2750 and 2300 bp and the hexaploid genome at 2300 and 1985 bp.
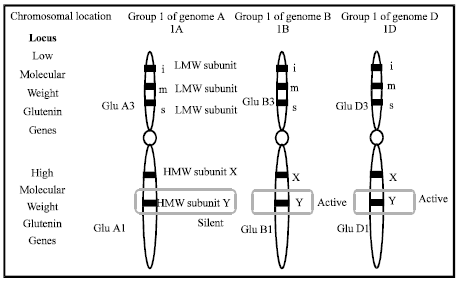 | |
| Fig. 1: | A diagram displaying the chromosomal location of high and low molecular weight glutenin genes and their subunits |
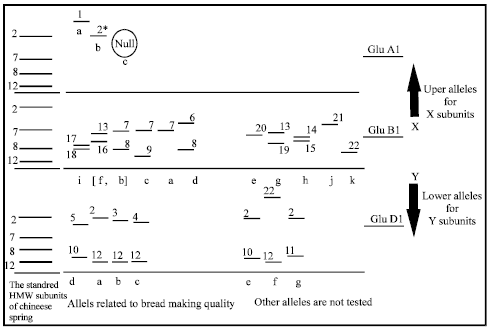 | |
| Fig. 2: | Diagram displays the SDS-PAGE allelic variation in HMW-GS at gene loci in relation to bread making quality (Payne et al., 1985), a, b, c, d, e, f, g, h, i, j and k are the allele designations (Payne and Lawerence, 1983) and their ranking is according to assessed quality |
These results disagreed with one of the fundamental studies about glutenin genes reported by Payne et al. (1981) who found that A genome y-type gene/s are always silent in hexaploid bread wheat but are active in many diploid and tetraploid wheat which support that species evolution stands behind this observation.
On the other hand, primers LMW 7,8 and LMW S were used specifically to amplify LMW loci Fig. 3c shows the amplification products of primer LMW 7,8. This primer amplified many variations of LMW alleles. These differences were realized not just among the species (genomes) but also among the individuals within the same genome. Each genome type of diploid (A and B genomes) and tetraploid (AB) exhibited one LMW allele.
 | |
| Fig. 3(a-d): | PCR amplification of high and low molecular weight glutenin genes using primers; Sx and Sy for HMW-GS and 7, 8 and SI for LMW-GS |
| Table 3: | The presence and absence of HMW and LMW glutenin alleles using specific primers for both glutenin genes |
 | |
| +: Present, -: Absent | |
However, the cultivars of hexaploid genome (ABD) showed two alleles of the same LMW locus. The tetraploid individuals agreed with the other diploid species (T. urartu, AA) in showing the same length of amplified band (1128 bp) which was the longest length. Whereas, the other diploid species (Ae. speltoides BB) showed the same allele with variation among individual samples; one of the cultivars (Turkish DB1) showed a band of 775 bp, while the other cultivars (Syrian and Iraqi) exhibited a band at 932 bp. The tetraploid at the same run exhibited two alleles (932 then 775 bp). Which indicated an addition of alleles belong to D genome. This observation was compatible with the results of Ikeda et al. (2002), who stated that s-type and m-type of LMW-GS gene are located at the Glu-D3 locus in hexaploid wheat. Moreover, most of LMW-GS genes are located in the D-genome, suggesting that the Glu-D3 locus is much larger than the Glu-B3 locus and Glu-A3 locus (Cassidy et al., 1998; Huang and Cloutier, 2008).
Primer LMW-S1 amplified one of the s-type of LMW gene, while is associated with good bread making quality (Maruyama et al., 2004). This primer distinguished in our study one clear and orthologous allele between the tetraploid and hexaploid species at 1149 bp and different allele between both diploid ancestors A and B genomes at sizes of 1321 and 1038 bp, respectively Fig. 3d. It has been suggested that conservation of orthologous sequences at the Glu-A3 locus is very limited between the A genomes (Wicker et al., 2003). In addition, the intergenic sequences in the Glu-3 regions between homoeologous A and B genomes of durum wheat are not conserved (Gao et al., 2007). Until now, little is known about the physical organization of the Glu-A3, Glu-B3 and Glu-D3 loci. Wicker et al. (2003) reported that two LMW-GS genes were separated from each other by 150 kb in T. monococcum (A genome), whereas Gao et al. (2007) found that the two paralogous LMW-GS genes were approximately 100 kb apart. The high utility of the PCR for diagnostic work makes it the method of choice for developing DNA markers to identify the glutenin genes associated with dough strength and high bread making quality in wheat species.
SDS-PAGE of glutenin proteins: According to the fundamental basics that glutenin proteins are wheat storage proteins found in the endosperms and play an important role with gliadin in determining one of the most important qualitative trait, which is bread making quality. Thus, protein profile of wheat glutenin subunits is very useful way to evaluate the expression of glutenin gene. The electrophoresis by SDS-PAGE of seed storage proteins is the standard method for glutenin subunit screening.
Wheat species; T. urartu, T. monococcum, Ae. speltoides, T. durum and T. aestivum were represented by three cultivars for each and were loaded within hexaploid Chinese cultivars of wheat (CS, CB, Jin20 and Jing771) which have known HMW glutenin subunits to identify the detected alleles in GluA1, GluB1 and GluD1 loci (Fig. 4), the reference protein ruler II (100-12 kDa) was used to determine the range of weight of the HMW-GS, gliadin and LMW-GS.
The molecular weights of HMW-GS ranged between 80-50 kDa and were seen in 4-6 patterns, while the LMW-GS was fractionated into 20 to 40 kDa and were represented in 10-12 bands (Fig. 4). This was in agreement with D’Ovidio and Masci (2004). Although the HMW-GSs represent approximately 10% and the LMW-GS about 40% of the total storage proteins in wheat endosperm, their role in determining the end-use quality of wheat flours could not be ignored, where HMW-GS is more important than the LMW-GS (Shewry and Halford, 2002). The comparison was done according to the known basis of allelic variation of the HMW-GS and the correlation between the alleles and their qualitative importance, as illustrated before through many researchers. Lawrence and Shepherd (1980) have reported an extensive variability in wheat cultivars produced by allelic variation at each locus, to relate the HMW-GS protein patterns to their qualitative importance. Based on analysis of large numbers of cultivars, a scoring system for HMW-GS has been developed by Payne et al. (1987).
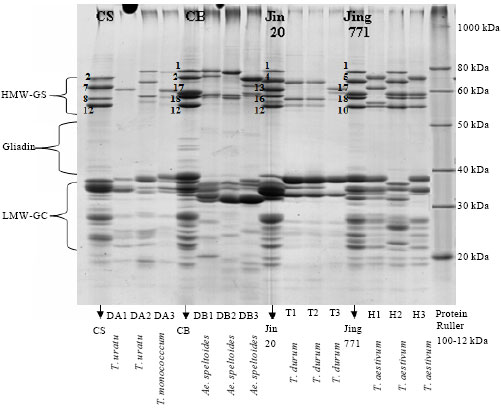 | |
| Fig. 4: | The SDS-PAGE profile of the glutenin genes (HMW, LMW), for four wheat species (three cultivars each), CS, CB, Jimai20 and Jing771 were used as referenced markers with the protein rulerII |
Through, the years, other researchers found that the same electrophoretic mobility in SDS-PAGE differs in some other features like small differences in protein sequences and surface hydrophobicity. For example Marchylo et al. (1992) found that four different alleles instead of just one, are expected for this pair (7+8, 7*+8, 7+8*, 7*+8*). Interestingly, there are contrasting effects on quality within these pairs and hence, the score originally given to the pair 7+8 is sometimes misleading. At each locus (Glu-A1, GluB1 and GluD1) there are two tightly linked HMW-GS genes; one of them is x-type -with higher molecular weight- and the other is y-type. For instance, at the Glu-1 there are subunits 1, 2* and null (there is no y allele), at the Glu-B1 locus there are B(x17+y18), B(x7+y8), B(x7+y9), B(x6+y8) and at the Glu-D1 locus, there are D(x5+y10), D(x2+y12), D(x3+y12), D(x4+y12). The presence of different allelic composition of the HMW-GS in one specific wheat variety is one of the most important genetic factors in determining the bread making quality.
According to the qualitative scores of HMW-GS which were created by Lukow et al. (1989), this study developed the quality scores of the studied cultivars as seen in Table 4. The joint occurrence of different alleles from the three loci is important in the accumulation of scores and also in determining quality (Payne, 1987). Our samples were divided into two groups, the first one is good in quality, which has high score <5 and the second one is poor quality with a score < 5. All the hexaploid cultivars belonging to T. aestivum had high quality score ranging from 5 to 10. Followed by the DB1 (Turkish) and DB2 (Syrian) that represented genome B which had scores of 9 and 6, respectively.
| Table 4: | The detected alleles for each cultivar at each locus, the calculated quality scores for all cultivars, total number of alleles per glutenin locus (GluA1, GluB1 and GluD1) and the average of allele frequency per locus |
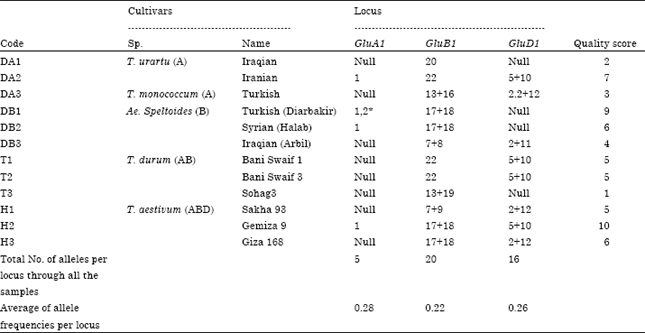 | |
However, cultivar DB3 (Iraqi) which had 4 score, reflected less quality than other cultivars of Ae. speltoides. Also two of the cultivars of T. urartu (Iraqi, DA1) T. monococcum (Yamen, DA3) were poor quality with 2 and 3 scores, respectively, while the T. urartu Iranian cultivar (DA2) has high score (7). All the cultivars of T. durum were of poor to intermediate quality, Baniswef1 and Baniswef3 expressed the same patterns and had the same quality score (5), while Sohag3 had just 1. That means the hexaploid cultivars were the best in bread making quality.
In the SDS-PAGE analysis, the calculation of molecular weight was ignored since the comparison was done according the electrophoretic mobility of the known HMW-GS of the used Chinese cultivars was much considerable than the molecular weight. It was observed that gene silencing in expression of none or only one protein from the GluA1 locus and either one of the two B genome types of loci failed to produce a subunit as allele (GluB22) which was also detected by Payne et al. (1981). The A genome y-type gene/s are always silent (GluA1c), while an Ax null allele fails to produce a subunit. The diversity in high molecular weight protein subunits is the result of gene silencing in some varieties encoding these proteins (Lawrence and Shephred, 1980).
Eight cultivars out of twelve had null alleles at GluA1 locus, while four cultivars were lacking GluD1. However, locus GluB1 was present at the cultivars. In consideration, both of DA1 and T3 lost the loci GluA1 and GluD1 together, which reflect the lowest quality score 2 and 1, respectively. Payne (1987) stated that the allelic variation at locus Glu-D1 had greater effects than other loci on bread making quality. But the present study reflects that locus GluA1 besides GluD1 have higher effect than GluD alone.
According to the previous studies of Gupta et al. (1989) and Gupta and MacRitchie (1994), subunit combination 5+10 associated with good quality of bread making, whereas subunit combination 2+12 associated with poor bread making quality. Eighteen alleles were detected and identified in three loci; 2 loci for Glu-A, 10 loci for Glu-B1 and 7 loci for Glu-D1. So locus GluB1 was the highest one. Also, Shewry et al. (1995) found so many Glu-Bx alleles present in hexaploid and tetraploid wheat as revealed by SDS-PAGE. Two alleles at Glu-A1 locus (a,1) and (b, 2*), six at Glu-B1 (b, 7+8) (c,7+9) (i, 17+18) (f, 13+16) (g,13+19) and (k, 22); and four at Glu-D1 (a,2+12) (d, 5+10) (f, 2.2+12) and (g, 2+11) were identified and displayed in Table 4.
From Table 4, we can realize that; a total of 41 alleles were obtained for all cultivars, with no common band among all the species. It can be seen that the Egyptian cultivar Gemiza9 (H2) has the highest number of alleles (five alleles) and highest score of quality (10), while DA1 has only one allele (20) at locus GluB1 and Sohag3 (T3) had the lowest quality score (1). However, Table 4 gave a conclusion about the detected alleles in each cultivar at each locus, in addition to the variability of the calculated quality score for each cultivar. The results showed that DB1 was much similar to DB2 because they have some common alleles (1, 17+18, null), while DB1 has an extra allele (2*) at locus GluA, which caused increasing in quality score than DB2. Cultivar T1 was identical to T2 which have common alleles (Null, 22, 5+10). On the other hand, H3 was similar to H1 at (Null, 2+12) and to H2 at (17+18).
Three main clusters were revealed in the dendrogram (Fig. 5) using average linkage between groups; where the first cluster contained cultivars T1, T2, T3, DA1, DA2, DB1, DB2, H2 and H3, while cultivars DB3 and H1 were found at the second cluster. Moreover, the third cluster was represented only by cultivar DA3. It was clear that the presence and absence of GluA1 alleles has the upper hand in dividing the cultivars into clusters followed by sub-clusters, then sub-sub- clusters and sub-sub-sub cluster. As seen from the dendrogram (Fig. 5).
Great variations were seen among the cultivars of progenitor genome A, cultivars DA1 and DA2 were found in the same cluster while each of them come from two different sub-clusters, whereas cultivar DA3 was found in the third main cluster. The results were reasonable because DA1 and DA2 belong to T. urartu, whereas cultivar D3 belongs to species T. monococcum. Zhang et al. (2006) from mapping some eSSRs concluded that T. aestivum and T. durum had closer relationships with T. urartu than with T. monococcum.
The progenitor B genome that represented in the cultivars which showed close relations among them; DB1 and DB2 were almost the same in [1x GluA, 17+18 GluB1, Null GluD1], while DB1 has an extra allele in GluA1 (2*). Cultivar DB3 was totally different from DB1 and DB2, thus it was found in another cluster.
Cultivars T1, T2 and T3 belonged to the same main cluster. The identical cultivars (T1 and T2) were similar to T3 with only 30%, the absence of GluA1 alleles was the common property among them and this observation is matched with the highest average of allele frequency (0.28) that scored by locus GluA1, which followed by locus GluD1 (0.26) and the lowest one was locus GluB1 (0.22) (Table 4).
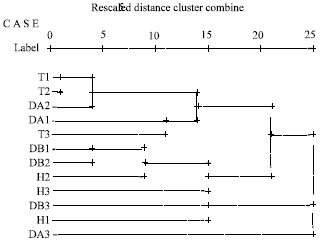 | |
| Fig. 5: | The dendrogram showing the relationships among the studied cultivars according to the protein patterns of SDS-PAGE |
The presence of GluA1 controlled the clustering of hexaploid cultivars; H1 cultivar was found in the second cluster (has allele 1 at locus GluA1). However the other hexaploids (H2 and H3) belonged to the first one.
The relation between tetraploid and hexaploid and their progenitors showed that; T1 and T2 are related to DA2 because they had common alleles (22 GluB1 and 5+10 GluD1). T3 is related to DA1 because both of them have null alleles at GluA1 and GluD1 loci. And the existence of DA3 alone in the third cluster may be due to that none of these T. durum or T. aestivum cultivars have a common ancestor belong to T. monococcum. The hexaploid H2 was found at the same cluster with DB1 and DB2, which might be due to that they may have same progenitor of Ae. Speltoides since they had common GluA1 and GluB1 loci 1 and 17+18, respectively, whereas, cultivars H1 and H3 were much similar to DB3. Generally speaking, the results of the HMW glutenin gene and its expression (protein profile) proved the compatibility between them. The absence of y subunit of the HMW glutenin gene in diploid species (A genome) and tetraploid individuals, on the molecular level was reflected as expected on silencing expression of y subunits in the genomes mentioned above (Fig. 4, Table 4).
Fufa et al. (2005) reported that the genetic diversity estimates based on seed storage protein were lowest because of the major determinants of end-use quality, which is a highly selected trait. It is therefore concluded that seed storage protein profiles could be useful markers in cultivar identification, registration of new varieties, pedigree analysis and in the studies of genetic diversity and classification of adapted cultivars, thereby improving the efficiency of wheat breeding programs in cultivar development.
CONCLUSION
This study aimed to detect some glutenin genes in different wheat species by designing four specific primers of HMW (HMW SX and HMW SY) and LMW (LMW 7,8 and LMW S1),these primers amplified both the HMWGS and LMWGS to identify and compare between according to their bread making quality.
The glutenin protein subunits were extracted from the samples comparing with referenced Chinese hexaploid cultivars. The 1D-SDS-PAGE method was used, to fraction the entire expressed proteins of HMW and LWM glutenin genes to get phylogenetic relationships among the diploid, tetraploid and hexaploid wheat genomes. Generally spreading, the results showed that, the absence or presence of GluA1 allele is responsible for the clustering of different cultivars between and within species. In addition, there was concordance between resulted genomic and proteomics analysis of the wheat genomes.
REFERENCES
- Cassidy, B.G., J. Dvorak and O.D. Anderson, 1998. The wheat low-molecular-weight glutenin genes: Characterization of six genes and progress in understanding gene family structure. Theor. Applied Genet., 96: 743-750.
CrossRefDirect Link - Cloutier, S., C. Rampitsch, G.A. Penner and O.M. Lukow, 2001. Cloning and expression of a LMW-i glutenin gene. J. Cereal Sci., 33: 143-154.
CrossRefDirect Link - D'Ovidio, R. and S. Masci, 2004. The low-molecular-weight glutenin subunits of wheat gluten. J. Cereal Sci., 39: 321-339.
CrossRefDirect Link - Fufa, H., P.S. Baenziger, B.S. Beecher, I. Dweikat, R.A. Graybosch and K.M. Eskridge, 2005. Comparison of phenotypic and molecular marker-based classifications of hard red winter wheat cultivars. Euphytica, 145: 133-146.
CrossRefDirect Link - Gao, S., Y.Q. Gu, J. Wu, D. Coleman-Derr and N. Huo et al., 2007. Rapid evolution and complex structural organization in genomic regions harboring multiple prolamin genes in the polyploid wheat genome. Plant Mol. Biol., 65: 189-203.
CrossRefPubMedDirect Link - Gupta, R.B. and K.W. Shepherd, 1990. Two-step one-dimensional SDS-PAGE analysis of LMW subunits of glutelin. Theor. Applied Genet., 80: 65-74.
CrossRef - Gupta, R.B., N.K. Singh and K.W. Shepherd, 1989. The cumulative effect of allelic variation in LMW and HMW glutenin subunits on dough properties in the progeny of two bread wheats. Theor. Applied Genet., 77: 57-64.
CrossRef - Huang, X.Q. and S. Cloutier, 2008. Molecular characterization and genomic organization of low molecular weight glutenin subunit genes at the Glu-3 loci in hexaploid wheat (Triticum aestivum L.). Theor. Applied Genet., 116: 953-966.
CrossRefPubMedDirect Link - Ikeda, T.M., T. Nagamine, H. Fukuoka and H. Yano, 2002. Identification of new low-molecular-weight glutenin subunit genes in wheat. Theor. Applied Genet., 104: 680-687.
CrossRefPubMedDirect Link - Ikeda, T.M., N. Ohnishi, T. Nagamine, S. Oda, T. Hisatomi and H. Yano, 2005. Identification of new puroindoline genotypes and their relationship to flour texture among wheat cultivars. J. Cereal Sci., 41: 1-6.
CrossRefDirect Link - Jiang, Q.T., Y.M. Wei, F. Wang, J.R. Wang, Z.H. Yan and Y.L. Zheng, 2009. Characterization and comparative analysis of HMW glutenin 1Ay alleles with differential expressions. BMC Plant Biol., Vol. 9.
CrossRefDirect Link - Johansson, E., G. Svensson and W.K. Heneen, 1995. Composition of High-Molecular-Weight glutenin subunits in Swedish wheats. Acta Agric. Scand. Sect. B: Soil Plant Sci., 45: 112-117.
CrossRefDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Lawrence, G.J. and K.W. Shepherd, 1980. Variation in glutenin protein subunits of wheat. Aust. J. Biol. Sci., 33: 221-233.
Direct Link - Lew, E.J.L., D.D. Kuzmicky and D.D. Kasarda, 1992. Characterization of low molecular weight glutenin subunits by reversed-phase high-performance liquid chromatography, sodium dodecyl sulfate-polyacrylamide gel electrophoresis and N-terminal amino acid sequencing. Cereal Chem., 69: 508-515.
Direct Link - Liu, Y., Z.Y. Xiong, Y.G. He, R.S. Peter and G.Y. He, 2007. Genetic diversity of HMW glutenin subunit in Chinese common wheat (Triticum aestivum L.) landraces from Hubei province. Genet. Resour. Crop. Evol., 54: 865-874.
CrossRef - Lukow, O.M., P.I. Payne and R. Tkachuk, 1989. The HMW glutenin subunit composition of Canadian wheat cultivars and their association with bread-making quality. J. Sci. Food Agric., 46: 451-460.
CrossRefDirect Link - Marchylo, B.A., O.M. Lukow and J.E. Kruger, 1992. Quantitative variation in high molecular weight glutenin subunit 7 in some Canadian wheats. J. Cereal Sci., 5: 29-37.
CrossRefDirect Link - Maruyama, F.W., K. Takata, Z. Nishio, T. Tabiki and E. Yahata et al., 2004. Identification of low-molecular weight glutenin subunits of wheat associated with bread-making quality. Plant Breed., 123: 355-360.
CrossRefDirect Link - Masci, S., R. D'Ovidio, D. Lafiandra and D.D. Kasarda, 1998. Characterization of a low-molecular-weight glutenin subunit from bread wheat and corresponding protenin that represents a major subunit of the glutenin polymer. Plant Physiol., 117: 1147-1158.
Direct Link - Payne, P.I., C.N. Law and E.E. Mudd, 1980. Control by homologous group 1 chromosomes of the high-molecular-weight subunits of glutenin, a major protein of wheat endosperm. Theor. Applied Genet., 58: 113-120.
CrossRef - Payne, P.I., 1987. Genetics of wheat storage proteins and the effect of allelic variation on bread-making quality. Annu. Rev. Plant Physiol., 38: 141-153.
CrossRefDirect Link - Payne, P.I., M.A. Nightingale, A.F. Krttiger and L.M. Holt, 1987. The relationship between HMW glutenin subunit composition and the bread-making quality of British-grown wheat varieties. J. Sci. Food Agric., 40: 51-65.
CrossRefDirect Link - Payne, P.I., K.G. Corfield, L.M. Holt and J.A. Blackman, 1981. Correlations between the inheritance of certain high-molecular-weight subunits of glutenin and breadmaking quality in progenies of six crosses of bread wheat. J. Sci. Food Agric., 32: 51-60.
CrossRef - Payne, P.I., L.M. Holt, M.G. Jarvis and E.A. Jackson, 1985. Two-dimensional fractionation of the endosperm proteins of bread wheat (Triticum aestivum): Biochemical and genetic studies. Cereal Chem., 62: 319-326.
Direct Link - Saiki, R.K., S. Scharf, F. Faloona, K.B. Mullis, G.T. Horn, H.A. Erlich and N. Arnheim, 1985. Enzymatic amplification of beta-globin genomic sequences and restriction site analysis for diagnosis of sickle cell anemia. Science, 230: 1350-1354.
CrossRefDirect Link - Shewry, P.R. and N.G. Halford, 2002. Cereal seed storage proteins: Structures, properties and role in grain utilization. J. Exp. Bot., 53: 947-958.
CrossRefPubMedDirect Link - Shewry, P.R. and A.S. Tatham, 1997. Disulphide bonds in wheat gluten proteins. J. Cereal Sci., 25: 207-227.
CrossRef - Shewry, P.R., A.S. Tatham, F. Barro, P. Barcelo and P. Lazzeri, 1995. Biotechnology of bread-making: unraveling and manipulating the multi-protein gluten complex. Nal. Biotechnol., 13: 1185-1190.
CrossRefDirect Link - Singh, N.K., K.W. Shepherd and G.B. Cornish, 1991. A simplified SDS-PAGE procedure for separating LMW subunits of glutenin. J. Cereal Sci., 14: 203-208.
Direct Link - Wan, Y., D. Wang, P.R. Shewry and N.G. Halford, 2002. Isolation and characterization of five novel high molecular weight subunit of glutenin genes from Triticum timopheevi and Aegilops cylindrica. Theor. Applied Genet., 104: 828-839.
CrossRefDirect Link - Wicker, T., N. Yahiaoui, R. Guyot, E. Schlagenhauf, Z.D. Liu, J. Dubcovsky and B. Keller, 2003. Rapid genome divergence at orthologous low molecular weight glutenin loci of the A and Am genomes of wheat. Plant Cell, 15: 1186-1197.
CrossRefDirect Link - Yan, Y., Y. Zhang, Q. Li, J. Zheng and X. An et al., 2006. Molecular characterization and phylogenetic analysis of a novel glutenin gene (Dy10.1t) from Aegilops tauschii. Genome, 49: 735-745.
CrossRefPubMedDirect Link - Yan, Z.H., S.F. Dai, D.C. Liu, Y.M. Wei and Y.L. Zheng, 2008. Isolation and characterization of a novel HMW-GS Glu-Dx allele from Tibet bread wheat landrace. Cereal Res. Commun., 36: 523-531.
CrossRefDirect Link - Yan, Z.H., Y.F. Wan, K.F. Liu, Y.L. Zheng and D.W. Wang, 2002. Identification of a novel HMW gultenin subunit and comparison of its amino acid sequence with those of homologous subunits. Chin. Sci. Bull., 47: 220-225.
CrossRefDirect Link - Zhang, L.Y., C. Ravel, M. Bernard, F. Balfourier, P. Leroy, C. Feuillet and P. Sourdille, 2006. Transferable bread wheat EST-SSRs can be useful for phylogenetic studies among the Triticeae species. Theor. Applied Genet., 113: 407-418.
CrossRefDirect Link