
I. P. Ogbuewu
Department of Animal Science and Technology, P.M.B 1526, Owerri, Imo State, Nigeria
A. A. Omede
Department of Animal Science and Technology, P.M.B 1526, Owerri, Imo State, Nigeria
O. K. Chukwuka
Department of Animal Science and Technology, P.M.B 1526, Owerri, Imo State, Nigeria
O. O.M. Iheshiulor
Department of Animal Science and Technology, P.M.B 1526, Owerri, Imo State, Nigeria
M. C. Uchegbu
Department of Animal Science and Technology, P.M.B 1526, Owerri, Imo State, Nigeria
A. C. Udebuani
Department of Biotechnology, Federal University of Technology, P.M.B 1526, Owerri, Imo State, Nigeria
B. C. Ekenyem
Department of Biotechnology, Federal University of Technology, P.M.B 1526, Owerri, Imo State, Nigeria
I. C. Okoli
Department of Animal Science and Technology, P.M.B 1526, Owerri, Imo State, Nigeria
M. U. Iloeje
Department of Animal Science and Technology, P.M.B 1526, Owerri, Imo State, Nigeria
International Journal of Agricultural Research
Year: 2010 | Volume: 5 | Issue: 12 | Page No.: 1084-1099







ABSTRACT
Phytoestrogens are naturally occurring phytochemicals found in plants and plant products, which are structurally and functionally similar to human or animal estrogens (17β-oestradiol) or synthetic estrogens such as diethylstilboestrol. The principal phytoestrogens are the isoflavones, which are similar to 17β-oestradiol and then the lignans derived from precursors in the diet by the gut microflora. Isoflavones are members of the flavonoid family, which are in turn members of the larger group of plant constituents known as polyphenols. The principle isoflavones in soy are genistein, daidzein and their metabolites. Since, soy isoflavone are naturally occurring non-steroidal compounds, which are structurally similar to endogenous gonadal steroid 17β-estradiol, they possess the ability to cause estrogenic or/ and antiestrogenic effects and therefore could trigger estrogen dependent physiological responses. As result of these actions, there is currently much interest within the scientific community regarding clinical benefits of soy based isoflavone.
PDF Abstract XML References Citation
Received: March 14, 2010;
Accepted: May 21, 2010;
Published: July 17, 2010
How to cite this article
I. P. Ogbuewu, A. A. Omede, O. K. Chukwuka, O. O.M. Iheshiulor, M. C. Uchegbu, A. C. Udebuani, B. C. Ekenyem, I. C. Okoli and M. U. Iloeje, 2010. The Overview of the Chemistry, Health Benefits and the Potential Threats Associated with Prolonged Exposure to Dietary Soy Isoflavones. International Journal of Agricultural Research, 5: 1084-1099.
DOI: 10.3923/ijar.2010.1084.1099
URL: https://scialert.net/abstract/?doi=ijar.2010.1084.1099
DOI: 10.3923/ijar.2010.1084.1099
URL: https://scialert.net/abstract/?doi=ijar.2010.1084.1099
INTRODUCTION
Soy and soy products are widely consumed throughout the world. In Asian countries, soy has been an important part of the diet for more than a thousand years. The consumption of soy products is estimated to be highest among the Japanese population, with the levels of isoflavones reaching 200 mg day-1 (Cassidy et al., 1994). In other parts of Asia, the diet provides 25-40 mg of total isoflavones per day, whereas in the western countries less than 5 mg day-1 is consumed (Murphy, 1982; Coward et al., 1993; Setchell and Cole, 2003).
Isoflavones can effectively and efficiently modulate estrogen levels in humans and animals, because of its ability to bind estrogen receptor (α and β) (Setchell and Adlercreutz, 1988; Adlercreutz, 1997; Kuiper et al., 1997; Setchell and Cassidy, 1999). They are of clinical value in estrogen sensitive conditions such as breast cancer, cystic ovaries and endometriosis among women (Verger and Leblane, 2003). These estrogen mimics have been shown in animal models and in limited clinical investigations to be protective in the prevention of cardiovascular disease, osteoporosis and hormone-dependent cancers (Setchell, 1998; Setchell and Cassidy, 1999; Middleton et al., 2000; Adlercreutz et al., 2000).
As a result of the numerous clinical benefits of soy isoflavones, there is much interest regarding the increased use of soy based products in animals and man as a component of healthy diets. This review is an attempt to present the general picture of the clinical and dietary significance of soy isoflavone in humans and animals.
Phytoestrogens
Phytoestrogens are naturally occurring phytochemicals found in plants and plant products, which are structurally and functionally similar to human or animal estrogens (17β-oestradiol) or synthetic estrogens such as diethystilboestrol (Burton and Wells, 2002; FSA, 2002). The principle phytoestrogens are the Isoflavones, which are similar to 17β-oestradiol and its examples are genistein, daidzein, formononetin, biochanin A and equol (Yildiz, 2005; Boue et al., 2000) and then the lignans, example: enterolactone and enterodiol derived from precursors in the diet by the gut microflora (Phipps et al., 1993).
There are also the mycotoxins derived from fungal moulds or grain (zearalenone) as reported (Murkies et al., 1995). Among these phytoestrogens, only zearalenmore has been isolated for use in animal production as zeranol and it gives good results but has an effect on genital formation and the level of thyroxine in plasma (Leopold et al., 1976), while others are taken up directly in the pasture. On the other hand, isoflavones have received the greatest attention with regard to female genital tract pathology.
Because of their structural similarity to endogenous estrogens, phytoestrogens bind to both Estrogens Receptors (ER) alpha and Beta (but more strongly to ER-beta) and exert estrogen- like effects (FSA, 2002; Ranich et al., 2001). Phytoestrogens are present as glycosides in legumes, grains, nuts and other fiber rich foods (Adlercreutz, 1990; Adlercreutz et al., 1991) and are present in the plasma and urine of both humans and animals eating a diet rich in such foods (Adlercreutz et al., 1992; Maskarinec et al., 2001) although there is a pronounced variation between individuals (Franke and Custer, 1994).
Isoflavones
Isoflavones of nutritional interest are substituted derivatives of isoflavone, being related to the parent by the replacement of two or three hydrogen atoms with hydroxyl groups as reported by Adlercreutz (1997) (Fig. 1). In genistein X1 and X2 are substituted with hydroxyl group (OH) whereas in daidzein X1 and X2 is substituted with hydroxyl group (OH) and hydrogen (H), respectively. The parent isoflavone is of no nutritional interest. Isoflavone differs from flavone (2-phenyl-4H-1-benzopyr-4-one) in location of the phenyl group.
The major isoflavones present in plant based foods are genistein, daidzein, glycetin, biochanin A and formononetin. Isoflavones are often present as glucoside conjugates, glucones in plants and foods which undergo metabolic transformation in the gut to the functional molecules, such as genistein and daidzein (Anderson et al., 1995).
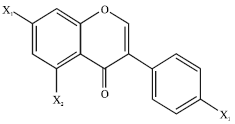 | |
| Fig. 1: | The basic structure of parent isoflavones |
| Table 1: | Isoflavones most commonly found in foodstuffs |
 | |
| Aglucones: Unconjugated to glucose, Glucones: or glucosides, conjugated to glucose, Acetylglucones: or acetylglucosides, esterified with an acetyl group, Malonylgucones: or Malonylglucoside, esterified with a malonyl group | |
Major metabolic changes include the conversion of genistin to genistein and daidzin to daidzein. The former molecules are called glucones (glycones) because they have a glucose molecule attached to them, whereas the later molecules are referred to as aglucones because the glucose molecules are enzymatically removed (Table 1).
Aglucones function in plants as antioxidants and as repellants of animals’ phytoalexins (Anderson et al., 1995; Fallon, 2006), while the glucones apparently serve only as storage molecules. Barnes et al. (1994) added that the glucose group is often esterified with an acetyl-or malonyl group to form acetyl-or malonylglucosides, while it will be important to state that Biochanin A and formononetin are derivatives of genistein and daidzein that have additional methyl (CH3) group. Isoflavones, isolated from the roots and bark of some plants and trees (Shirataki et al., 1999; Bojase et al., 2002) are compounds of similar structure to isoflavones and therefore, may have similar estrogenic properties. However, they have not been identified in food and their estrogenic properties have been established.
Biosynthesis of soy isoflavones: Isoflavones are produced via a branch of the general phenylpropanoid pathway that produces flavonoid compounds in higher plants. The phenylpropanoid pathway begins from the amino acid phenylalanine and an intermediate of the pathway, naringenin, is sequentially converted into the isoflavone genistein by two legume-specific enzymes, isoflavone synthase and a dehydratase. Similarly, another intermediate naringenin chalcone is converted to the isoflavone daidzein by sequential action of three legume-specific enzymes: chalcone reductase, type II chalcone isomerase and isoflavone synthase.
Water Solubility of Isoflavones
Isoflavones are low molecular weight hydrophobic compounds. Conjugation to glucose, glucuromide or sulphate groups increases water solubility. Acetylation or malonylation of glucose conjugates and methylation of the isoflavone moiety will alter water solubility chemical stability.
Under acidic conditions, the glucones can be deconjugated to give aglucones. Whilst under acidic or basic condition the acetyl- and malonyl groups can also be removed. In addition, malonyl groups can decarboxylate (lose CO2) thus yielding acetyl groups. In the body, enzymes in the gut can carry out these reactions during metabolism (FSA, 2002).
Coumestans
Compared with isoflavones, coumestans have been less studied although Burton and Wells (2002) claimed them to be the most potent of the phytoestrogens. They are structurally similar to isoflavones and possess similar physical and chemical properties (Humfrey, 1998). The structure of coumestrol, the coumestan most commonly found in foods, is shown in Fig. 2.
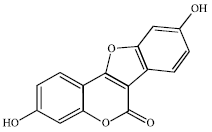 | |
| Fig. 2: | The chemical structure of coumestrol, which has been identified in some foods |
Prenylated Flavonoids
A number of prenylated flavonoids with estrogenic properties have also been identified (Kitaoka et al., 1998; Miyamoto et al., 1998; Milligan et al., 1999):
| • | 8-prenylnaringenin |
| • | 6-prenylnaringenin |
| • | Xanthohumol |
| • | Isoxanthohumol |
These compounds are also structurally similar to the isoflavones but substituted with a prenyl group (B) and the phenol ring (A) is orientated in a different direction. The presence of the prenyl group makes these compounds less water soluble than the isoflavones.
Lignans
Members of the lignan group of phytoestrogens are defined as possessing the 2, 3- substituted di-1-4 benzylbutance structure (FSA, 2002). The principal lignans identified in food are:
| • | Larciciresinol |
| • | Isolariciresinol |
| • | Matairesinol |
| • | Secoisolariciresinol |
The form in which the lignans occur in foods is unknown but it has been suggested they are present as long-chain polymers (Liggins et al., 2000). For this reason, isolation of these compounds from plants and foods requires chemical treatment after which they are in the form of aglucones or glucones (Liggins et al., 2000). Lignans are thought not be estrogenic themselves, but are converted to the oestrogenic compounds, enterolactone and enterodiol by the gut microflora. Liggins et al. (2000) identified in food include lariciresinol, isolariciresinol, secoisolariciresinol and matairesinol and the metabolities enterodiol and enterolactone
Dietary Source and Intake of Soy Isoflavone
Estimation of the intake of isoflavones is clearly an important element in the assessment of their potential importance to human health. Isoflavones exist primarily in soybeans and in most soy foods as a complex mixture of glucoside conjugates (Setchell et al., 2002; Omede, 2003). Soy is a low cost source of protein that has been consumed in Asian nations for many centuries. The development of comprehensive computer based databases of the phytochemical content of food is commencing and this, together with more accurate analytical methods, will assist in more accurately estimating dietary intake of isoflavones and hence their role in human health.
Bioavailability of Soy Isoflavone
Soy isoflavones is a complex mixture of glucoside conjugates that are not bioavailable in this form (Setchell et al., 2002). After ingestion, the isoflavone glucosides are hydrolyzed by bacterial ß-glucosidases releasing the aglycones (McMahon et al., 1997; Day et al., 1998; Setchell et al., 2002), which are then either absorbed directly or further metabolized by intestinal microflora in the large intestine into other metabolites, including equol (Axelson et al., 1982, 1984; Setchell et al., 2002) and O-desmethylangolensin (Bannwart et al., 1984; Joannou et al., 1995).
The bioavailability and pharmacokinetics of isoflavones are mainly influenced by the type of food (Setchell et al., 2002). Isoflavones of liquid foods such as soy milk gives faster absorption. Aglycones present in fermented soy foods are absorbed more rapidly than isoflavones glucosides. Isoflavones glycosides are not absorbed intact across the enterocyte of healthy adults and their bioavailability requires initial hydrolysis of the sugar moiety by intestinal ß-glucosidases for uptake to the peripheral circulation (Setchell et al., 2002).
Effects of Soybean based Soy Isoflavones
The issue of the safety of use of phytoestrogenic diets in animal and man in relation to isoflavone toxicity and benefit considerations has been the subject of discussion (Fallon, 2006). Authors argue and state that they are not convinced that long history of apparent safe use of phytoestrogenic products can provide confidence that they are indeed without risk. Interestingly, as the argument for the potential adverse effects of phytoestrogens is gaining support so also another group are rallying round phytoestrogens with the view they have potential beneficial effects both on man and animals as follows:
Digestion, Absorption and Utilization of Isoflavone
Within the lumen of the gastro intestinal tract, bacteria have the enzymatic capacity to metabolically modify the aglucone isoflavone to other structures, such as conversion of formononetin to daidzein and then daidzein to equol. These modified isoflavones i.e., metabolites are handled similarly to the original glycones in terms of absorption and distribution, while the degradation products of the aglycone molecules are eventually excreted in either the feces or urine.
Bacterial enzymes known as glycosidase digest the glycones within the fluid environment of both the lower small intestine and the large intestine. The released aglycones and their metabolites solubilize in the micelles that are derived from the bile that is secreted into the GI tract by the liver (Miksicek, 1994). The aglycones and their metabolites are absorbed like other fast-soluble molecules in diverse foods. Once the lipid soluble aglycones become incorporated in micelles within the gut lumen, they can migrate to and be taken up at the luminal surface of the absorbing epithelial cells lining the gut wall. Micelles permit their contents of fat –soluble molecules, including aglycones to enter the cells by passive diffusion. Thereafter, the aglycones and their metabolites such as equol are repackaged forming chylomicrons for subsequent release by these absorbing cells to the lymphatics and then the blood. This last step completes the process of absorption of isoflavones (Miksicek, 1994).
Equol is apparently made in large amounts from daidzein within the gut lumen and is recycled in the enterohepatic circulation many times before it is excreted in the feces or urine (Xu et al., 1995). In this way their concentrations in the blood are able to increase until a steady level is achieved so long as isoflavones containing feeds are consumed on a daily basis. Once in the cells of the different tissues of the body, the isoflavones act as weak agonists or antagonists of estrogens using estradiol as the model estrogen molecule.
Cellular mechanisms of action: Phytoestrogens act on cells in a similar way as estradiol, though are not as potent in their actions (Burton and Wells, 2002). Possible reasons being first, that when animals consume phytoestrogenic feeds, the isoflavones has to be digested, modified by the gut flora, absorbed and distributed around the body via chylomicrons. Then the isoflavones are thought to re-circulate in the enterohepatic circulation numerous times before a steady-state blood concentration is achieved. Secondly, the affinity of the isoflavone with Estrogen Receptors (ER) cells is not as great as that of estradiol (Miksicek, 1994).
Whitten et al. (1994) and Burton and Wells (2002) suggested that phytoestrogens may exert their biological activity by
| • | Mimicking the action of endogenous estrogens |
| • | Acting as estrogen antagonists |
| • | Altering the pattern of synthesis and metabolism of endogenous hormones |
| • | Modifying hormone receptor values |
In addition it has been shown that there is a positive correlation between fiber intake, urinary excretion of enterolactone and serum Steroid Hormone Binding Globulin (SHBG) and a negative correlation between urinary excretion of enterolactone and serum estradiol, thus, the ingestion of phytoestrogens stimulates the hepatic synthesis of SHBG and indirectly reduces the amount of free biologically active estradiol in the serum. Therefore, in the absence of estrogen, isoflavones have a weakly estrogenic effect, but may exhibit an antioestrogenic effect when estrogen is present (Adams, 1995; Cline et al., 1996).
However, it must be remembered that phytoestrogens may exert biological activity by other mechanisms, for example, the isoflavone genistein is a potent selective inhibitor of tyrosine kinase in both man (Morrison et al., 1996) and rat myometrial cells (Palmier et al., 1996) and as well genistein, daidzein and biochanin A inhibited thyroid peroxidase (TPO), an enzyme involved in the synthesis of T3 and T4 (tri-iodo thyronine and thyroxine, respectively) as reported by FSA (2002) and Divi et al. (1997).
Isoflavones and Estrogen Receptor Sites
Isoflavones bind competitively to both estrogens α (ERα) and estrogen β (ERβ) receptors and activate them (Kuiper et al., 1997). The ERα is called a classic ER and has been known for decades, while ERβ was described and characterized for the first time only a few years ago (Mosselman et al., 1996). It binds to ERα with a ten times lower affinity than estradiol17β, but its dissociation is close to that of estradiol-17β (Scarlata and Miksicek, 1995). Daidzein has a higher binding affinity to ER than its methoxy derivative, formononetin (Shutt and Cox, 1972). It has been suggested that hydroxylation is necessary for a flavonoid to have estrogenic activity (Miksicek, 1995).
The flavonoids with hydroxyl substituents at 4’ and 7 positions are estrogenic and an additional hydroxyl group at the 5 position-like that possessed by genistein-increases estrogenic activity (Miksicek, 1995). On the other hand, if a flavonoid has more than four hydroxyl substituents, (such as flavonol quercetin) or has a 4’-methoxylated substituents (such as hesperitin), the estrogenic activity is abolished (Miksicek, 1995). However, the affinity of plant derived estrogens bind competitively to both estrogen α (ERα) and estrogen β (ERβ) receptors and activate them appeared to be linked to several possible mechanisms (Gyorgy et al., 1964).
Soybean (Glycine sp.)
Products may contain up to 0.25% isoflavones. The major isoflavones in soybean are genistein (~60%),daidzein(~25%) and glycetin (~15%) based diets have been reported to cause estrogenic effects in swine and laboratory animals but there are no reports of effects in ruminants. In addition, soy plants use isoflavones and their derivatives as phytoalexin compounds to ward off disease-causing pathogenic fungi and other microbes.
Soy Isoflavones and Cancer
The incidence of breast, endometrial and ovarian cancer is lower in Asia and Eastern Europe than in Western countries (Rose et al., 1986). Migrants from Asia who maintain their traditional diet have a decreased risk even when living in western countries (Kolonel, 1988), whereas the increased risk of these diseases follows a change towards a westernized diet (Lee et al., 1991). An increased soy intake, for example, is associated with reduced breast cancer risk in both pre- and post-menopausal women (Wu et al., 1996) and with lowered prostate cancer risk in men (Strom et al., 1999).
A soy protein diet has been reported by Bylund et al. (2000) inhibits the growth of prostate adenocarcinoma in mice. In rats, three subcutaneous injections of genistein administered neonatally protect against mammary cancer (Lamartiniere et al., 1995).
A mechanism by which researchers think soy isoflavones could potentially play a role in reduction of prostate cancer risk is that genistein has shown to inhibit the growth of both androgen-dependent and androgen-independent prostate cancer cells in vitro. A second possibility is that the estrogenic effects of isoflavones have a protective role in inhibitinHumfreyg metastatic prostate cancer.
Soy Isoflavones and Cardiovascular Diseases
Clinical studies have focused on the effects of consumption of isoflavone rich foods such as soybean on known risk factors for cardiovascular disease, usually the levels of cholesterol and Low Density Lipoproteins (LDL) in plasma, as well as the ability of LDL to withstand oxidation ex vivo, since oxidation of LDL is recognized as an important process in the initiation of Atherosclerosis (King, 2000). Diets containing soy protein lower plasma cholesterol concentrations in humans and experimental animals (Anderson et al., 1995; Carroll and Kurowska, 1995; Potter, 1996; Sirtori and Lovati, 2001; Ogbuewu et al., 2010). Early studies in rabbits fed cholesterol-free diets demonstrated that isolated soy protein was one of the most hypocholesterolemic proteins of various dietary plant and animal proteins (Huff et al., 1977; Huff et al., 1982).
In rabbits, soy protein enhanced plasma cholesterol turnover, increased fecal neutral and acidic steroid secretion, decreased cholesterol reabsorption and reduced the secretion of apoB into plasma (Huff and Carroll, 1980; Khosla et al., 1989; Samman et al., 1989). Soy protein has been linked to up regulation of the Low Density Lipoprotein (LDL) receptor in human studies (Lovati et al., 1987)
Epidemiologic studies have demonstrated a reduced rate of mortality due to coronary heart disease in Japanese populations consuming a traditional Japanese diet compared to a western diet (Kagan et al., 1974). Expatriate Japanese living in the United Kingdom have higher blood pressure and cholesterol levels and lower triglyceride levels than the Japanese still living in Japan (Robinson et al., 1995), which suggests that these differences are not of genetic origin but may be due to diet.
Isoflavones and Antioxidants
Studies in non-human primates (Adams et al., 2005; Clarkson et al., 1998) and rabbits (Yamakoshi et al., 2000) have demonstrated retardation of atherogenesis during dietary isoflavone phytoestrogen administration. In the latter study, increased anti-oxidant protection of LDL and an athero-protective effect was observed, but these did not correlate with the serum lipid profile of the animals (Yamakoshi et al., 2000). Soy-isoflavones are known to be powerful antioxidants in lipid aqueous systems in vitro (Kapiotis et al., 1997). The observed decrease liver cholesterol level of rabbit bucks administered soymilk could be attributed in part by the antioxidant activity of soy isoflavones (Ogbuewu et al., 2010). The antioxidant mechanism of phenolic compounds, such as genistein, is dependent on the hydrogen atom or electron transfer from the phenolic C-3 -hydroxyl group (Wright et al., 2001). Genistein as a free radical scavenger donates the hydrogen atoms of the phenolic hydroxyl groups (Heim et al., 2002).
There are, however, other possible effects of genistein that may be physiologically relevant in suppressing the oxidative stress and related inflammation in the vascular intima. In studies with macrophages, genistein inhibited the activation of nuclear factor κB (NKκB), which is inducible by oxidative stress and regulates the expression of genes involved in immune and inflammatory responses (Choi et al., 2000). Genistein increases the expression of antioxidant enzymes such as glutathione peroxidase in human prostate cancer cells and protects these cells against oxidative DNA damage in vitro (Raschke et al., 2006).
Isoflavones and Bone Metabolism
Osteoporosis is related to aging and especially to the menopause. After the cessation of ovarian function, women begin to lose their bone mass. The incidence of osteoporosis differs within populations and according report of WHO (1994) the incidence is lower in Asian women than in western women. One of the reasons for this could be the dietary differences between the areas, which are partly related to the consumption of soy products.
Soy isoflavones have been shown to attenuate bone loss in perimenopausal women (Alekel et al., 2000) and in ovariectomized (OVX) rats (Arjmandi et al., 1998). This may be due to enhanced bone formation rather than to slowed bone resorption (Alekel et al., 1998). Although both genistein and daidzein are effective in preventing bone loss, daidzein is the more potent of these two compounds (Picherit et al., 2000). To sum up, genistein and daidzein have estrogenic effects, which may be beneficial in treating menopausal symptoms.
Soy Isoflavones and Endogenous Reproductive Hormones
Isoflavones act as estrogen receptor act as ER agonists or antagonists, depending on the hormonal status of the animal or man. Isoflavonoids at concentrations 100-1000 times higher than that of estradiol-17α have been considered to compete with endogenous mammalian estrogens, to bind ER and to prevent estrogen-stimulated growth in mammals (Adlercreutz et al., 1995). It is therefore possible that the consumption of a diet rich in plant-derived estrogens could affect endogenic hormone production. The mid-cycle peaks of the luteinizing hormone and the follicle stimulating hormone are suppressed (Cassidy et al., 1995) or sometimes unaffected (Lu et al., 2000) and in premenopausal women the length of the follicular phase of the menstrual cycle is increased during an isoflavone-rich diet (Cassidy et al., 1995; Lu et al., 1996). The serum estradiol-17β concentration is unaffected (Cassidy et al., 1995; Honore et al., 1997) or decreased (Lu et al., 1996, 2000), the progesterone level is decreased (Lu et al., 1996) and the serum testosterone concentration is unaffected (Honore et al., 1997) or reduced (Strauss et al., 1998) by dietary isoflavone supplementation.
Potential Reproductive Toxicity of Isoflavones
The issue of the safety of use of phytohormones in the diets in animal production in relation to isoflavone toxicity and benefit considerations has been the subject of a recently discussion (Fallon, 2006; Omede et al., 2008). The author argued that the long history of apparent safe use of phytoestrogenic products may not provide enough confidence that they are indeed without risk. However, in relation to soy isoflavones, even if the claims that there are no toxic effects at dietary levels were correct, this does not provide evidence that soy products are safe. This is because a report by Adams (1995) stated that low levels of estrogenicity may cause temporary infertility in the absence of any visible signs. Alfalfa containing 25 ppm coumestrol decreased the ovulation rate in ewes and cattle fed haylage containing 37 ppm coumestrol showed clinical signs of estrogenic stimulation. It was the toxicity of dietary levels of isoflavones to animals that first raised the awareness of the scientific community to the fact that soy isoflavones were endocrine disruptors. Reproductive effects, infertility, thyroid disease or liver disease due to dietary intake of isoflavones had been observed for several laboratory animals.
Toxicity Effects of Dietary Isoflavones on Reproductive Characteristics of Large Animals
Both red clover silage contains isoflavones which have been reported to cause infertility in cattle. The role of phytoestrogens in causing infertility was recognized by accompanying signs of estrogenism including mammary development, swelling of the vulva discharge of cervical mucus and enlargement of the uterus. Many cow suffered cystic ovaries, with behavioral abnormalities including irregular estrus, nymphomania and anestrous. Cows failed to conceive to service (Adams, 1995; Burton and Wells, 2002).
The condition resolved several weeks or months after removal of the estrogenic feed as ovarian function recovered. Resumption of ovarian activity may be hastened by treatments used for cystic ovarian disease. In combination with estrogenic implants used for growth promotion, phytoestrogens produce additive effects with steers showing sexual behavior and heifers, showing udder development and prolapse of the vagina and rectum.
It has been reported that in the past 50 years, millions of ewes have been rendered infertile by phytoestrogens in Western Australia, a problem that still continues in a subclinical form toady (Adams, 1995). The infertility is caused by estrogenic isoflavone compounds found in certain strains of subterranean clover (Trifolium subterraneum L.) The effects of phytoestrogens on sheep are Clover disease, temporary infertility and permanent infertility.
Toxicity Effects of Dietary Isoflavone on Reproductive Characteristics of Laboratory Animals
Laboratory animals like rats and mice have been used to test the effects of phytoestrogens and enormous reports exist to give an insight on the potentials of phytoestrogens in animal production. It has been now established that a low dose of isoflavone based diet can induce developmental and maturational abnormalities in laboratory animal. In rats Whitten et al. (1994) have shown that a low does (0.01%) coumestrol based diet induces vaginal opening within female at a lower body weights than those with higher body weights. Neonatal exposure to coumestrol in mice can induce significant long term reproductive tract abnormalities, including vaginal cysts, persistent vaginal cornification, endometrial squamous metaplasia, absence of corpora lutea, increased ceroid deposition in the ovaries and the presence of hemorrhagic follicles (Burton and Wells, 2002). Soy isoflavone supplement treatment resulted in a significant decrease in receptive behavior in estrogen and progesterone primed females (Patisaul et al., 2001). The observed disruption of sexual receptivity by the isoflavone supplement is probably due to antiestrogenic effects observed in the brain, suggesting that isoflavone phytoestrogens are antigenic on both ER-alpha and ER-beta dependent gene expression in the brain and estrogen-dependent behavior. It has been reported (Wanibuchi et al., 2003) that carcinogenic potential effect on the uterus of rodents to prenatal exposure of genistein depends on the timing and duration of exposure. Genistein treatment during the perinatal period resulted in lower body weight and lower relative uterine-ovarian weight at 35 days and a prolonged estrus cycle. Treatment of rat dams with a 100 ppm coumestrol diet from birth to postnatal day 21 induced premature anovulation in female offspring and treatment from birth to postnatal day 10 suppressed sexual behaviors in male offspring (Whitten et al., 2002). Maternal exposure to subcutaneous administration of genistein can increase mammary tumorigenesis in the offspring, mimicking the effects of in utero estrogenic exposures (Hilakivi-Clarke et al., 1999). This is feasible since it has been shown that genistein aglycone crosses the rat placenta and can reach fetal brain from maternal serum genistein (Doerge et al., 2001).
CONCLUSION
Clinical data support that soy isoflavones have some potential beneficial and deleterious effects on animals and humans. At the same time, animal experiments and in vitro studies have shown that the biological effects of isoflavone can as well be beneficial especially in the treatment of some cancer and cardiovascular related diseases. These effects may be dependent on the timing and duration of exposure.
Although, phytohormones have been known for their toxic effects on laboratory and large animals in the past, recently phytohormones are receiving attention in the scientific community because of their potential beneficial effects. Medical researchers should concentrate efforts on how to control the toxic effects of the phytohormones and at the same time find ways to improve and maximize the few beneficial effects of the isoflavones.
It is apparent that the study of soy-phytochemicals and their potential effects on human health will become a topic of ever-increasing topic in the future because of their numerous beneficial effects. As our understanding of specific phytochemicals increases, there will undoubtedly be a multitude of applications to modern medicine that will benefit humans.
REFERENCES
- Adams, N.R., 1995. Detection of the effects of phytoestrogens on sheep and cattle. J. Anim. Sci., 73: 1509-1515.
Direct Link - Adams, M.R., G.D.L. Williams, J.K. Franke, A.A. Register and J.R. Kaplan, 2005. Soy protein containing isoflavones reduces the size of atherosclerotic plaques without affecting coronary artery reactivity in adult male monkeys. J. Nutr., 135: 2852-2856.
Direct Link - Adlercreutz, H., 1990. Western diet and west diseases: Some hormonal and biochemical mechanisms and associations. Scandinavian J. Clin. Lab. Invest., 201: 3-23.
PubMed - Adlercreutz, H., 1997. Evolution, nutrition, intestinal microflora and prevention of cancer: A hypothesis. Proc. Soc. Exp. Biol. Med., 217: 241-246.
Direct Link - Adlercreutz, C.H., B.R. Goldin, S.L. Gorbach, K.A. Hockerstedt and S. Watanabe et al., 1995. Soybean phytoestrogen intake and cancer risk. J. Nutr., 125: 757S-770S.
PubMed - Adlercreutz, H., E. Hamalainen, S. Gorbach and B. Goldin, 1992. Dietary phyto-oestrogens and the menopause in Japan. Lancet, 339: 1233-1233.
PubMed - Adlercreutz, H., H. Honjo, A. Higashi, T. Fotsis, E. Hamalainen, T. Hasegawa and H. Okada, 1991. Urinary excretion of lignans and isoflavonoid phytoestrogens in Japanese men and women consuming a traditional Japanese diet. Am. J. Clin. Nutr., 54: 1093-1100.
PubMed - Adlercreutz, H., W. Mazur, P. Bartels, V.V. Elomaa and S. Watanabe et al., 2000. Phytoestrogens and prostate disease. J. Nutr., 130: 658S-659S.
Direct Link - Alekel, D.L., A.S. Germain, C.T. Peterson, K.B. Hanson, J.W. Stewart and T. Toda, 2000. Isoflavone-rich soy protein isolate attenuates bone loss in the lumbar spine of perimenopausal women. Am. J. Clin. Nutr., 72: 844-852.
PubMed - Anderson, J.W., B.M. Johnstone and M.E. Cook-Newell, 1995. Meta-analysis of the effects of soy protein intake on serum lipids. N. Engl. J. Med., 333: 276-282.
CrossRefPubMedDirect Link - Arjmandi, B.H., R. Birnbaum, N.V. Goyal, M.J. Getlinger and S. Juma et al., 1998. Bone-sparing effect of soy protein in ovarian hormone-deficient rats is related to its isoflavone content. Am. J. Clin. Nutr., 68: 1364-1368.
Direct Link - Alekel, L., C.M. Hasler, S. Juma, B.W. Drum and S.C. Kukreja, 1998. Role of soy protein with normal or reduced isoflavone content in reversing bone loss induced by ovarian hormone deficiency in rats. Am. J. Clin. Nutr., 68: 1358S-1363S.
PubMed - Axelson, M., D.N. Kirk, R.D. Farrant, G. Cooley, A.M. Lawson and K.D. Setchell, 1982. The identification of the weak oestrogen equol [7-hydroxy-3-(4`-hydroxyphenyl)chroman] in human urine. Biochem. J., 201: 353-357.
PubMed - Axelson, M., J. Sjovall, B.E. Gustafsson and K.D. Setchell, 1984. Soy-a dietary source of the non-steroidal oestrogen equol in man and animals. J. Endocrinol., 102: 49-56.
PubMed - Barnes, S., M. Kirk and L. Coward, 1994. Isoflavones and their conjugates in soy foods: Extraction conditions and analysis by HPLC-Mass spectrometry. J. Agric. Food Chem., 42: 2466-2474.
CrossRef - Bojase, G., R.R. Majinda, B.A. Gashe and C.C. Wanjala, 2002. Antimicrobial flavonoids from Bolusanthus speciosus. Planta Med., 68: 615-620.
PubMed - Boue, S.M., C.H. Carter, K.C. Ehrlich and T.E. Cleveland, 2000. Induction of soybean phytoalexins coumestrol and glyceollin by Aspergillus. J. Agric. Food Chem., 48: 2167-2172.
CrossRefDirect Link - Burton, J.L. and M. Wells, 2002. The effect of phytoestrogens on the female genital tract. J. Clin. Pathol., 55: 401-407.
PubMed - Bylund, A., J.X. Zhang, A. Bergh, J.E. Damber and A. Widmark et al., 2000. Rye bran and soy protein delay growth and increase apoptosis of human LNCaP prostate adenocarcinoma in nude mice. Prostate, 42: 304-314.
PubMed - Carroll, K.K. and E. Kurowska, 1995. Soy consumption and cholesterol reduction: Review of animal and human studies. J. Nutr., 125: 594S-597S.
PubMed - Cassidy, A., S. Bingham and K.D. Setchell, 1994. Biological effects of a diet of soy protein rich in isoflavones on the menstrual cycle of premenopausal women. Am. J. Clin. Nutr., 60: 333-340.
Direct Link - Cassidy, A., S. Bingham and K. Setchell, 1995. Biological effects of isoflavones in young women: Importance of the chemical composition of soybean products. Br. J. Nutr., 74: 587-601.
PubMed - Choi, Y.H., W.H. Lee, K.Y. Park and L. Zhang, 2000. p53-independent induction of p21 (WAF1/CIP1), reduction of cyclin B1 and G2/M arrest by the isoflavone genistein in human prostate carcinoma cells. Jap. J. Cancer Res., 91: 164-173.
PubMed - Clarkson, T.B., M.S. Anthony, J.K. Williams, E.K. Honore and J.M. Cline, 1998. The potential of soybean phytoestrogens for postmenopausal hormone replacement therapy. Proc. Soc. Exp. Biol. Med., 217: 365-368.
PubMed - Cline, J.M., J.C. Paschold, M.S. Anthony, I.O. Obasanjo and M.R. Adams, 1996. Effects of hormonal therapies and dietary soy phytoestrogens on vaginal cytology in surgically postmenopausal macaques. Fertil. Steril., 65: 1031-1035.
PubMed - Coward, L., N.C. Barnes, K.D.R. Setchell and S. Barnes, 1993. Genistein and daidzein and their β-glucoside conjugates: Anti-tumor isoflavones in soybean foods from American and Asian diets. J. Agric. Food Chem., 41: 1961-1967.
Direct Link - Day, A.J., M.S. DuPont, S. Ridley, M. Rhodes, M.J. Rhodes, M.R. Morgan and G. Williamson, 1998. Deglycosylation of flavonoid and isoflavonoid glycosides by human small intestine and liver β-glucosidase activity. FEBS Lett., 436: 71-75.
PubMed - Divi, R.L., H.C. Chang and D.R. Doerge, 1997. Anti-thyroid isoflavones from soybean. Biochem. Pharmacol., 54: 1087-1096.
CrossRef - Franke, A.A. and L.J. Custer, 1994. High-performance liquid chromatographic assay of isoflavenoids and coumestrol for human urine. J. Chromatgr. B Biomed. Appl., 662: 47-60.
PubMed - Gyorgy, P., K. Murata and H. Ikehata, 1964. Antioxidant isolated from fermented soybeans (Tempeh). Nature, 203: 870-872.
CrossRefDirect Link - Heim, K.E., A.R. Tagliaferro and D.J. Bobilya, 2002. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem., 13: 572-584.
CrossRefDirect Link - Hilakivi-Clarke, L., E. Cho, I. Onojafe, M. Raygada and R. Clarke, 1999. Maternal exposure to genistein during pregnancy increases carcinogen-induced mammary tumorigenesis in female rat offspring. Oncol. Rep., 6: 1089-1095.
PubMed - Honore, E.K., J.K. Williams, M.S. Anthony and T.B. Clarkson, 1997. Soy isoflavones enhanced coronary vascular reactivity in atherosclerotic female macaques. Fertil. Steril., 67: 148-154.
PubMed - Huff, M.W., R.M.G. Hamilton and K.K. Carroll, 1977. Plasma cholesterol levels in rabbits fed low fat, cholesterol-free, semi-purified diets: Effects of dietary proteins, protein hydrolysates and amino acid mixtures. Atherosclerosis, 28: 187-195.
Direct Link - Huff, M.W., D.C. Roberts and K.K. Carroll, 1982. Long-term effects of semipurified diets containing casein or soy protein isolate on atherosclerosis and plasma lipoproteins in rabbits. Atherosclerosis, 41: 327-336.
PubMed - Huff, M.W. and K.K. Carroll, 1980. Effects of dietary protein on turnover, oxidation and absorption of cholesterol and on steroid excretion in rabbits. J. Lipid Res., 21: 546-548.
Direct Link - Humfrey, C.D., 1998. Phytoestrogens and human health effects: Weighing up the current evidence. Nat. Toxins, 6: 51-59.
PubMed - Joannou, G.E., G.E. Kelly, A.Y. Reeder, M. Waring and C. Nelson, 1995. A urinary profile study of dietary phytoestrogens. The identification and mode of metabolism of new isoflavonoids. J. Steroid Biochem. Mol. Biol., 54: 167-184.
PubMed - Kagan, A., B.R. Harris, W. Winkelstein Jr., K.G. Johnson and H. Kato et al., 1974. Epidemiologic studies of coronary heart disease and stroke in Japanese men living in Japan, Hawaii and California: Demographic, physical, dietary and biochemical characteristics. J. Chronic. Dis., 27: 345-364.
PubMed - Kapiotis, S., M. Hermann, I. Held, C. Seelos, H. Ehringer and B.M. Gmeiner, 1997. Genistein, the dietary-derived angiogenesis inhibitor, prevents LDL oxidation and protects endothelial cells from damage by atherogenic LDL. Arteriosclerosis Thrombosis Vascular Biol., 17: 2868-2874.
PubMed - Setchell, K.D.R., N.M. Brown, L. Zimmer-Nechemias, W.T. Brashear, B.E. Wolfe, A.S. Kirschner and J.E. Heubi, 2002. Evidence for lack of absorption of soy isoflavone glycosides in humans, supporting the crucial role of intestinal metabolism for bioavailability. Am. J. Clin. Nutr., 76: 447-453.
Direct Link - Kitaoka, M., H. Kadokawa, M. Sugano, K. Ichikawa and M. Taki et al., 1998. Prenylflavonoids: A new class of non-steroidal phytoestrogen (Part 1). Isolation of 8-isopentenylnaringenin and an initial study on its structure-activity relationship. Planta Medica, 64: 511-515.
PubMed - Khosla, P., S. Samman, K.K. Carroll and M.W. Huff, 1989. Turnover of 125IVLDL and 131I-LDL apolipoprotein B in rabbits fed diets containing casein or soy protein. Biochem. Biophys. Acta, 1002: 157-163.
PubMed - Kolonel, L., 1988. Variability in diet and its relation to risk in ethnic and migrant groups. Basic Life Sci., 43: 129-135.
PubMed - Kuiper, G.G., B. Carlsson, K. Grandien, E. Enmark, J. Haggblad, S. Nilsson and J.A. Gustafsson, 1997. Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and beta. Endocrinology, 138: 863-870.
PubMed - Lamartiniere, C.A., J. Moore, M. Holland and S. Barnes, 1995. Neonatal genistein chemoprevents mammary cancer. Proc. Soc. Exp. Biol. Med., 208: 120-123.
PubMed - Lee, H.P., L. Gourley, S.W. Duffy, J. Esteve, J. Lee and N.E. Day, 1991. Dietary effects on breast-cancer risk in Singapore. Lancet, 337: 1197-1200.
PubMed - Leopold, A.S., M. Erwin, J. Oh and B. Browning, 1976. Phytoestrogens: Adverse effects on reproduction in California quail. Science, 191: 98-100.
CrossRefPubMedDirect Link - Liggins, J., R. Grimwood and S.A. Bingham, 2000. Extraction and quantification of lignan phytoestrogens in food and human samples. Anal. Biochem., 278: 102-109.
CrossRef - Lu, L.J., K.E. Anderson, J.J. Grady, F. Kohen and M. Nagamani, 2000. Decreased ovarian hormones during a soya diet: Implications for breast cancer prevention. Cancer Res., 60: 4112-4121.
Direct Link - Lu, L.J., K.E. Anderson, J.J. Grady and M. Nagamani, 1996. Effects of soy consumption for one month on steroid hormones in premenopausal women: Implications of breast cancer risk reduction. Cancer Epidemiol. Biomarkers Prev., 5: 63-70.
Direct Link - Phipps, W.R., M.C. Martini, J.W. Lampe, J.L. Slavin and M.S. Kurzer, 1993. Effect of flax seed ingestion on menstrual cycle. J. Clin. Endocrinol. Metab., 77: 1215-1219.
Direct Link - Maskarinec, G., S. Singh, L. Meng and A.A. Franke, 2001. Dietary soy intake and urinary isoflavone excretion among women from multiethonic population. Cancer Epidemiol. Biomarkers Prev., 7: 613-619.
PubMed - McMahon, L.G., H. Nakano, M.D. Levy and J.F. Gregory, 1997. Cytosolic pyridoxine-β-D-glucoside hydrolase from porcine jejunal mucosa. Purification, properties and comparison with broad specificity β-glucosidase. J. Biol. Chem., 272: 32025-32033.
PubMed - Middleton Jr. E., C. Kandaswami and T.C. Theoharides, 2000. The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacol. Rev., 52: 673-751.
PubMedDirect Link - Miksicek, R.J., 1994. Interaction of naturally occurring mousterodial estrogens with expressed recombinant lumen estrogen receptor. J. Steroid Biochem. Mol. Biol., 49: 153-160.
PubMed - Miksicek, R.J., 1995. Estrogenic flavonoids: Structural requirements for biological activity. Proc. Soc. Exp. Biol. Med., 208: 44-50.
Direct Link - Milligan, S.R., J.C. Kalita, A. Heyerick, H. Rong, L. de Cooman and D. de Keukeleire, 1999. Identification of a potent phytoestrogen in hops (Humulus lupulus L.) and beer. J. Clin. Endocrinol. Metab., 83: 2249-2252.
PubMed - Miyamoto, M., Y. Matsushita, A. Kiyokawa, C. Fukuda, Y. Iijima, M. Sugano and T. Akiyama, 1998. Prenylflavonoids: A new class of non-steroidal phytoestrogens (Part 2). Estrogenic effects of 8-isopentenylnaringenin on bone metabolism. Planta Medica, 64: 516-519.
PubMed - Morrison, J.J., S.R. Dearn, S.K. Smith and A. Ahmed, 1996. Activation of protein kinase C is required for oxytocin-induced contractility in human myometrium. Hum. Reprod., 11: 2285-2290.
PubMed - Mosselman, S., J. Polman and R. Dijkema, 1996. ERβ: Identification and characterization of a novel human estrogen receptor. FEBS Lett., 392: 49-53.
CrossRef - Murkies, A.L., C. Lombard, B.J. Strauss, G. Wilcox, H.G. Burger and M.S. Morton, 1995. Dietary flour supplementation decreases post menopausal hot flushes: Effect of soy and wheat. Maturitas, 21: 189-195.
PubMed - Murphy, P.A., 1982. Phytoestrogen content of processed soybean products. Food Tech., 36: 60-64.
Direct Link - Omede, A.A., M.N. Opara and I.C. Okoli, 2008. The significance of phytohormones in animal production. Int. J. Trop. Agric. Food Syst., 2: 89-104.
Direct Link - Palmier, B., D. Leiber and S. Harbon, 1996. Pervanadate mediated an increased generation of inositol phosphates and tension in rat myometrium. Activation and phosphorylation of phospholipase C-gamma 1. Biol. Repord., 54: 1383-1389.
PubMed - Picherit, C., V. Coxam, C. Bennetau-Pelissero, S. Kati-Coulibaly, M.J. Davicco, P. Lebecque and J.P. Barlet, 2000. Daidzein is more efficient than genistein in preventing ovariectomy-induced bone loss in rats. J. Nutr., 130: 1675-1681.
PubMedDirect Link - Ranich, T., S.J. Bhathena and M.T. Velasquez, 2001. Protective effects of dietary phytoestrogens in chronic renal disease. J. Ren. Nutr., 11: 183-193.
PubMed - Raschke, M., I.R. Rowland, P.J. Magee and B.L. Pool-Zobel, 2006. Genistein protects prostate cells against hydrogen peroxide-induced DNA damage and induces expression of genes involved in the defence against oxidative stress. Carcinogenesis, 27: 2322-2330.
PubMed - Robinson, D., T. Kawamura, S. Hinohara, Y. Sakamoto and T. Takahashi, 1995. Levels of cardiovascular risk factors in Japanese people living in the UK. J. Cardiovasc. Risk, 2: 449-458.
PubMed - Rose, D.P., A.P. Boyar and E.L. Wynder, 1986. International comparison of mortality rates for cancer of the breast, ovary, prostate and colon and per capita food consumption. Cancer, 58: 2363-2371.
PubMed - Samman, S., P. Khosia1 and K.K. Carroll, 1989. Effects of dietary casein and soy protein on metabolism of radiolabelled low density apolipoprotein B in rabbits. Lipids, 24: 169-172.
CrossRefDirect Link - Scarlata, S. and R. Miksicek, 1995. Binding properties of coumestrol to expressed human estrogen receptor. Mol. Cell. Endocrinol., 115: 65-72.
CrossRef - Setchell, K.D., 1998. Phytoestrogens: The biochemistry, physiology and implications for human health of soy isoflavones. Am. J. Clin. Nutr., 68: 1333S-1346S.
Direct Link - Setchell, K.D. and A. Cassidy, 1999. Dietary isoflavones: Biological effects and relevance to human health. J. Nutr., 129: 758s-767s.
PubMedDirect Link - Setchell, K.D. and S.J. Cole, 2003. Variations in isoflavone levels in soy foods and soy protein isolates and issues related to isoflavone databases and food labeling. J. Agric. Food Chem., 51: 4146-4155.
PubMed - Shirataki, Y., S. Matsuoka, M. Komatsu, M. Ohyama, T. Tanaka and M. Iinuma, 1999. Four isoflavanones from roots of Sophora tetraptera. Phytochemistry, 50: 695-701.
CrossRef - Shutt, D.A. and R.I. Cox, 1972. Steroid and phytoestrogen binding to sheep uterine receptors in vitro. J. Endocrinol., 52: 299-310.
Direct Link - Sirtori, C.R. and M.R. Lovati, 2001. Soy proteins and cardiovascular disease. Curr. Atherosclerosis Rep., 3: 47-53.
CrossRefDirect Link - Strauss, L., S. Makela, S. Joshi, I. Huhlanierri and R. Santti, 1998. Genistein exerts estrogen-like effects in male mouse reproductive tract. Mol. Cell. Endocrinol., 144: 83-93.
Direct Link - Strom, S.S., Y. Yamamura, C.M. Duphorne, M.R. Spitz, R.J. Babaian, P.C. Pillow and S.D. Hursting, 1999. Phytoestrogen intake and prostate cancer: A case-control study using a new database. Nutr. Cancer, 33: 20-25.
CrossRefPubMedDirect Link - Verger, P.H. and J.C. Leblane, 2003. Concentration of phytohormone in food and feed and their impact on the human exposure. J. Pure Applied Chem., 75: 1873-1880.
Direct Link - Wanibuchi, H., J.S. Kang, E.I. Salim, K. Morimura and S. Fukushima, 2003. Toxicity versus beneficial effects of phytoestrogens. Pure Applied Chem., 75: 2047-2053.
Direct Link - Whitten, P.L., E. Russell and F. Naftolin, 1994. Influence of phytoestrogen diets on estradiol action in the rat uterus. Steroids, 59: 443-449.
PubMed - Whitten, P.L., H.B. Patisaul and L.J. Young, 2002. Neurobehavioral actions of coumestrol and related isoflavonoids in rodents. Neurotoxicol. Teratol., 24: 47-54.
CrossRef - Wright, J.S., E.R. Johnson and G.A. DiLabio, 2001. Predicting the activity of phenolic antioxidants: Theoretical method, analysis of substituent effects and application to major families of antioxidants. J. Am. Chem. Soc., 123: 1173-1183.
CrossRefDirect Link - Wu, A.H., R.G. Ziegler, P.L. Horn-Ross, A.M. Nomura and D.W. West et al., 1996. Tofu and risk of breast cancer in Asian-Americans. Cancer Epidemiol. Biomarkers Prev., 5: 901-906.
PubMed - Xu, X., K.S. Harris, H.J. Wang, P.A. Murphy and S. Hendrich, 1995. Bioavailability of soybean isoflavones depends upon gut microflora in women. J. Nutr., 125: 2307-2315.
PubMed - Yamakoshi, J., M.K. Piskula, T. Izumi, K. Tobe and M. Sait et al., 2000. Isoflavone aglycone-rich extract without soy protein attenuates atherosclerosis development in cholesterol-fed rabbits. J. Nutr., 130: 1887-1893.
Direct Link