
Xing Fan
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang 611130, Sichuan, China
Zi-Jian Song
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang 611130, Sichuan, China
Hou-Yang Kang
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang 611130, Sichuan, China
Rui-Wu Yang
College of Biology and Science, Sichuan Agricultural University, Yaan 625014, Sichuan, China
Yong-Hong Zhou
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang 611130, Sichuan, China
International Journal of Agricultural Research
Year: 2009 | Volume: 4 | Issue: 8 | Page No.: 237-249







ABSTRACT
To evaluate High Molecular Weight (HMW) glutenin subunit comparison in Triticum turgidum sp. polonicum, HMW glutenin subunits of 22 accessions were analyzed using SDS-PAGE. The result showed that the variation in the number of HMW glutenin subunits in this taxon ranged from 1-3. A dwarfing accession of T. turgidum sp. polonicum (dwarfing polish wheat) from Xingjiang in China has two putative expressed HMW glutenin subunits. Three complete coding region sequences of HMW glutenin subunit genes were amplified from dwarfing polish wheat using PCR method. The PCR products were derived from 1Ax (1Ax-dp), 1Bx gene (1Bx-dp) and a silent 1Ay (1Ay-dp) gene. The complete ORF of x-type HMW glutenin genes, 1Ax-dp and 1Bx-dp, were 2508 and 2385 bp in size, encoding 836 and 795 amino acids, respectively. The silent 1Ay-dp gene contained an in-frame internal stop codon and was 1806 bp in size. Sequence comparison indicated that they had primary structure identical to the published HMW glutenin subunits. Phylogenetic analysis suggested that 1Ax-dp from dwarfing polish wheat was most closely related to 1Ax1 and 1Ax2* from Triticum aestivum, while 1Bx-dp from dwarfing polish wheat was most closely related to 1Bx7 from Triticum aestivum. Based on sequence comparison, SDS-PAGE and phylogenetic analysis, it can be deduced that 1Ax-dp and 1Bx-dp are additional alleles at the Glu-A1 and Glu-B1 loci, respectively.
PDF Abstract XML References Citation
How to cite this article
Xing Fan, Zi-Jian Song, Hou-Yang Kang, Rui-Wu Yang and Yong-Hong Zhou, 2009. Identification and Characterization of HMW Glutenin Subunits and their Coding
Sequences in Dwarfing Polish Wheat. International Journal of Agricultural Research, 4: 237-249.
DOI: 10.3923/ijar.2009.237.249
URL: https://scialert.net/abstract/?doi=ijar.2009.237.249
DOI: 10.3923/ijar.2009.237.249
URL: https://scialert.net/abstract/?doi=ijar.2009.237.249
INTRODUCTION
The High Molecular Weight (HMW) glutenin subunits are important components of seed storage proteins and account for 10% of the total seed proteins in wheat. The HMW glutenin subunits play particularly important role in determining wheat gluten and dough elasticity (Payne, 1987; Shewry et al., 2002), making the isolation and characterization of the HMW glutenin gene to draw increasing attention (Yan et al., 2006). The HMW glutenin subunits are encoded by Glu-1 loci on chromosomes 1A, 1B and 1D in bread wheat, each locus encoding one x-type and one y-type subunits (Payne, 1987). Due to gene silencing and allelic variation, the composition of HMW glutenin subunits usually differs among hexaploid wheat varieties (Payne and Lawrence, 1983; Wan et al., 2002). The genes coding for subunits 1Bx, 1Dx and 1Dy are always expressed, while genes coding for subunits 1Ax and 1By are expressed in some cultivars (Payne, 1987). 1Ay subunits are occasionally present in hexaploid wheat (Johansson and Svensson, 1995) and more widely in A-genome diploids (Waines and Payne, 1987; Wan et al., 2002; Jiang et al., 2009). Many genes of HMW glutenin subunits have been isolated from bread wheat and the related wild species and the coding region sequences are highly homologous (Wan et al., 2002; Yan et al., 2002, 2006, 2009; Liu et al., 2007). The molecular comparison revealed that the primary structure of different HMW glutenin subunits shares a common model, which consists of three domains: (1) a small N-terminal domain ranging from 80 to 120 amino acids with 3 to 5 cysteine residues; (2) a large repetitive domain with many repeated short peptide motifs (GQQ; PGQGQQ; GYYPTSP/LQQ or GY/HYPT/ASLQQ); (3) the C-terminus, a non-repetitive domain of ~ 50 residues with one cysteine (Shewry and Tatham, 1997). A signal peptide is adjacent to the N-terminal region. The sizes of signal peptide, the N-terminal and the C-terminal region are well conserved among different HMW subunits and the number of short repeated peptide motif results in variation of length among the repetitive domains of different HMW subunits (Shewry et al., 2002).
Triticum turgidum sp. polonicum L. (2n = 28, AABB) is a most important taxon with agronomic traits of large grains, more spikes and higher tillering ability in Emmer wheat of Triticum. It is one of the best genetic resources for wheat breeding program. Phytogeographically, they are mainly distributed in the area of Mediterranean, Ethiopia and the northwestern part of China. In the present study, HMW glutenin diversity among 22 accessions of T. turgidum sp. polonicum was investigated by using one dimensional SDS-PAGE analysis. As part of ongoing study of molecular identification of the HMW glutenin genes from T. turgidum sp. polonicum, this investigation was made to explore the complete Open Reading Frames (ORFs) of the HMW glutenin gene sequences from dwarfing polish wheat. Dwarfing polish wheat, a dwarfing accession of T. turgidum sp. polonicum, was collected from Tulufan, Xingjiang, China by Prof. Chi Yen and Jun-Liang Yang. It is the only one polish wheat that possesses characteristic of dwarfing in China (Liu et al., 2002).
MATERIALS AND METHODS
Plant Materials
Twenty-two accessions of T. turgidum sp. polonicum were used in this study, including seven from Xinjiang of China, two from the Middle East (Iran and Iraq), four from Africa (Egypt and Ethiopia), five from Europe (Portugal, Germany, Hungary, Italy and Ukraine), one from India, one from the United States and one from Ecuador. The sources of the accessions of T. turgidum sp. polonicum used in this study are listed in Table 1. The accessions with PI and CItr numbers were kindly provided by American National Plant Germplasm System (Pullmam, Washington, USA). The other accessions were provided by Triticeae Research Institute, Sichuan Agricultural University, Sichuan, China. Chinese Spring, LongFu1 and ChuanYu12 are common wheat varieties, which were collected locally from Sichuan Province in China. This study was conducted in July 2004 at Dujiangyan City, Triticeae Research Institute of Sichuan Agricultural University.
SDS-PAGE Analysis
Seed proteins were prepared for SDS-PAGE analysis as described previously (Yan et al., 2002). HMW glutenin subunits from hexaploid wheat varieties LongFu1 (1Ax2*, 1Bx7+1By8, 1Dx5+1Dy10), Chinese Spring (null, 1Bx7+1By8, 1Dx2+1Dy12) and ChuanYu12 (1Ax, 1Bx7+1By8, 1Dx5+1Dy10) were used as reference for accessing the electrophoretic mobilities of the HMW glutenin subunits of T. turgidum sp. polonicum in SDS-PAGE analysis (Yan et al., 2006).
| Table 1: | Plant materials used in this study |
 | |
PCR Amplification
Total genomic DNA was isolated from fresh leaf using a modified CTAB extraction method (Doyle and Doyle, 1987). The complete ORFs of the HMW glutenin subunit genes were amplified by PCR using the following oligonucleotides as primers: P1: 5’-ATGGCTAAGCGGC/TTA/GGTCCTCTTTG-3’, P2: 5’-CTATCACTGGCAT/GGCCGACAATG CG-3’. PCR reactions were performed in a reaction volume of 50 μL according to the methods of Yan et al. (2002). The PCR products were separated by 1.0% agarose gels.
Cloning and Sequence Analysis
Amplified DNA fragments were purified with EZNATM gel extraction kit (Omega). Cleaned PCR products were cloned into pMD18-T vector (TaKaRa Biotechnology (Dalian) Co., Ltd, Liaoning, China) and the products of ligation were transformed into DH5α competent cells. Positive transformants were used directly as templates in a PCR with standard primers M13R and M13F (TaKaRa), which attach on either side of the cloning site in the plasmid. Three independent clones were selected for analysis of the nested deletion subclones. A set of nested deletion subclones of the positive clone, including pAx-dp, pBx-dp and pAy-dp, were conducted and sequenced according to the procedure of Yan et al. (2002). The construction of full length sequence was assembled from sequence data generated from seven to eight subclones. The amino acid sequences of HMW glutenin gene (1Ax-dp, 1Bx-dp and 1Ay-dp) were deduced from the DNA sequence using ORF Finder program in the NCBI. Comparisons of the deduced amino acid sequence with those of previous published typical HMW glutenin\ subunits were performed with program BLAST and PSI-BLAST in the NCBI non-redundant database (Altschul et al., 1990; Park et al., 1998). Phylogenetic trees were constructed with p-distance methods using the software MEGA 3 (Kumar, S., K. Tamura, I. Jakobsen and M. Nei, www.megasoftware.net). Nodal supports in phylogenetic trees were estimated using 1000 fast bootstrap replicates (Felsenstein, 1985).
RESULTS
Hmw Glutenin Subunits of T. turgidum sp. polonicum
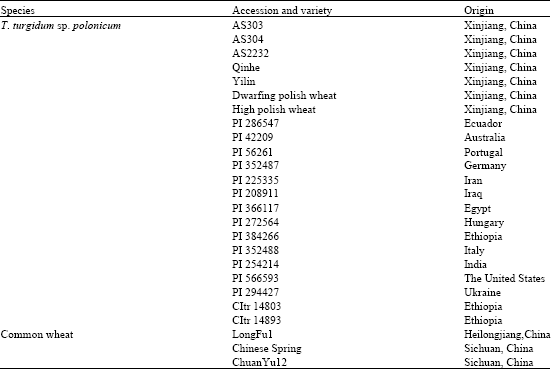
In SDS-PAGE analysis of seed protein extracts, three main types of HMW glutenin subunits from T. turgidum sp. polonicum were found based on the number of protein band (Fig. 1). The electrophoretic mobilities of these protein bands were compared to those of hexaploid wheat HMW glutenin subunits. The first type of HMW glutenin subunits found in accession PI56261 (Bx7), PI225335 (By8), PI208911 (Bx7), PI36117 (Bx7), PI384266 (By8), PI254214 (By8), PI566593 (By8), PI29447 (Bx7), CItr14892 (By8), Yilin (Bx7) and high polish wheat (Bx7) includes only one protein band. The electrophoretic mobility of these subunits was similar to that of the 1Bx7 or 1By8, respectively. The second type of HMW glutenin subunits found in accession AS303 (Bx7+?) (? represents unknown subunits), AS304 (By8+?), Qinhe (Bx7+By8), dwarfing polish wheat (Ax-dp+Bx7) (designed as Ax-dp+Bx-dp), PI42209 (Bx7+?), PI352487 (?+Bx7) and PI272564 (?+By8) contains two protein bands. The mobility of HMW glutenin subunits of accession AS303, AS304, Qinhe, PI4229 and PI272564 was near that of 1Bx7 and 1By8. For dwarfing polish wheat, the mobility of one subunit was similar to that of 1Bx7, while the mobility of the other subunit was situated that of between 1Ax2* and 1Dx5 subunit. The mobility of HMW glutenin subunits of accession PI352487 was different from that of dwarfing polish wheat, one of which was similar to that of 1Bx7, while the other similar to that of 1Ax1. The third type of subunits found in accession AS2232 (2*+8+?), PI352488 (?1+Bx7+?2), PI286547 (2*+?1+?2) and CItr14803 (?+Bx7+By8) comprises three bands. The mobility of subunit among accession is different from each other.
Cloning and Sequencing of HMW Glutenin Gene ORFs
Two DNA fragments were amplified using the primers P1 and P2 in genomic PCR experiments. After ligating the two fragments into the plasmid vector pMD18-T, three types of clones with insert sizes including approximately 1.8 and 2.5 kb were obtained. The inserts within three plasmid clones were p1Ax-dp, p1Bx-dp and p1Ay-dp. These plasmid clones were represented the 1Ax-dp, 1Bx-dp and 1Ay-dp gene by restriction enzyme digestion analysis and two terminal sequencing, respectively. The ORFs of 1Ax-dp, 1Bx-dp and 1Ay-dp were sequenced completely using subclones prepared by nested deletion.
Amino Acid Sequence Comparisons
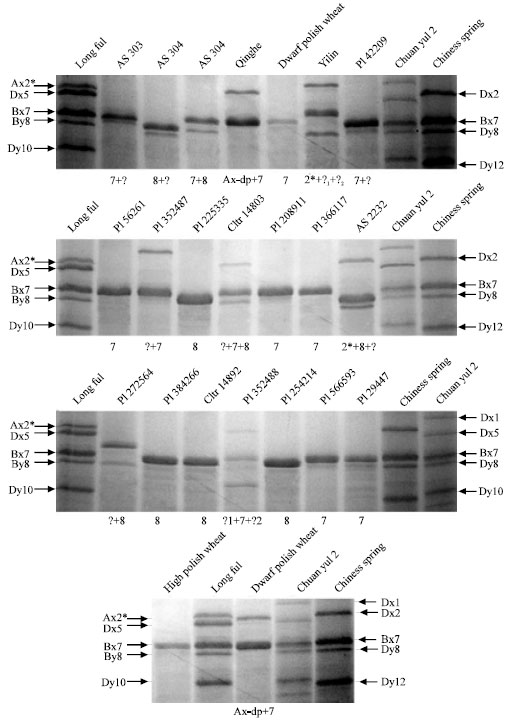
All the amplified products were completely sequenced using subclones. Sequence combination showed that the complete ORFs of HMW glutenin genes, 1Ax-dp and 1Bx-dp, were 2508 and 2385 bp in size, encoding 836 and 795 amino acids, respectively (Fig. 2 A, B and Fig. 3). Amino acid sequences derived from 1Ay-dp sequences suggest that it may be a silent gene resulting from an in-frame premature stop codon (Fig. 2C). The putative gene 1Ay-dp has 1806 bp, encoding 602 amino acids including an in-frame premature stop codon.
Analysis of amino acid sequences derived from the cloned ORFs showed that both HMW glutenin subunits 1Ax-dp and 1Bx-dp include a signal peptide, a non-repetitive N-and C-terminal domain and a central repetitive domain. These two subunits share a similar repeat structure to other known x-type subunits, which contain three types of repeat units (tripeptides, hexapeptides and nonapeptides).
 | |
| Fig. 1: | SDS-PAGE analysis of HMW glutenin subunits of 22 accessions of T. turgidum sp. polonicum LongFu1 (1Ax2*, 1Bx7 + 1By8, 1Dx5 + 1Dy10), Chinese Spring (null, 1Bx7 + 1By8, 1Dx2 + 1Dy12) and ChuanYu12 (1Ax1, 1Bx7 + 1By8, 1Dx5 + 1Dy10) were used as reference markers |
 | |
 | |
| Fig. 2: | Nucleotide and deduced amino acid sequences of HMW glutenin subunit genes, (A) 1Ax-dp, (B) 1Bx-dp and (C) 1Ay-dp in dwarfing polish wheat. The putative signal peptide, N-terminal and C-terminal are marked. represent for the tripeptide, hexapeptide and nonapeptide repeat units, respectively. Two stop codons are indicated with asterisks. An asterisk with box in 1Ay-dp gene sequence indicates in-frame premature stop codon |
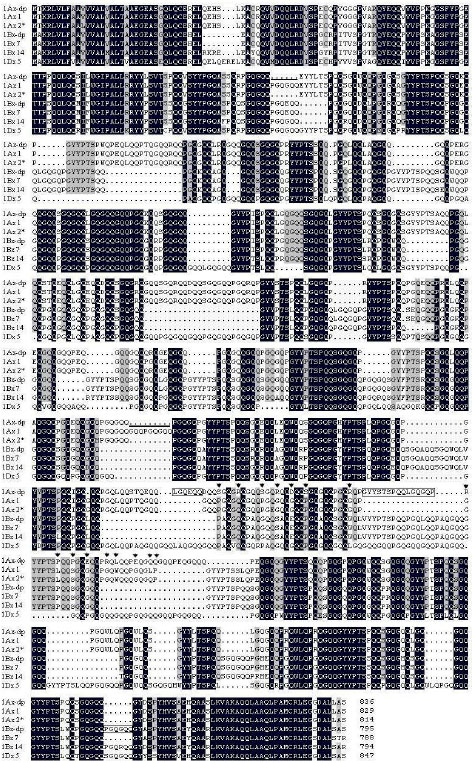 | |
| Fig. 3: | Comparison of the primary structure of 1Ax-dp and 1Bx-dp with those of known x-type HMW glutenin subunits. The dots with underline indicate the position of the amino acid deletions, while arrows show the position of the amino acid substitution compared 1Ax-dp with 1Ax. The amino acid residues with box indicate the position of the amino acid insertions |
Four cysteine residues with three in the N-terminal region and one in the C-terminal region are conservative in 1Ax-dp and 1Bx-dp.
These characteristic structures were identical to the one shared by published x-type HMW glutenin subunits (Fig. 2A, B). In comparison to those of the representatives of published HMW glutenin subunits, some properties of the 1Ax-dp and 1Bx-dp subunits were summarized in Table 2.
A high degree of homology was observed between the 1Ax-dp and 1Ax. 1Ax-dp has 6 tripeptides, 76 hexapeptides and 21 nonapeptides. These repeated units are regularly arranged in tandem and interspersed arrays. In the central repetitive domain of 1Ax-dp, two insertions of hexapeptide (LGQEQQ and LGQGQP), one insertion of nonapeptide (GYYSTSPQQ), two deletions of hexapeptide (PGQGQQ and PGQGQQ) and one deletion of tripeptide (GQQ) were observed compared with the published 1Ax1 gene. There are sixteen amino acid substitutions in the central repetitive region compared 1Ax-dp to 1Ax (Fig. 3). In addition to the single amino acid substitutions, 1Ax-dp has three insertions of hexapeptide (LGQEQQ, LGQGQP and GQQPEQ), one insertion of tetrapeptide (GQQQ), one insertion of nonapeptide (GYYSTSPQQ), one deletion of nonapeptide (GYYPTSSLQ) compared to the published 1Ax2*.
1Bx-dp has 5 tripeptides, 68 hexapeptides and 23 nonapeptides. In the central repetitive domain of 1Bx-dp, there was one insertion of hexapeptide (PGQGQQ) compared with the published 1Bx7 subunits. 1Bx-dp and 1Bx7 differed in two amino acid residues at positions 794 and 795 with alanine/serine substitution.
Phylogenetic Analysis
Phylogenetic trees were created to understand the relationships between 1Ax-dp and 1Bx-dp and other known HMW glutenin subunits. The amino acid sequences, including 1Ax-dp, 1Bx-dp, 1Ax (AJ 306976), 1Ax2* (M 22208), 1Bx7 (X 13927), 1Bx14 (AY 367771), 1Bx20 (CAD24586), 1Bx23 (AY 553933), 1Dx5 (S 02262), 1Ay (AY 245578), 1By8 (AAO 64642), 1By9 (X 61026), 1By22* (AAP 76393) and 1Dy10 (P 10387), were used to produce a dendrogram (Fig. 4). D-hordein of Hordeum vulgare (AAP 31051) was used as outgroup. In the phylogenetic tree, it is obvious that the HMW glutenin subunits were separated into two groups, one group was x-type subunits and the other group was y-type subunits. 1Bx-dp was grouped with 1Bx7 with 99% bootstrap support and clustered with 1Bx23, 1Bx14 and 1Bx20. 1Ax-dp was placed outside 1Ax and 1Ax2* with 100% bootstrap support.
| Table 2: | A summary of some of the properties of the primary structure of the 1Ax-dp and 1Bx-dp subunits in comparison with those of the representatives of published HMW glutenin subunits |
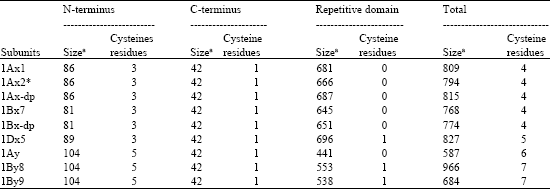 | |
| aThe numbers indicate the number of amino acid residue | |
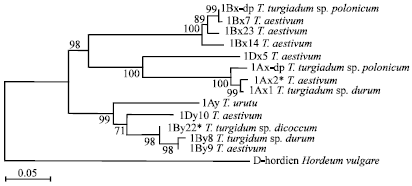 | |
| Fig. 4: | A phylogenetic tree derived from amino acid sequences of HMW glutenin subunits from different Triticum species |
DISCUSSION
The HMW glutenin subunits are encoded by multi-alleles at Glu-1 loci of each of the three genomes (A, B and D) of bread wheat (Payne, 1987). In theory, hexaploid wheat and tetraploid wheat could contain 6 and 4 different HMW glutenin subunits, respectively. Due to the silencing of some of genes at the Glu-A1 and Glu-B1 loci, most bread wheat possess 3 to 5 HMW glutenin subunits and durum wheat has 1 to 3 HMW glutenin subunits (Payne, 1987; Jiang et al., 2009). In this study, SDS-PAGE analysis showed that the variation in the number of HMW subunits in T. turgidum sp. polonicum varieties ranged from one to three, indicating a polymorphism for the HMW glutenin subunits among T. turgidum sp. polonicum varieties. This result did not contradict the suggestion that durum wheat has 1 to 3 HMW glutenin subunits. For the polymorphism of the HMW glutenin subunits among T. turgidum sp. polonicum varieties, we prefer the explanation that the silencing of genes at the Glu-A1 and Glu-B1 loci might result in the variation in the number of HMW subunits.
Dwarfing polish wheat is one of the accessions of T. turgidum sp. polonicum varieties. SDS-PAGE analysis showed that dwarfing polish wheat has two expressed HMW glutenin subunits (1Ax-dp and 1Bx-dp). The mobility of 1Ax-dp was situated that of between 1Ax2* and 1Dx5 subunit, while the mobility 1Bx-dp was similar to that of 1Bx7. These results indicate that two x-type HMW glutenin genes (1Ax-dp and 1Bx-dp) were expressed and two y-type HMW glutenin genes may be silenced in dwarfing polish wheat. This hypothesis is strengthened by the present sequence analysis. Sequence analysis indicated that 1Ax-dp and 1Bx-dp gene encode 836 and 795 amino acids, respectively. However, one of the y-type HMW glutenin genes, 1Ay-dp, contains an in-frame premature stops codon, which might result in the silencing of the HMW glutenin gene 1Ay (Forde et al., 1985; Jiang et al., 2009).
It is apparent that the slight differences of molecular weight among HMW glutenin subunits are mainly due to the central repetitive region size (Shewry et al., 2002). Anderson and Greene (1989) proposed that sequence alteration may also result from single base or single repeat changes and from indels within a repeat. Recent reports have also revealed that slight alterations of central repetitive region are shared by 1Dy12.4t, 1Dy12 and 1D12.1t (Thompson et al., 1985; Gianibelli and Solomon, 2003; Yan et al., 2004). In this study, sequence comparison indicated that both 1Ax-dp and 1Bx-dp had the primary structure which was identical to the published HMW glutenin genes (e.g., 1Ax2* and 1Bx7) with a high degree of homology. However, two hexapeptide insertions, one nonapeptide insertion, two hexapeptide deletions and one tripeptide deletion were found when the HMW glutenin gene 1Ax-dp from dwarfing polish wheat was compared with 1Ax1 from bread wheat. The comparison of the amino acid sequence of the 1Ax-dp and 1Ax subunits also showed that they differ by 16 amino acid substitutions. Apart from the amino acid substitutions, three insertions of hexapeptide (LGQEQQ, LGQGQP and GQQPEQ), one insertion of tetrapeptide (GQQQ), one insertion of nonapeptide (GYYSTSPQQ), one deletion of nonapeptide (GYYPTSSLQ) were found when the HMW glutenin gene 1Ax-dp was compared with 1Ax2*. In the central repetitive domain of 1Bx-dp, one hexapeptide insertion was observed. Therefore, the results suggested that the complete ORF of the HMW glutenin gene 1Ax-dp from dwarfing polish wheat differ from 1Ax1 and 1Ax2*, indicating that 1Ax-dp is a novel allele at the Glu-A1 loci. The HMW glutenin gene 1Bx-dp is also an additional allele of Glu-B1 loci.
Replication slippage, transposition, gene conversion and unequal crossing-over are alternate interpretations for the gene length modification of the HMW glutenin gene (Shewry et al., 2003). Comparisons of the amino acid sequences among subunits 1Dx2.2, 1Dx2.2* and 1Dx2 have indicated that the large size of 1Dx2.2 and 1Dx2.2* may result from the separate duplication events within the repetitive domain of Dx2 (Wan et al., 2005). A recent study presented by Yan et al. (2009) also suggested that the replication slippage may lead to the shorter sequence of Bx6** in comparison with Bx7. In this study, different insertions/deletions were found in the central repetitive region of the HMW glutenin subunits when the 1Ax-dp and 1Bx-dp gene from dwarfing polish wheat was compared with 1Ax1 and 1Bx7 from bread wheat, respectively. It is suggested that the insertion/deletion events within the 1Ax-dp and 1Bx-dp gene may responsible for the replication slippage of 1Ax1 and 1Bx7, respectively.
Phylogenetic analysis of the HMW subunit amino acid sequences clearly distinguished the subunits of different genomes, including the subunits of A, B and D genomes. 1Ax-dp, 1Ax1 and 1Ax2* were in one group. 1Bx-dp was grouped with 1Bx7 and all the subunits from the Glu-B1 loci formed a group. Both the analysis of amino acid sequences and the phylogenetic tree suggested that 1Ax-dp and 1Bx-dp from the dwarfing polish wheat were most closely related to 1Ax1 and 1Ax2* and 1Bx7 from Triticum aestivum, respectively.
The HMW glutenin subunits have been closely associated with breadmaking quality. 1Ax2* may be correlated to the extensibility of flour (Branlard and Dardevet, 1985). The 1Ax-dp subunit of present study was a novel member of the HMW glutenin subunit protein family. 1Ax-dp was highly related to 1Ax and 1Ax2* in amino acid sequence and might have a positive effect on the processing quality of dough.
ACKNOWLEDGMENTS
The authors are thankful to the National Natural Science Foundation of China (No. 30670150 and 30870154) and the Education Committee and the Science and Technology Committee of Sichuan Province for financial supports. The GeneBank accession number for the ORFs of 1Ax-dp, 1Bx-dp and 1Ay-dp is GU177739, AY722709 and AY722710, respectively.
REFERENCES
- Anderson, O.D. and F.C. Greene, 1989. The characterization and comparative analysis of high-molecular-weight glutenin genes from genomes A and B of a hexaploid bread wheat. Theor. Applied Genet., 77: 689-700.
CrossRefDirect Link - Altschul, S.F., W. Gish, W. Miller, E.W. Myers and D.J. Lipman, 1990. Basic local alignment search tool. J. Mol. Biol., 215: 403-410.
CrossRefPubMedDirect Link - Branlard, G. and D. Dardevet, 1985. Diversity of grain protein and bread wheat quality. II. Correlation between high molecular weight subunits of glutenin and flour quality characteristics. J. Cereal Sci., 3: 345-354.
CrossRef - Doyle, J.J. and J.L. Doyle, 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull., 19: 11-15.
Direct Link - Felsenstein, J., 1985. Confidence limits on phylogenies: An approach using the bootstrap. Evolution, 39: 783-791.
CrossRefDirect Link - Forde, J., J.M. Malpica, N.G. Halford, P.R. Shewry, O.D. Anderson, F.C. Greene and B.J. Miflin, 1985. The nucleotide sequence of a HMW subunit gene located on chromosome 1A of wheat (Triticum aestivum L.). Nucleic Acids Res., 13: 6817-6832.
PubMedDirect Link - Gianibelli, M.C. and R.G. Solomon, 2003. A novel y-type high Mr glutenin subunit (12.4t) present in Triticum tauschii. J. Cereal Sci., 37: 253-256.
CrossRef - Johansson, E. and G. Svensson, 1995. Contribution of the high molecular weight glutenin subunit 21* to breadmaking quality of Swedish wheats. Cereal Chem., 72: 287-290.
Direct Link - Liu, S.W., S.Y. Zhao, F.G. Chen and G.M. Xia, 2007. Generation of novel high quality HMW-GS genes in two introgression lines of Triticum aestivum/Agropyron elongatum. BMC Evol. Biol., 7: 76-76.
PubMed - Payne, P.I., 1987. Genetics of wheat storage proteins and the effect of allelic variation on bread-making quality. Annu. Rev. Plant Physiol., 38: 141-153.
CrossRefDirect Link - Shewry, P.R., N.G. Halford, P.S. Belton and A.S. Tatham, 2002. The structure and properties of gluten: An elastic protein from wheat grain. Phil. Trans. R. Soc. Lond. B, 357: 133-142.
CrossRefDirect Link - Shewry, P.R. and A.S. Tatham, 1997. Disulphide bonds in wheat gluten proteins. J. Cereal Sci., 25: 207-227.
CrossRef - Shewry, P.R., S.M. Gilbert, A.W.J. Savage, A.S. Tatham and Y.F. Wan et al., 2003. Sequence and properties of HMW subunit 1Bx20 from pasta wheat (Triticum durum) which is associated with poor end use properties. Theor. Applied Genet., 106: 744-750.
CrossRefPubMedDirect Link - Thompson, R.D., D. Bartels and N.P. Harberd, 1985. Nucleotide sequence of a gene from chromosome 1D of wheat encoding a HMW-glutenin subunit. Nucleic Acids Res., 13: 6833-6846.
CrossRefPubMedDirect Link - Wan, Y., D. Wang, P.R. Shewry and N.G. Halford, 2002. Isolation and characterization of five novel high molecular weight subunit of glutenin genes from Triticum timopheevi and Aegilops cylindrica. Theor. Applied Genet., 104: 828-839.
CrossRefDirect Link - Wan, Y.F., Z.H. Yan, K.F. Liu, Y.L. Zheng and R.D. Ovidio et al., 2005. Comparative analysis of the D genome-encoded high-molecular weightsubunits of glutenin. Theor. Applied Genet., 111: 1183-1190.
CrossRefPubMedDirect Link - Yan, Y., J. Zhang, Y. Xiao, J. Yu and Y. Hu et al., 2004. Identification and molecular characterization of a novel y-type Glu-Dt1 glutenin gene of Aegilops tauschii. Theor. Applied Genet., 108: 1349-1358.
PubMed - Yan, Z.H., S.F. Dai, D.C. Liu, Y.M. Wei, J.R. Wang and Y.L. Zheng, 2009. Isolation and characterization of a novel Glu-Bx HMW-GS allele from tibet bread wheat landrace. Int. J. Agric. Res., 4: 38-45.
CrossRefDirect Link - Yan, Z.H., Y.F. Wan, K.F. Liu, Y.L. Zheng and D.W. Wang, 2002. Identification of a novel HMW glutenin subunit and comparison of its amino acid sequence with those homologous subunits. Chinese Sci. Bull., 47: 220-225.
Direct Link - Yan, Z.H., Y.M. Wei, J.R. Wang, D.C. Liu, S.F. Dai and Y.L. Zheng, 2006. Characterization of two HMW glutenin subunit genes from Taenitherum Nevski. Genetica, 127: 267-276.
CrossRefPubMedDirect Link - Jiang, Q.T., Y.M. Wei, F. Wang, J.R. Wang, Z.H. Yan and Y.L. Zheng, 2009. Characterization and comparative analysis of HMW glutenin 1Ay alleles with differential expressions. BMC Plant Biol., Vol. 9.
CrossRefDirect Link