
Usman Pato
Department of Agricultural Technology, Faculty of Agriculture, Universitas Riau, Kampus Bina Widya KM 12.5 Simpang Baru, Pekanbaru Indonesia
LiveDNA: 62.29464
Yusmarini Yusuf
Department of Agricultural Technology, Faculty of Agriculture, Universitas Riau, Kampus Bina Widya KM 12.5 Simpang Baru, Pekanbaru Indonesia
Shanti Fitriani
Department of Agricultural Technology, Faculty of Agriculture, Universitas Riau, Kampus Bina Widya KM 12.5 Simpang Baru, Pekanbaru Indonesia
Tartila
Department of Agricultural Technology, Faculty of Agriculture, Universitas Riau, Kampus Bina Widya KM 12.5 Simpang Baru, Pekanbaru Indonesia
Rahma Yeni
Department of Agricultural Technology, Faculty of Agriculture, Universitas Riau, Kampus Bina Widya KM 12.5 Simpang Baru, Pekanbaru Indonesia
Fani Fadillah
Department of Agricultural Technology, Faculty of Agriculture, Universitas Riau, Kampus Bina Widya KM 12.5 Simpang Baru, Pekanbaru Indonesia
Latifa Husnanini
Department of Agricultural Technology, Faculty of Agriculture, Universitas Riau, Kampus Bina Widya KM 12.5 Simpang Baru, Pekanbaru Indonesia







ABSTRACT
Background and Objective: Bacteriocin, a peptide produced by Lactic Acid Bacteria (LAB) widely used as a natural and safe preservative in food products. This study aimed to determine the optimal incubation conditions and medium contents to increase the growth and antimicrobial activity of Pediococcus pentosaceus strain 2397 against Staphylococcus aureus. Materials and Methods: Pediococcus pentosaceus strain 2397 was used to study the antimicrobial activity against S. aureus FNCC-15. The cell-free supernatant and bacteriocin's antimicrobial activity from strain 2397 was carried out by the well diffusion method. Results: The results showed that the incubation time, the pH of the growth medium, the concentration of starter, the addition of various nitrogen and carbon sources affected the growth and antimicrobial activity of Pediococcus pentosaceus strain 2397. Optimal conditions for growth and increased antimicrobial activity of strain 2397 against S. aureus were obtained at 24 hrs incubation time, starter concentration 5.0%, medium pH 6, addition of N sources and C sources. Conclusion: Incubation conditions and medium composition determined the growth and antimicrobial activity of P. pentosaceus strain 2397 against S. aureus.
PDF Abstract XML References Citation
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Usman Pato, Yusmarini Yusuf, Shanti Fitriani, Tartila, Rahma Yeni, Fani Fadillah and Latifa Husnanini, 2021. Enhancement of the Growth and Antimicrobial Activity of Pediococcus pentosaceus Strain 2397 against Staphylococcus aureus. Biotechnology, 20: 8-14.
DOI: 10.3923/biotech.2021.8.14
URL: https://scialert.net/abstract/?doi=biotech.2021.8.14
DOI: 10.3923/biotech.2021.8.14
URL: https://scialert.net/abstract/?doi=biotech.2021.8.14
INTRODUCTION
Staphylococcus aureus is a commensal and opportunistic pathogen that can cause a broad spectrum of infections, from superficial skin infections to severe and potentially fatal, invasive diseases in humans and animal1. S. aureus can grow in a wide temperature range, from 7-48.5°C with an optimum temperature of 30-37°C, pH in the range of 4.2-9.3 with an optimum pH of 7-7.5 and concentrations of NaCl up to 15%. These bacteria can also survive in dry and stressful environments, such as the human nose and on the skin and inanimate surfaces such as clothing and surfaces2. These characteristics support the growth and contamination of S aureus in many food products3.
Some S. aureus strains can produce staphylococcal enterotoxins (SEs) and SEs are the causative agents of staphylococcal food poisonings4. This SEs is one of those compounds causing Staphylococcal Food Poisoning (SFP) by S. aureus5. The SEs are grouped in a family of nine main serologic heat-stable enterotoxin types: SEA, SEB, SEC, SED, SEE, SEG, SEH, SEI and SEJ6. It was estimated that about 20-30% of the human population are carriers for S. aureus7, which coexist with human micro biota and cause abscesses and infections of the skin, respiratory system and Staphylococcal food-borne disease8,9. Heating can kill vegetative cells of S. aureus but cannot deactivate SE in food products, thus, SE is still able to cause SFP10. Therefore, the growth of S. aureus in food must be prevented, for example, through the use of preservatives. Synthetic chemical preservatives have long been used in the preservation of various food products11. However, these chemical preservatives are not always safe for human consumption, especially if their excessive use can cause minor and severe health problems such as allergies to cancer12. Therefore, it is necessary to find safe and natural preservatives for human consumption, such as bacteriocins. Bacteriocins are antimicrobial compounds that are synthesized ribosomally by many members of the Lactic Acid Bacteria (LAB). Bacteriocin is a polypeptide compound produced by LAB, which has various Molecular Weight(MB). Bacteriocins were divided into four groups, namely Group I lantibiotics and group II small hydrophobic heat-resistant peptides, which had MB <13 kDa, Group III large heat-labile proteins have MB> 30 kDa and Group IV bacteriocins complexes which show protein complex molecules with lipids or carbohydrates13. These bacteriocins have high stability, low toxicity and a narrow to a broad spectrum of activity, making them suitable for clinical applications and safe, natural food preservatives14-16. Some of the LAB genera, such as lactobacilli, lactococci, pediococci and others, can produce bacteriocins such as Pediococcus pentosaceus strain 2397. This LAB was isolated from a traditional fermented milk product called dadiah and it was identified initially as Streptococcus faecalis subsp. liquefaciens R-55 using API CH 5017, can inhibit the growth of Listeria monocytogenes18. The present study aimed to assess P. pentosaceus strain 2397 against S. aureus growth and antimicrobial activity.
MATERIALS AND METHODS
Study area: The present study was experimentally conducted to evaluate the growth and antimicrobial activity of Pediococcus pentosaceus strain 2397 against Staphylococcus aureus FNCC 15. The study was carried out at the Agricultural Product Analysis Laboratory, Faculty of Agriculture, Universitas Riau, Pekanbaru, Indonesia, from June-October, 2020.
Media and chemicals: The media used for the activation of Pediococcus pentosaceus strain 2397 and S. aureus FNCC 15 cultures and the antimicrobial test were MRS Broth (MRSB), Nutrient Broth (NB) and Nutrient Agar (NA). The chemicals used include ammonium sulfate, phosphate buffers and other chemicals. All the media and chemicals were purchased from Merck and Sigma Aldrich (Singapore).
Pediococcus pentosaceus strain 2397 and pathogenic bacteria: Pediococcus pentosaceus strain 2397 was isolated from dadiah from Bukittinggi, West Sumatra, Indonesia18. The gram-positive pathogenic bacterium used was Staphylococcus aureus FNCC 15 was obtained from the Laboratory of Microbiology, Center for Food and Nutrition Studies, Universitas Gadjah Mada, Yogyakarta, Indonesia.
Activation of the LAB culture and Staphylococcus aureus FNCC 15: The activation of Pediococcus pentosaceus strain 2397 and S. aureus cultures were carried out19. Each LAB culture was taken as much as 0.1 mL and put into a test tube containing 5 mL MRSB, then shaken evenly and incubated aerobically for 18 hrs at 37°C. The S. aureus was activated by inoculating 0.1 mL of the bacterial suspension into 5 mL NB, shaken evenly and incubated aerobically for 18 hrs at 37°C.
In vitro antimicrobial activity of LAB: The cultures of Pediococcus pentosaceus strain 2397 were grown in MRSB and incubated aerobically for 24 hrs at 37°C. The indicator bacterium, S. aureus, was grown in NB for 24 hrs at 37°C. The indicator bacterium of 100 μL was inoculated and then flattened with a hockey stick until it was dry. Then a well was built by using a sterile blue tip and was coated with sterile agar. As much as 100 μL of free-cell supernatants or bacteriocins were poured into a well then incubated at 37°C for 24 hrs. The diameter of a clear zone formed was measured20.
Effect of different incubation times on the growth and antimicrobial activity: To study the effect of the incubation time, the tested organisms were grown in MRS broth medium at various incubation times (12, 24, 36, 48 and 72 hrs), then incubated at 37°C21. After incubation, the bacterial growth and the antimicrobial activity of P. pentosaceus strain 2397 against S. aureus were analyzed20.
Effect of pH on the growth and antimicrobial activity: The effect of initial medium pH on growth and production of bacteriocin of selected isolates was tested. The MRS broth medium was prepared in 250 mL Erlenmeyer flasks at different pH as following 4, 5, 6, 7, 8 and 10 with 6 M HCl or 6 M NaOH and then autoclaved. Each flask was inoculated with inoculums of selected strains and incubated at 37°C for 24 hrs22. After incubation, the bacterial growth and the antimicrobial activity of P. pentosaceus strain 2397 against S. aureus were analyzed20.
Effect of starter concentrations on the growth and antimicrobial activity: Different five concentrations of starters (2.5, 5.0, 7.5 and 10.0%) were used for studying the effect of inoculums concentration on the growth and antimicrobial activity of strain R-55 against S. aureus21. After incubation, the bacterial growth and the antimicrobial activity of P. pentosaceus strain 2397 against S. aureus were analyzed20.
Effects of medium composition on the growth and antimicrobial activity: Sources of nutrients to support growth and antimicrobial activity used are N sources including peptone, beef extract, yeast extract and ammonium sulfate and C sources including glucose, fructose, sucrose, lactose and mannitol. The concentration of the origins of N or C used in this study was 2%, respectively23. After incubation at 37°C for 24 hrs, the bacterial growth and the antimicrobial activity of P. pentosaceus strain 2397 against S. aureus were analyzed20.
Production of bacteriocin: P. pentosaceus strain 2397 was propagated in MRSB by adding inoculum as much as 2.5% of the active culture and incubation at 37°C for 24 hrs. Then, the whole broth was centrifuged at 10,000 rpm for 15 min to get the supernatant. The supernatant separated from the cells was then added with 70% ammonium sulfate and put in the refrigerator (4°C) for 12 hrs to precipitate the protein. This mixture was centrifuged at 10,000 rpm at 4°C for 30 min to obtain crude bacteriocin24.
RESULTS AND DISCUSSION
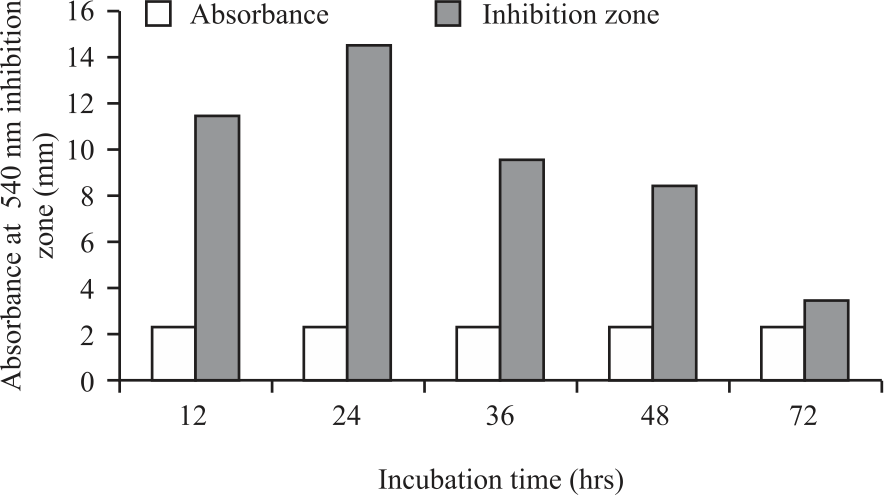
The growth and inhibition zone of cell-free supernatant of P. pentosaceus strain 2397 against Staphylococcus aureus is shown in Fig. 1.
The growth of strain 2397 increased with incubation time from 12-48 hrs, which was indicated by an increase in the medium's absorption. This fact is because the medium's nutrient content was still widely available for energy sources and cell division of strain 2397. The increase in the number of cells in the medium causes an increase in the medium's absorption value. However, the growth of strain 2397 decreased when incubated for 72 hrs. This fact is probably because the amount of nutrients in the medium has decreased. Primary metabolites have also been formed, such as lactic acid, which accumulated in medium and inhibited the growth of strain 2397. The L. casei subsp. casei R-68, LAB, which was also isolated from dadiah, grew well on skim milk medium with optimal growth achieved at 15 hrs after incubation time and at 18-21 hrs showed decreased LAB viability25. In kefir manufacturing, the best incubation time was obtained at 16 hrs26. This difference in optimal growth is due to different growing mediums. During growth, the LAB in a medium will produce lactic acid as the main metabolite product, which causes a decrease in the medium's pH and can inhibit the growth of LAB itself27.
 |
| Fig. 1: | Growth and antimicrobial activity of cell-free supernatant of P. pentosaceus strain 2397 at various incubation times against Staphylococcus aureus |
 |
| Fig. 2: | Growth and antimicrobial activity of cell-free supernatant of P. pentosaceus strain 2397 at various medium pH against Staphylococcus aureus |
 |
| Fig. 3: | Growth and antimicrobial activity of cell-free supernatant of P. pentosaceus strain 2397 at various starter concentrations against Staphylococcus aureus |
Although the growth of strain 2397 was almost the same at various incubation times, the antimicrobial activities differed. The highest zone of inhibition of cell-free supernatant strain 2397 was found at 24 hrs of incubation time, the lowest was found at 72 hrs of incubation times. This is probably because the medium's nutrients have been depleted for growth, so that there was little left to produce antimicrobial compounds, especially bacteriocin.
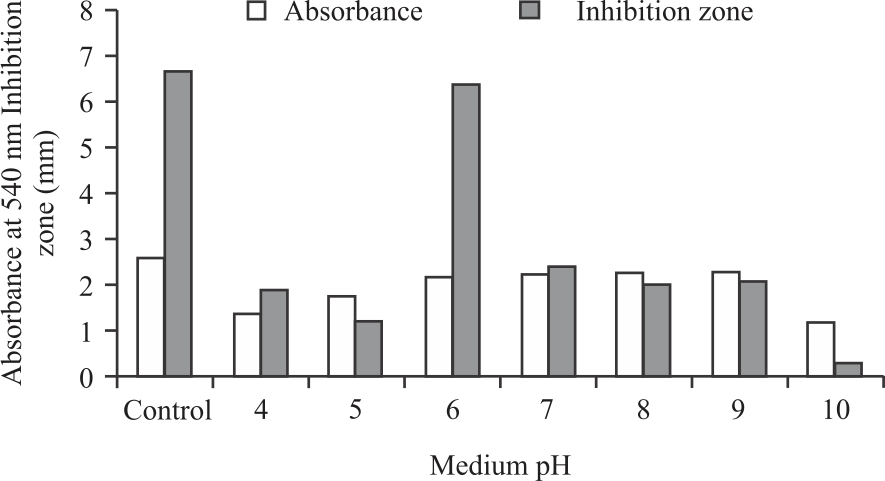
The Fig. 2 shows the growth of P. pentosaceus strain 2397 at various initial pH of the medium. This LAB was able to grow well in the pH range between 6-9 including control, which had pH 6.3. However, the growth of strain 2397 was not so good at acidic conditions of pH 3-5 and alkaline conditions of pH 10. The P. pentosaceus has a lower optimum temperature for growth (28-32°C) than P. acidilactici (40°C) but the latter grows at 50°C. The optimum pH for growth is 6.0-6.5. Half of the species grow at pH 4.2 and most of them (except P. damnosus) grow at pH 7.0. Pediococcus pentosaceus LB44 could demonstrate similar growth at pH 5.0-8.028.
Although P. pentosaceus strain 2397 could grow well in a pH range between 6-9, the highest antimicrobials were obtained in the range, especially in controls with a pH of 6.3 with an inhibition zone of 6.7 mm followed by a pH of 6 of 6.4 mm. Beyond this pH, the cell-free supernatant strain 2497 had very little antimicrobial activity against the growth of S. aureus. The results of this study are somewhat contradictory to the results of research by Kaur et al.28. They reported that P. pentosaceus LB44 could grow and produce bacteriocin under acidic and alkaline conditions at 37°C.
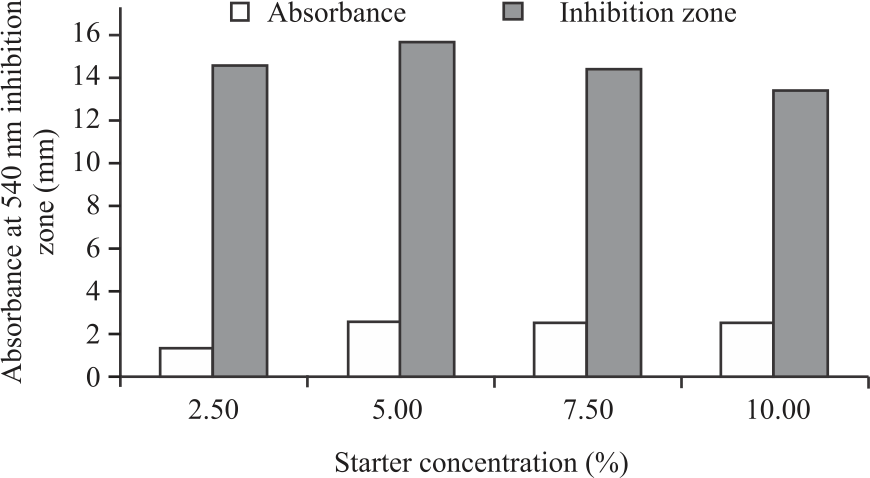
Figure 3 showed the effect of starter concentration on the growth of P. pentosaceus strain 2397 on MRS Both medium. The higher the starter concentration used, the lower the growth of strain 2397. The highest growth of strain 2397 was obtained using a starter concentration of 2.5% and the lowest at 10.0%. This is because the higher the concentration used, the greater the amount of LAB at the beginning of fermentation that competes for nutrients in the medium so that the LAB growth decreases, which is indicated by a decrease in absorbance. In kefir manufacturing, a starter concentration of 10.0% produces the best quality kefir in terms of pH and lactic acid production. The difference in the best concentration depends on the medium used26. In this study, the MRS Broth medium was used, while in the manufacture of kefir, milk was used as the growth medium so that there was a difference in the best starter concentration for microbial growth.
The antimicrobial activity of cell-free supernatant from strain 2397 against S. aureus showed almost the same inhibition zone. Still, the use of 5.0% starter showed a higher inhibition zone than other concentrations. The optimal growth of strain 2397 in a medium will produce metabolite compounds, especially secondary metabolites such as bacteriocin, which can inhibit the growth of S. aureus. Several previous researchers have reported the ability of LAB grown optimally in a medium to inhibit the growth of S. aureus29,30.
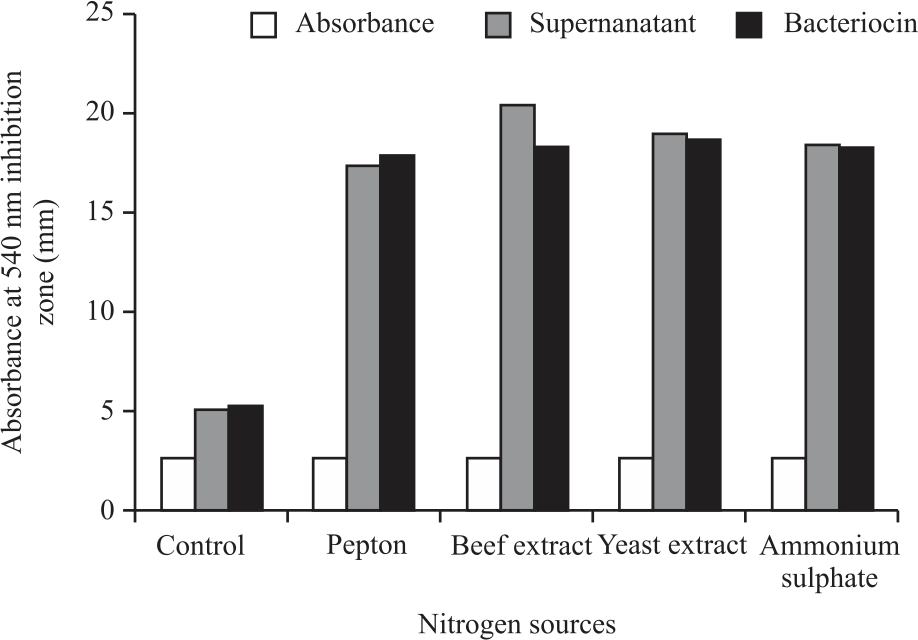
The growth and antimicrobial of cell-free supernatant and bacteriocin from strain 2397 grown at various nitrogen sources were presented in Fig. 4. The growth of strain 2397 showed almost the same absorbance values with or without different nitrogen sources. The absorbance value of the control (without the addition of N) was 2.564, while the addition of N sources such as ammonium sulfate, peptone, beef extract and yeast extract ranged from 2.586-2.723. This was due to the MRSB medium containing sufficient N growth of LAB, including strain 2397.
Although the growth showed relatively similar absorbance values, the addition of four types of N sources significantly increased the antibacterial activity of strain 2397 against S. aureus. The supernatant inhibition zone increased from 5.09 mm in the control to 20.34 mm in the beef extract. Likewise, the bacteriocin inhibition zone increased from 5.34 mm in the control to 18.67 mm in the yeast extract.
 |
| Fig. 4: | Growth and antimicrobial activity of cell-free supernatant and bacteriocin of P. pentosaceus strain 2397 at various nitrogen sources against Staphylococcus aureus |
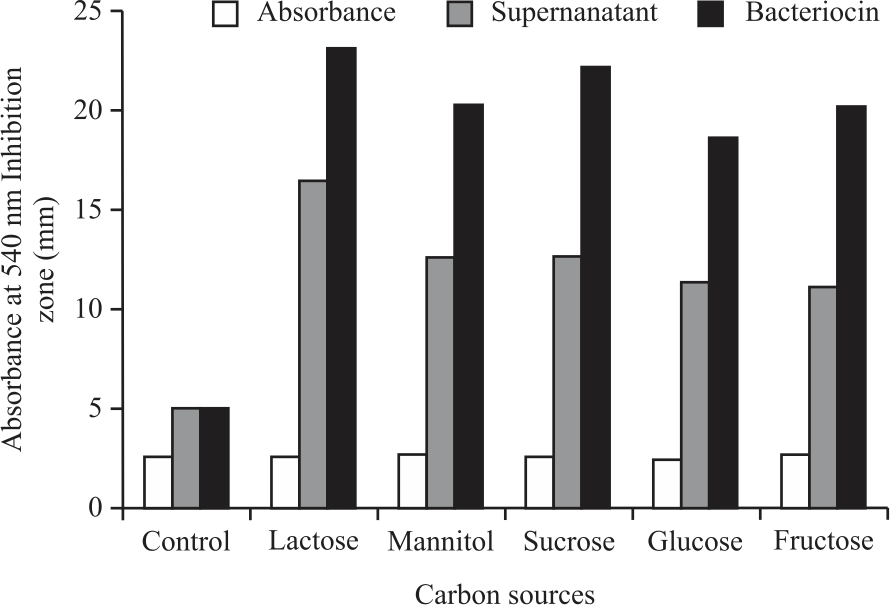 |
| Fig. 5: | Growth and antimicrobial activity of cell-free supernatant and bacteriocin of P. pentosaceus strain 2397 at various carbon sources against Staphylococcus aureus |
The increase in the inhibition zone between supernatant and bacteriocin showed almost the same values for the four N sources, from 17.34-20.34 mm for supernatant and from 17.92-18.67 mm for bacteriocin. These results indicate that the main compound in the supernatant that inhibits S. aureus growth was bacteriocin. This finding is in line with the results of previous research18.
The growth of strain 2397 with or without various carbon sources showed almost the same absorbance values (Fig. 5). The absorbance value of the control (without the addition of C) was 2.642, while for the addition of C sources such as glucose, sucrose, lactose, fructose and mannitol ranged from 2.533-2.772. These results indicate that MRSB contains sufficient C sources to support LAB's growth, including strain 2397. Although the absorbance values were almost the same, the addition of various types of C sources significantly increased the antimicrobial activity of strain 2397 against S. aureus. The zone of inhibition of the supernatant increased from 5.09 mm in the control to 16.50 mm in lactose. Likewise, the bacteriocin inhibition zone increased from 5.10 mm in the control to 23.17 mm in lactose. The addition of C source causes more sugar available that could be used by the strain 2397 as an energy source and to be used to form bacteriocins. This is supported by the fact that bacteriocins' microbial activity is much higher than that of cell-free supernatants.
Some researchers reported that bacteriocin isolated from various fermented foods could suppress S. aureus growth31. The molecular weight of bacteriocins isolated and purified from strain 2397 was 14.4 kDa. The bacteriocins produced by several lactic acid bacteria vary greatly depending on the genus and species. Some bacteriocins have a molecular weight of greater, less or almost the same as the molecular weight of bacteriocins produced by strain 2397. The bacteriocin's molecular weight from Lactobacillus plantarum isolated from Chinese pickle was nearly the same as the bacteriocin molecular weight produced by strain 2397, namely 16.5 kDa32. However, Lactobacillus lactic isolated from the marine environment produced bacteriocin with a molecular weight of 94 kDa, much greater than that produced by strain 2397. Most bacteriocins of LAB are peptide compounds with small molecular weights of less than 10 kDa15. Enterococcus faecalis KT11 produced bacteriocin with a molecular weight of 3.5 kDa31. The bacteriocin produced by Pediococcus pentosaceus zy B isolated from the intestine of Mimachlamys nobilis had a molecular weight of 2.22 kDa32,33.
CONCLUSION
It can be concluded that the incubation time, the pH of the growth medium, the concentration of starter, the addition of various nitrogen and carbon sources affected the growth and antimicrobial activity of Pediococcus pentosaceus strain 2397. Optimal conditions for growth and increased antimicrobial activity of strain 2397 against S. aureus were obtained at 24 hrs incubation time, medium pH 6, starter concentration 5.0%, addition of N sources such as peptone, beef extract, yeast extract, ammonium sulfate and C sources such as lactose and sucrose.
SIGNIFICANCE STATEMENT
This study discovered the optimal incubation conditions and medium composition to enhance the growth and antimicrobial activity of P. pentosaceus strain 2397 that can be beneficial for inhibiting S. aureus FNCC-15. This study will help the researchers to uncover natural preservation agents from local lactic acid bacteria. Thus a new theory on optimization of bacteriocin production may be achieved.
ACKNOWLEDGMENT
The research was funded by the Directorate of Research and Community Service, Ministry for Research, Technology and Higher Education of the Republic of Indonesia with Contract Number: 421/UN.19.5.1.3/PT.01.03/2020.
REFERENCES
- Kadariya, J., T.C. Smith and D. Thapaliya, 2014. Staphylococcus aureus and staphylococcal food-borne disease: An ongoing challenge in public health. BioMed Res. Int. Vol. 2014.
CrossRefDirect Link - Chaibenjawong, P. and S.J. Foster, 2011. Desiccation tolerance in Staphylococcus aureus. Arch. Microbiol., 193: 125-135.
CrossRefDirect Link - Le Loir, Y., F. Baron and M. Gautier, 2003. Staphylococcus aureus and food poisoning. Genet. Mol. Res., 2: 63-76.
Direct Link - Taddesse, Z., M. Tiruneh and M. Gizachew, 2014. Staphylococcus aureus and its antimicrobial susceptibility. Glob. J. Med. Res. Microbiol. Pathol., 14: 1-8.
Direct Link - Rasooly, R. and M. Friedm, 2012. Food Compounds Inhibit Staphylococcus aureus Bacteria and the Toxicity of Staphylococcus Enterotoxin A (SEA) Associated with Atopic Dermatitis. In: Atopic Dermatitis Disease Etiology and Clinical Management, Esparza-Gordillo, J. (Ed.)., InTech, London, UK, pp: 387-404,.
CrossRefDirect Link - Argudin, M.A., M.C. Mendoza and M.R. Rodicio, 2010. Food poisoning and Staphylococcus aureus enterotoxins. Toxin, 2: 1751-1773.
CrossRefDirect Link - Tong, S.Y., J.S. Davis, E. Eichenberger, T.L. Holland and V.G. Fowler, 2015. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations and management. Clin. Microbiol. Rev., 28: 603-661.
CrossRefDirect Link - Carroll, D.E., I. Marr, G.K.L. Huang, D.C. Holt, S.Y.C Tong and C.S. Boutlis, 2017. Staphylococcus aureus prostatic abscess: A clinical case report and a review of the literature. BMC Infect. Dis., 17: 1-7.
CrossRefDirect Link - Vickery, T.W., V.R. Ramakrishnan and J.D. Suh, 2019. The role of Staphylococcus aureus in patients with chronic sinusitis and nasal polyposis. Curr. Allergy Asthma Rep., Vol. 19.
CrossRefDirect Link - Hu, D.L. and A. Nakane, 2014. Mechanisms of staphylococcal enterotoxin-induced emesis. Eur. J. Pharmacol., 722: 95-107.
CrossRefDirect Link - Sharif, Z, F. Mustapha, J. Jai, N.M. Yusof and N. Zaki, 2017. Review on methods for preservation and natural preservatives for extending the food longevity. Chem. Eng. Res. Bull., 19: 145-153.
CrossRefDirect Link - Dicks, L.M.T., A.D.P. van Staden and B. Klumperman, 2017. Bacteriocins and Nanotechnology, In: Functionalized Nanomaterials for the Management of Microbial Infection, Boukherroub, R., S. Szunerits and D. Drider (Eds.)., Elsevier, New York, pp: 267-288.
CrossRefDirect Link - Saeed, M., W.A. Khan, M.A. Shabbir, M.I. Khan, M.A. Randhawa and I. Yasmin, 2014. Bacteriocins as a natural antimicrobial agent in food preservation: A review. Pak. J. Food Sci., 24: 244-255.
Direct Link - Kitagawa, N., T. Otani and T. Inai, 2019. Nisin, a food preservative produced by Lactococcus lactis, affects the localization pattern of intermediate filament protein in HaCaT cells. Anat. Sci. Int., 94: 163-171.
CrossRefDirect Link - Zacharof, M.P. and R.W. Lovitt, 2012. Bacteriocins produced by lactic acid bacteria a review article. APCBEE Procedia, 2: 50-56.
CrossRefDirect Link - Bharti, V., A. Mehta, S. Singh, N. Jain, L. Ahirwal and S. Mehta, 2015. Bacteriocin: A novel approach for preservation of food. Int. J. Pharm. Pharm. Sci., 17: 20-29.
Direct Link - Hosono, A., T. Tanabe and H. Otani, 1990. Binding properties of lactic acid bacteria isolated from kefir milk with mutagenic amino acid pyrolyzates. Milchwissenschaft, 45: 647-651.
Direct Link - Pato, U., Y. Yusuf, S. Fitriani, N.N. Jonnadi, M.S. Wahyuni, J.A. Feruni and I. Jaswir, 2020. Inhibitory activity of crude bacteriocin produced by lactic acid bacteria isolated from dadih against Listeria monocytogenes. Biodiversitas J. Biol. Divers., 21: 1295-1302.
CrossRefDirect Link - Hosono, A., R. Wardojo and H. Otani, 1990. Inhibitory effects of lactic acid bacteria from fermented milk on the mutagenicities of volatile nitrosamines. Agric. Biol. Chem., 54: 1639-1643.
CrossRefDirect Link - Pato, U., V.S. Djohan, F. Khairunnisa, R.D. Hasibuan and T. Scopus, 2017. Antibiotic resistance and antibacterial activity of dadih originated Lactobacillus casei subsp. casei R-68 against food borne pathogens. Asian J. Microbiol. Biotechnol. Environ. Sci., 19: 577-587.
Direct Link - Danial, E.N., S.H.M. Al-Zahrani and Z.H.M. Al-Mahmoudi, 2016. Enhancement of novel extracellular bacteriocin production by media optimization using LAB isolate from meat. J. Appl. Pharma. Sci., 6: 20-27.
CrossRefDirect Link - Sure KP., P.V. Kotnis, P.K. Bhagwat, R.C. Ranveer, P.B. Dandge and A.K. Sahoo, 2016. Production and characterization of bacteriocin produced by Lactobacillus Viridescence (NICM 2167). Braz. Arch. Biol. Technol., 59: 1-7.
CrossRefDirect Link - Fahim, H.A., W.M.A.E. Rouby, A.O. El-Gendy, A.S. Khairalla, I.A. Naguib and A.A. Farghali, 2017. Enhancement of the productivity of the potent bacteriocin avicin A and improvement of its stability using nanotechnology approaches. Sci. Rep., 7: 1-13.
CrossRefDirect Link - Sankar, N.R., V.D. Priyank, P.S. Reddy, P. Rajanikanth, V.K. Kumar and M. Indira, 2012. Purification and characterization of bacteriocin produced by Lactobacillus plantarum isolated from cow milk. Int. J. Microbiol. Res., 3: 133-137.
Direct Link - Pato, U., V.S. Johan, A.R. Raidinawan, A.A. Ginting and I. Jaswir, 2020. Viability and quality of fermented milk made using local and commercial starters during fermentation and cold storage. J. Agr. Sci. Tech., 22: 1473-1485.
Direct Link - Othman, H., A. Elbab, H. Abdelsame, A. Abdou and E. Ibrahim, 2017. Evaluation the effect of Lactobacillus acidophilus probiotic culture over Staphylococcus aureus during the production and storage of acidophilus yoghurt. Benha. Vet. Med. J., 32: 127-131.
CrossRefDirect Link - Rakhmanova, A., Z.A. Khan and K. Shah, 2018. A mini review fermentation and preservation: Role of lactic acid bacteria. MOJ Food Process Technol., 6: 414-417.
CrossRefDirect Link - Kaur, R. and S.T. Kumar, 2017. Optimization of culture conditions for bacteriocin production by soil isolates Pediococcus pentosaceus LB44 and Weissella confusa LM85. Int. J. Infect., Vol. 4.
CrossRefDirect Link - Karska-Wysocki, B., M. Bazo and W. Smoragiewicz, 2010. Antibacterial activity of Lactobacillus acidophilus and Lactobacillus casei against methicillin-resistant Staphylococcus aureus (MRSA). Microbiol. Res., 165: 674-686.
CrossRefPubMedDirect Link - Sari, M. and D. Suryanto, 2018. Antimicrobial activity of lactic acid bacteria isolated from bekasam against Staphylococcus aureus ATCC 25923, Escherichia coli ATCC 25922 and Salmonella sp. IOP Confer. Ser.: Earth Environ. Sci., Vol. 130.
CrossRefDirect Link - Abanoz, H.S. and B. Kunduhoglu, 2018. Antimicrobial activity of a bacteriocin produced by Enterococcus faecalis KT11 against some pathogens and antibiotic-resistant bacteria. Korean J. Food Sci. Anim. Resour., 38: 1064-1079.
CrossRefDirect Link - Zhou, F., H. Zhao, F. Bai, D. Piotr, Y. Liu and B. Zhang, 2014. Purification and characterisation of the bacteriocin produced by Lactobacillus plantarum, isolated from Chinese pickle. Czech J. Food Sci., 32: 430-436.
CrossRefDirect Link - Zhang, Y., J. Yang, Y. Liu, Y. Wu and Z. Fang et al., 2020. A novel bacteriocin PE-ZYB1 produced by Pediococcus pentosaceus zy-B isolated from intestine of Mimachlamys nobilis: Purification, identification and its anti-listerial action. LWT, Vol. 118.
CrossRefDirect Link