
Vartika Sinha
Biomembrane Toxicology Division, Indian Institute of Toxicology Research, Lucknow, India
Ritesh Mishra
Biomembrane Toxicology Division, Indian Institute of Toxicology Research, Lucknow, India
Abhinav Kumar
Molecular Virology Laboratory, National Botanical Research Institute, Lucknow Council of Scientific and Industrial Research, New Delhi, India
Ambrose Kannan
Biomembrane Toxicology Division, Indian Institute of Toxicology Research, Lucknow, India
Raj K. Upreti
Biomembrane Toxicology Division, Indian Institute of Toxicology Research, Lucknow, India
Biotechnology
Year: 2011 | Volume: 10 | Issue: 1 | Page No.: 101-107







ABSTRACT
The aim of the study was to develop arsenite resistance in Lactobacillus acidophilus and to show the presence of arsenical resistant gene in its plasmid. The arsH gene and its homologs are fundamental to confer high level of arsenite resistance in bacteria. In the present study in vitro resistance against arsenite (up to 32 ppm) was developed in Lactobacillus acidophilus and the gene responsible for high level of arsenite resistance (arsH gene) was cloned and sequenced. Concomitantly, arsenite Minimal Inhibitory Concentration (MIC), growth phase studies, antibiotic and heavy metal tolerance were tested for this strain. The arsenite-resistant strain IITR-RKU1 showed similar growth phase patterns to that of normal parent strain in the absence or in presence of arsenite in the media. PCR using specific primers of arsH gene showed the presence of 606 bp arsH gene on a 23 kb (kilo base) plasmid of Lactobacillus acidophilus. The putative product of this gene is 202 amino acids long, having calculated molecular weight of 23 kDa (kilo Dalton) and isoelectric point of 6.0. The amino acid sequence of arsH of L. acidophilus showed 99% identity with arsenical resistance protein of Acetobacter sp. Comparison of the predicted amino acid sequence of arsH with CD (Conserved Domain) server revealed the signatures of Flavin Mono Nucleotide (FMN) reductase protein. So far arsH, in general, has been reported in Gram-ve bacterial isolates. This is for the first time arsH has been shown to be present in L. acidophilus plasmid pRKU101.
PDF Abstract XML References Citation
Received: August 27, 2010;
Accepted: December 26, 2010;
Published: February 22, 2011
How to cite this article
Vartika Sinha, Ritesh Mishra, Abhinav Kumar, Ambrose Kannan and Raj K. Upreti, 2011. Amplification of arsH Gene in Lactobacillus acidophilus Resistant to Arsenite. Biotechnology, 10: 101-107.
DOI: 10.3923/biotech.2011.101.107
URL: https://scialert.net/abstract/?doi=biotech.2011.101.107
DOI: 10.3923/biotech.2011.101.107
URL: https://scialert.net/abstract/?doi=biotech.2011.101.107
INTRODUCTION
Arsenic, which is ubiquitous in the environment, has become a world wide health problem. Acute and chronic arsenic exposure via drinking water/ground water has been reported in many countries especially in Bengal Delta (Peters et al., 1999; Berg et al., 2001; Chowdhury et al., 2006; Mahfuzar, 2006; Sohel et al., 2009; Bose et al., 2010). Arsenic mainly occurs in two inorganic form, arsenate (As-V) and arsenite (As-III) and the trivalent form is more toxic than the pentavalent form (Toxicological Profile for Arsenic, 2005). Due to natural abundance of arsenic in the environment, representatives from various bacterial genera have developed different resistance mechanisms for arsenic compounds (Silver, 1996; Mukhopadhyay et al., 2002). Resistance to arsenical compounds in bacteria is mediated by ars operons which are either plasmid or chromosomally encoded. In all this operon, arsH gene has been reported to be important to confer high levels of arsenite resistance (Branco et al., 2008).
Recently it has been inferred that various geogenic and anthropogenic factors are responsible for arsenic contamination in the environment and in turn water and food are the primary sources of arsenic contamination in various organisms (Bhakta et al., 2010). Lactobacilli are one of the most abundant bacteria of mammalian gastrointestinal (GI) tract and hence susceptible for arsenic toxicity. Therefore, arsenic tolerant friendly bacteria like Lactobacillus can become a useful probiotic. However, heavy metal resistance in a number of different bacteria is known to be present together with antibiotic resistances (Nies, 1999; Kamala-Kannan and Lee, 2008). Thus, the sensitivity of arsenite-resistant Lactobacillus strains for antibiotics also needs due consideration. These bacteria can also be used in the bioremediation of arsenic toxicity (Mateos et al., 2006). Therefore, in the present study arsenite resistance was developed in Lactobacillus acidophilus MTCC 447 and attempts were made to detect arsH gene.
MATERIALS AND METHODS
Source of the organism and MIC determination: Pure culture of L. acidophilus (MTCC 447) was procured from M.T.C.C. (Microbial Type Culture Collection and Gene Bank; Institute of Microbial Technology (IMTECH), Chandigarh-160036, India) and maintained in Lactobacillus MRS broth (HiMedia, India). The Minimal Inhibitory Concentration (MIC) of arsenite (As-III; Sodium m-arsenite from Sigma-Aldrich) was determined by the standard dilution method (NCCLS, 2002). Viability was tested by Colony Forming Unit (CFU) on agar plates.
In vitro development of arsenite resistance: The L. acidophilus was grown in Lactobacillus MRS broth at 37°C for 18 h and approximately 5x107 CFU were inoculated into a series of tubes containing 9.9 mL of MRS broth with arsenite concentrations consisting of doubling dilutions below and above the MIC. The culture was subjected to 15 serial passages with stepwise increasing the concentration of arsenite. After 15 cycles, a bacterial mutant designated as strain IITR-RKU1 capable of growing in 32 ppm of arsenite was obtained. The arsenite-resistant strain was kept frozen at 40°C and time to time sub cultured to assess the stability of resistance.
Growth phase studies: The parent strain and the arsenite-resistance developed strain (IITR-RKU1) were grown in MRS media at 37°C. Bacterial growth was measured at different time intervals up to 30 h using turbidimetry at 610 nm. In case of resistant strain, growth measurements were carried out in the absence and presence of arsenite in the media.
Antibiotic susceptibility test: The parent and the arsenite-resistance, developed strains were tested for antibiotic sensitivity following the National Committee for Clinical Laboratory (NCCL) standard disk diffusion method (Bauer et al., 1966). The following antibiotic disks from HiMedia, India were used: Amoxycillin (25 μg), Chloramphenicol (25 μg), Ciprofloxacin (10 μg), Gentamicin (10 μg), Kanamycin (30 μg), Norfloxacin (10 μg), Novabiocin (30 μg), Erythromycin (10 μg), Streptomycin (10 μg) and Teicoplanin (30 μg).
Plasmid isolation: Plasmid DNA of arsenite-resistant L. acidophilus was extracted according to the method of Klaenhammer (1984). Isolated DNA was dissolved in 50 μL of sterilized double distilled water. Electrophoresis was carried out in 1% agarose gel containing ethidium bromide.
Amplification of arsenical resistant gene: After plasmid isolation, PCR (Polymerase Chain Reaction) was performed to amplify arsenical resistance gene. The consensus primer sequence used for amplification of arsH gene was as follows: F-5’ - GCTGCTCTACGGCTCGCTGC 3’, R-5’- CACAGGCTTTCCGGGAGGCG 3’. Primers were synthesized and supplied by Integrated DNA Technologies, Belgium. The 50 μL PCR reaction mixture consisted of 10XPCR buffer, 25 mM MgCl2, 2 mM dNTP, 5 U Taq DNA polymerase (Fermentas, Germany), 1 μL each of forward and reverse primer and 4 μL of template. Amplification consisted of one cycle at 95°C for 5 min and 34 cycles at 94°C for 1 min and annealing was performed at 60°C for 1 min. This step was followed by extension at 72°C for 2 min and a final cycle at 72°C for 15 min. Amplified products were loaded in 1% agarose gel to access the quality of the product. The concentration of product was checked on Nanodrop (Thermo Scientific, USA).
Cloning, transformation and sequencing: A band of approximately 650 bp was recovered from the Agarose gel and purified using HiPura Gel extraction kit (HiMedia Pvt Ltd. India) following the manufacturer’s instructions and then cloned into pDrive vector (Qiagen, USA). The recombinant plasmid was then transformed into Escherichia coli DH5α cells. Appeared colonies were checked by restriction digestion and PCR for the potential presence of the gene insert. The positive clones were sequenced using Automated Sequencer 3730 XL DNA Analyzer, Applied Biosystems, USA.
Computer analysis: The nucleotide sequence was taken into account for similarity search using BLASTn search program (http://www.ncbi.nlm.nih.gov/BLAST/). Complete nucleotide sequence of arsH (arsenical resistance gene) was aligned with various bacterial arsH gene by using ClustalW program. Gene characterization of arsH gene was carried out using AnnHyb 4.0 software. Prediction of protein physio-chemical properties was carried out using ProtParam Tools as described by Gasteiger et al. (2003). Protein conserved domain was searched using CD server (www.ncbi.nlm.nih.gov). Phylogenetic analysis of arsH and deduced amino acid sequences were performed using MEGA version 4.0 software (Tamura et al., 2007). Default parameter was used, one character-based algorithm (Maximum Parsimony) and two distance-based algorithms (Minimum Evolution and Neighbour-Joining). A consensus dendogram was generated using bootstrap value of 1000 replicates for these algorithms.
RESULTS AND DISCUSSION
In vitro development of arsenite resistance: Arsenite resistance was developed in normal Lactobacillus acidophilus by chronological chronic exposures to the arsenite. Bacteria, in general, have good ability to adapt to environment and develop resistance to metals and antibiotics. This ability is mainly because of the presence of plasmid (Silver, 1996). Sub-culturing in the presence of arsenite gave increased MICs, with MICs rising from 0.05-1.0 to 2.0-32 mg L-1 after 6 to 15 sub-cultures.
Growth phase studies: Arsenite-resistant strain IITR-RKU1 followed entire growth phase patterns very similar to that of wild parent strain when grown either in the absence or in presence of arsenite (up to 32 ppm) in the media. The wild parent strain did not grow when grown in presence of higher than minimal inhibitory concentration of arsenite.
Antibiotic susceptibility test: We observed that arsenite-resistant strain did not acquire resistance against various antibiotics. Concomitantly, results also revealed that the arsenite-resistant mutant was sensitive to Hg2+, Cd2+, Pb2+ and Cr6+. The mutant strain IITR-RKU1 apparently showed all morphological and phenotypic characteristics similar to wild strain except for arsenite resistance. This selective change may be because of exposure to the arsenite during growth and the resistance mechanism responsible for arsenite may not be effective against other chemicals. Furthermore, to investigate the possible intracellular and membrane alterations following the development of in vitro arsenite resistance, various biochemical toxicity parameters as described by Upreti et al. (2007, 2008) were analyzed and compared with wild strain. Results revealed similarities in all parameters of the arsenite-resistant strain as compared to the wild strain (Upreti et al., 2011).
Plasmid isolation and amplification of arsenical resistance gene: Agarose gel electrophoresis of arsenite-resistant L. acidophilus strain IITR-RKU1 showed the presence of a 23 kb plasmid (Fig. 1) and it was designated as plasmid pRKU101.
Cloning, transformation and sequencing: When primer specific for arsH gene was used, a band of 650 bp was obtained in both wild and mutant strain. The eluted 650 bp band was cloned and sequenced. The sequence of arsH gene obtained from both wild and mutant showed 100% homology, suggesting no mutations in the arsH gene, probably the resistance to higher concentrations was developed in mutant strain due to change in other region of ars operon. Further studies in this direction are in progress. The sequence of mutant strain was deposited in GenBank (Accession No: HM003402).
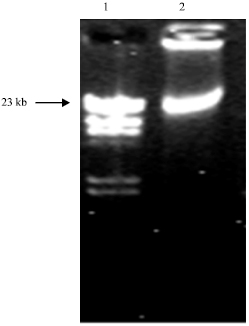 | |
| Fig. 1: | Agarose gel electrophoresis of plasmid DNA of L. acidophilus. Lane 1- Lambda Hind III, Lane 2- L. acidophilus plasmid |
Computer analysis: The gene sequence (Fig. 2) showed high degree of similarity with bacterial arsH (arsenical resistance gene). The arsH gene of our strain showed homology score of 96% with arsH gene of Methylobacterium nodulans ORS 2060; 90% with Pseudomonas putida KT 2440 and Pseudomonas putida F1, complete genome; 87% with Pseudomonas putida GB-1 and 78% with Methylobacterium extorquens DM4. The putative product of gene (arsenical-resistant protein- ArsH) was 202 amino acids long and this homologue was 94% identical with ArsH of Azospirillum sp. 81% with Acetobacter pasteurians IFO and Phenylobacterium zucineum HLK1; 80% with Acidiphilium cryptum JF-5; and 79% with Azorhizobium caulinodans.
Phylogenetic analysis of arsH with arsenical resistance gene showed that it was similar to the arsH of various gram negative bacteria having closest relationship with Methylobacterium extorquens DM4 (Fig. 3). Concomitantly, ArsH also showed its similarity to the ArsH of gram negative bacterial species having 99% nearness to Acetobacter sp. (Fig. 4). NCBI conserved domain search results suggested that a FMN reductase domain was found within 28-193 residue of ArsH protein (Fig. 5). Ours is the first report on arsH gene, which has been found in the plasmid of L. acidophilus strain IITR-RKU1.
Arsenic and its compounds are widespread in nature at near toxic levels since the origin of life. Earlier studies on arsenic resistant bacteria have been carried out in various bacterial strains, which were isolated from the arsenic rich environment (Salam et al., 2009).
 | |
| Fig. 2: | DNA sequence of the 606-bp arsH of L. acidophilus IITR-RKU1 plasmid pRKU101. The predicted amino acid sequences of encoded polypeptides are shown in the single-letter code |
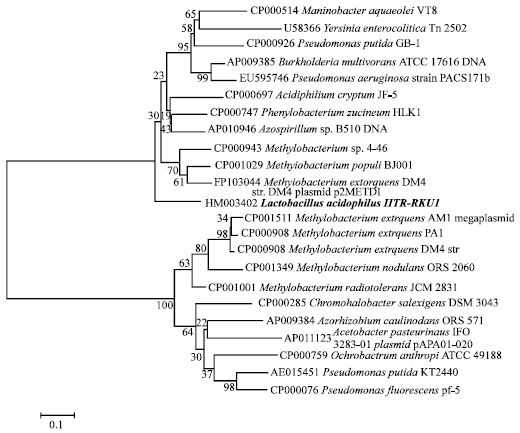 | |
| Fig. 3: | Phylogenetic analysis of arsH nucleotides of L. acidophilus IITR-RKU1 with arsH genes of other bacteria |
The arsH gene and its homologs are the frequent part of arsenic resistance mechanisms in bacteria and eukaryotes (Liger et al., 2004; Branco et al., 2008). The arsH gene was firstly identified in Y. enterocolitica (Neyt et al., 1997). This is for the first time resistance against arsenite was developed in L. acidophilus and one of the arsenic resistance gene (arsH) obtained in this bacteria was amplified and sequenced.
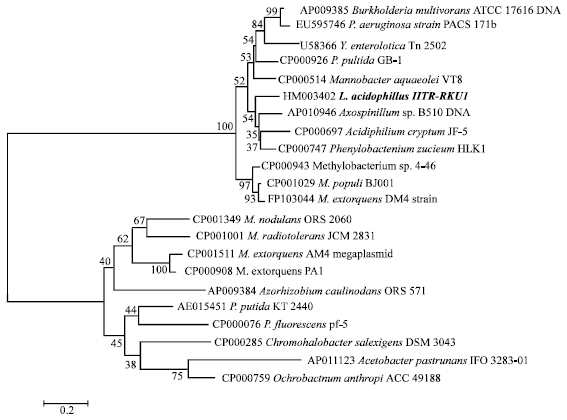 | |
| Fig. 4: | Phylogenetic analysis of amino acid sequences of ArsH of L. acidophilus IITR-RKU1 with ArsH amino acids of other bacteria. Other details are as mentioned earlier |
 | |
| Fig. 5: | Results of conserved domain search. A FMN-reducatse was found within ArsH protein with high scoring hit when analysed using CD server (Marchler-Bauer and Bryant, 2004). * shows stop codon |
Our arsenite resistant L. acidophilus showed entire growth phase pattern similar to its respective wild strain and were found susceptible to various antibiotics (Upreti et al., 2011). The resistance tract of our arsenite-resistant strain IITR-RKU1 was found to be stable. The phenotype was not lost when the strain was sub cultured in a medium without arsenite.
L. acidophilus IITR-RKU1 harbors a 23 kb plasmid. The presence of plasmids in various strains of Lactobacillus is well established. A 70 kb plasmid from Lactobacillus acidophilus C7 and 3.8 and 5.5 kb plasmids of L. acidophilus isolated from fermented dairy products have been reported (Altermann et al., 2005; Osuntoki et al., 2008). Furthermore, Lactobacillus plantarum WCFS1 have been shown to harbor 1, 2 and 36 kb plasmids (Kranenburg et al., 2005). A 606 bp PCR amplicon of our mutant strain was obtained with specific primer of arsH gene which showed its closest similarity with Methylobacterium nodulans, a gram negative bacterium. The resultant 202 bp amino acid sequence showed its nearness with Acetobacter sp. and Burkholderia sp (gram negative). In general, the presence of arsH gene has been reported in gram negative bacterial species (Branco et al., 2008). This is the first report showing the presence of arsH gene in genus Lactobacillus. Alignment of the major portion of conserved domain of this protein with FMN reductase agreed well with the hypothesis that this protein has a reductase function. This fact was further supported by the presence of 12 cysteine residues in one of its reading frames. It has been reported that ArsH protein is an atypical flavodoxin with a non-canonical FMN binding site that catalyzes oxidation of NADPH, generating H2O2 and with a low azoreductase activity (Vorontsov et al., 2007). This gene has also been shown to confer high levels of arsenite resistance and fundamental to arsenic resistance in the four gene operon. Removal of this gene resulted in a reduction of arsenite resistance by E. coli cells in the presence of high levels of As(III) (Branco et al., 2008). The present study further supports that the arsH gene is responsible for the tolerance of high concentrations of arsenite. However, further studies on other genes of ars operon is needed to elucidate the mechanism of arsenical resistance in our strain.
Lactobacilli are one of the predominant species of mammalian gastrointestinal tract and are known to convert toxic forms of metals into their less toxic forms and also help in their detoxification (Shrivastava et al., 2003). It has been suggested that Lactobacillus strains may be propagated as potential probiotics (Anukam and Reid, 2007). Furthermore, L. acidophilus has been considered to be the best known probiotic amongst Lactobacilli (Nguyen et al., 2007). Arsenic-resistant Lactobacilli, which can survive in case of arsenic exposures through drinking water and food, may thus provide fruitful advantages as probiotic in future. Studies on its probiotic efficacy including acid and bile tolerance, intestinal adhesion, persistence on mucosal surface are in progress. In addition, Lactobacilli being a friendly and non-pathogenic group of bacteria, the arsenical resistant L. acidophilus can be a better choice in comparison to other arsenic resistant bacteria for the development of environmental arsenic bioremediation technologies.
CONCLUSION
This is for the first time resistance against arsenite was developed in L. acidophilus and one of the arsenic resistance gene obtained in this bacteria was amplified and sequenced. The arsenite-resistant L. acidophilus strain IITR-RKU1 showed the presence of arsenical resistance gene (arsH) in the plasmid. Further, CD server search revealed that arsH gene of our strain is identical to the FMN reductase.
ACKNOWLEDGMENTS
The authors are grateful to the Director, IITR, Lucknow, for his keen interest in the study. This work was supported by Supra-institutional Project, CSIR (Council of Scientific and Industrial Research), New Delhi and Ad-hoc Project, ICMR (Indian Council of Medical Research), New Delhi, India.
REFERENCES
- Altermann, E., W.M. Russell, M.A. Azcarate-Peril, R. Barrangou and B.L. Buck et al., 2005. Complete genome sequence of the probiotic lactic acid bacterium Lactobacillus acidophilus NCFM. Proc. Natl. Acad. Sci. USA., 102: 3906-3912.
PubMed - Bauer, A.W., W.M.M. Kirby, J.C. Sherris and M. Turck, 1966. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol., 45: 493-496.
CrossRefPubMedDirect Link - Berg, M., H.C. Tran, T.C. Nguyen, H.V. Pham, R. Schertenleib and W. Giger, 2001. Arsenic contamination of groundwater in Vietnam: A human health threat. Environ. Sci. Technol., 35: 2621-2626.
CrossRefDirect Link - Bhakta, J.N., K. Ohnishi, Y. Munekage and K. Iwasaki, 2010. Isolation and probiotic characterization of arsenic-resistant lactic acid bacteria for uptaking arsenic. Int. J. Chem. Bio. Eng., 32: 167-174.
Direct Link - Bose, U., M. Rahman and M. Alamgir, 2010. Arsenic toxicity and speciation analysis in ground water samples. Int. J. Chem. Toxicol., (In Press).
Direct Link - Branco, R., A.P. Chung and P.V. Morais, 2008. Sequencing and expression of two arsenic resistance operons with different functions in the highly arsenic-resistant strain Ochrobactrum tritici SCII24T. BMC Microbiol., 8: 95-95.
CrossRef - Chowdhury, M.A.I., M.T. Uddin, M.F. Ahmad, M.A. Ali and S.M. Uddin, 2006. How does arsenic contamination of ground water causes severity and health hazard in Bangladesh. J. Applied Sci., 6: 1275-1286.
CrossRefDirect Link - Gasteiger, E., A. Gattiker, C. Hoogland, I. Ivanyi, R.D. Appel and A. Bairoch, 2003. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res., 31: 3784-3788.
CrossRefPubMedDirect Link - Kamala-Kannan, S. and K.J. Lee, 2008. Metal tolerance and antibiotic resistance of Bacillus species isolated from Sunchon Bay sediments Korea. Biotechnology, 7: 149-152.
Direct Link - Anukam, K.C. and G. Reid, 2007. Lactobacillus plantarum and Lactobacillus fermentum with probiotic potentials isolated from the vagina of healthy Nigerian women. Res. J. Microbiol., 2: 81-87.
CrossRefDirect Link - Van Kranenburg, R., N. Golic, R. Bongers, R.J. Leer, W.M. de Vos, R.J. Siezen and M. Kleerebezem, 2005. Functional analysis of three plasmids from Lactobacillus plantarum. Applied Environ. Microbiol., 71: 1223-1230.
CrossRefDirect Link - Mahfuzar, R., 2006. International research on arsenic contamination and health. J. Health Population Nutr., 24: 123-128.
Direct Link - Mateos, M., E. Ordonez, M. Letek and J.A. Gil, 2006. Corynebacterium glutamicum as a model bacterium for the bioremediation of arsenic. Int. Microbiol., 9: 207-215.
Direct Link - Mukhopadhyay, R., B.P. Rosen, L.T. Phung and S. Silver, 2002. Microbial arsenic: From geocycles to genes and enzymes. FEMS Microbiol. Rev., 26: 311-325.
CrossRef - Nies, D.H., 1999. Microbial heavy-metal resistance. Applied Microbiol. Biotechnol., 51: 730-750.
PubMed - Nguyen, T.H., B. Splechtna, S. Krasteva, W. Kneifel, K.D. Kulbe, C. Divne and D. Haltrich, 2007. Characterization and molecular cloning of a heterodimeric β-galactosidase from probiotic strain Lactobacillus acidophilus R22. FEMS Microbiol. Lett., 269: 136-144.
Direct Link - Peters, S.C., J.D. Blum, B. Klaue and M.R. Karagas, 1999. Arsenic occurrence in New Hampshire drinking water. Environ. Sci. Technol., 33: 1328-1333.
Direct Link - Salam, M.A., M.S. Hossain, M.E. Ali, M.A. Asad and M.H. Ali, 2009. Isolation and characterization of arsenic resistant bacteria from different environment in south west region of Bangaladesh. Res. J. Environ. Sci., 3: 110-115.
CrossRefDirect Link - Shrivastava, R., R.K. Upreti and U.C. Chaturvedi, 2003. Various cells of the immune system and intestine differ in their capacity to reduce hexavalent chromium. FEMS Immunol. Med. Microbiol., 38: 65-70.
CrossRefDirect Link - Sohel, N., P.L. Ake, M. Rahman, S.P. Kim, Y. Muhammad, E. E. Charlotte and V. marie, 2009. Arsenic in drinking water and adult mortality: A population-based cohort study in rural Bangladesh. Epidemiology, 20: 824-830.
Direct Link - Klaenhammer, T.R., 1984. A general method for plasmid isolation in lactobacilli. Curr. Microbiol., 10: 23-28.
CrossRefDirect Link - Tamura, K., J. Dudley, M. Nei and S. Kumar, 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol., 24: 1596-1599.
CrossRefPubMedDirect Link - Upreti, R.K., A. Kannan and A.B. Pant, 2007. Experimental exposure of arsenic in cultured rat intestinal epithelial cells and cell line: Toxicological consequences. Toxicol. in vitro, 21: 32-40.
CrossRef - Upreti, R.K., A. Kannan and A.B. Pant, 2008. Alterations in rat gut bacteria and intestinal epithelial cells following experimental exposure of antimicrobials. FEMS Immun. Med. Microbiol., 54: 60-69.
CrossRef - Vorontsov, I.I., G. Minasov, J.S. Brunzelle, L. Shuvalova, O. Kiryukhina, F.R. Collart and W.F. Anderson, 2007. Crystal structure of an apo form of Shigella flexneri ArsH protein with an NADPH-dependent FMN reductase activity. Protein Sci., 16: 2483-2490.
CrossRef - Neyt, C., M. Iriarte, V.H. Thi and G.R. Cornelis, 1997. Virulence and arsenic resistance in yersiniae. J. Bacteriol., 179: 612-619.
Direct Link - Liger, D., M. Graille, C.Z. Zhou, N. Leulliot and S. Quevillon-cheruel et al., 2004. Crystal structure and functional characterization of yeast YLRO11wp, an enzyme with NAD(P)H-FMN and ferric ion reductase activities. J. Biol. Chem., 279: 34890-34897.
Direct Link - Osuntoki, A.B., O.R. Ejide and E.A. Omonigbehin, 2008. Antagnonistic effects on entropathogens and plasmid analysis of Lactobacillus isolated from dairy products. Biotechnology, 7: 311-316.
CrossRefDirect Link