
Idress H. Attitalla
Omar Al-Mukhtar University, Faculty of Science, Department of Botany, Box 919, Al-Bayda, Libya
Baharuddin Salleh
Universiti Sains Malaysia, School of Biological Sciences, 11800, Pulau Pinang, Malaysia
Biotechnology
Year: 2010 | Volume: 9 | Issue: 4 | Page No.: 529-532







ABSTRACT
Conidia of two isolates of Trichoderma harzianum (T7 and T8) (Rifai) were formulated to make alginate pellets with or without xylan or CMC as a food-base material. The formulations were compared for their ability of in vitro carboxymethycellulase and xylanase production with free fungal spore suspensions. Conidia entrapped in alginate with or without adjuvant showed high production of enzymes even when repeated 4 times. The addition of adjuvant significantly enhanced the enzyme production. Alginate concentration and surface area of the beads affected the enzyme production. The optimum initial pH and incubation temperature were pH = 5-7 and 35°C for CMC-ase and pH = 5 and 30°C for xylanase. Alginate encapsulated Trichoderma not only prolonged the metabolic activity of the entrapped organism, but also it promotes slow release of microbial spores into the medium for successful enzyme production.
PDF Abstract XML References Citation
Received: June 18, 2010;
Accepted: October 04, 2010;
Published: November 25, 2010
How to cite this article
Idress H. Attitalla and Baharuddin Salleh, 2010. Improvement of Carboxymethyl Cellulase and Xylanase Production by Alginate Immobilized Trichoderma harzianum. Biotechnology, 9: 529-532.
DOI: 10.3923/biotech.2010.529.532
URL: https://scialert.net/abstract/?doi=biotech.2010.529.532
DOI: 10.3923/biotech.2010.529.532
URL: https://scialert.net/abstract/?doi=biotech.2010.529.532
INTRODUCTION
Trichoderma is a fungal genus widely distributed all over the world. In addition to its biological control activities, different species belonging to the genus Trichoderma have been reported to produce cellulases (Chaudhary and Tauro, 1982). Cellulases produced by T. harzianum, is the most efficient enzyme system for the complete hydrolysis of cellulosic substrates into its monomeric glucose, which is a fermentable sugar (Ahmed et al., 2009). Cellulolytic enzyme production is one of the key steps in any process involving enzymatic hydrolysis of lignocellulosics into sugar monomers (Warzywoda et al., 1982). The use of such enzymes to convert wastes for the production of sugars, syrups, alcohol and single cell protein for food and feed has been investigated (Rye and Mandels, 1980). Thus, the necessity to achieve large-scale, cost-effective production of active preparations of Trichoderma has been increased (El-Katatny et al., 2003, 2004).
Immobilization of microbial cells and enzymes has become one of the most valuable tools in the field of biotechnology (EL-Komy, 2005). Moreover, microbial entrapment gives prolonged metabolic activities when microbial cells are reused and protects the organisms from inhibitory compounds or metabolites (Shaban and El-Komy, 2000). Alginate formulations of conidia and mycelia or ascospores of several fungi resulted in rapid fungal increase and proliferation in the soil (Bashan et al., 2002).
The objective of this study was to investigate the in vitro production of CMC-ase and xylanse by alginate encapsulated Trichoderma sp. and to optimize the conditions required for improving the production of these enzymes by the immobilized fungi species.
MATERIALS AND METHODS
Fungal isolates and cultivation condition: Two local isolates of Trichoderma harzianum (T7 and T8) used in this study were isolated from soil samples collected from El Faidia and El Mansora cities, El Bayda, Libya, respectively. Fungal cultures were maintained on Potato Dextrose Agar (PDA) at 4°C.
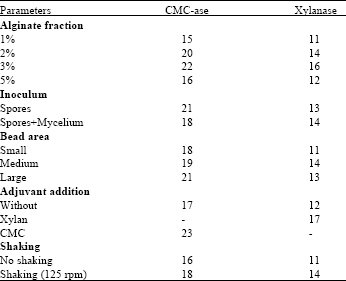
Production of inocula and microencapsulation: Methods used for the production of inocula and microencapsulation were described earlier (Bashan et al., 2002). Microencapsulation was performed using different alginate concentrations: 1, 2, 3, 5%. In some other experiments 0.5% Carboxy Methyl Cellulose (CMC) was added to the alginate spore suspension mixture as adjuvant. Nozzles with different diameters were also used to obtain beads with different surface areas (3, 3.5 and 4 cm3). The fresh beads were either used directly or kept at 4-5°C in sealed flasks for several days. The viable population size of Trichoderma was determined in the pellets before its use in the batch culture fermentation.
CMC-ase and xylanase production by immobilized or free Trichoderma isolates in batch culture fermentation: Batch culture fermentation was carried out in Erlenmyer flasks (50 mL) each containing 10 mL of a medium containing (g L-1): NaNO3, 2; KPO4, 1; MgSO4.7H2O, 0.5; FeSO4.7H2O, 0.001. The appropriate carbon source (0.5% CMC or xylan) was supplied and the pH was adjusted to 5.5 with 50 mM acetate buffer. Flaks were inoculated with either 1 mL of fungal spore suspension or 3 g of fresh beads containing 3x106 CFU per flasks. Flasks were incubated for 7 days at 30°C. After incubation, cultures were separated by filtration for enzyme assays. The effects of initial pH (3-8) and temperature (20-45°C) on the production of enzymes were tested.
The reusability of the immobilized cultures was tested in batch cultures by replacing the culture broth with fresh sterile one every 7 days. Cultivation conditions were as previously described for each set.
Enzyme assays: CMC-ase and xylanase activities were assayed using the viscometric method described by El-Naghy et al. (1991) using CMC and xylan as substrates. Reaction mixture containing 10 mL of 1% substrate dissolved in 0.1 M acetate buffer pH 4.5, 3 mL of 0.1 M acetate buffer pH 4.5 and 2 mL of enzyme preparation and incubated at 30°C for 1 h. Activity was estimated as percentage in reduction of viscosity during specific period of incubation as the following equation:
where, REA is relative enzyme activity; T0 is flow time immediately after the addition of enzyme filtrate and T1 is flow time after incubation and flow time of water.
RESULTS
Results presented in Table 1 showed that Trichoderma harzianum isolate (T7) showed higher CMC-ase and xylanase production than isolate (T8) in both free and immobilized cultures. Therefore it has been selected for further investigations.
The reusability of the immobilized fungus (T7) for enzyme production was studied. Entrapped spores of Trichoderma were successfully used in 4 repetitions for both CMC-ase and xylanase production (Fig. 1).
| Table 1: | CMC-ase and xylanase production* by free and alginate encapsulated Trichoderma isolates |
 | |
| *As percentage of reduction in viscosity | |
| Table 2: | Consecutive improvement of immobilized Trichoderma (T7) for CMC-ase and xylanase production* |
 | |
| *As percentage of reduction in viscosity | |
 | |
| Fig. 1: | Repeated use of immobilized Trichoderma (T7) for CMC-ase and xylanase production |
However, enzymes activities were at the maximum at the 4 th reuse. Optimum alginate concentration for CMC-ase and xylanase was 3%. Enzyme activities increased with the increase of bead surface area (Table 2). Beads entrapping fungal spores showed higher CMC-ase activity, however, beads entrapping spores and mycelium showed higher xylanase activity. The addition of CMC or xylan as adjuvant significantly improved enzyme production.
Data in Fig. 2 showed that CMC-ase had a wide optimum pH range of 5-7, xylanase exhibited its maximum production at pH = 5. The optimum temperatures for CMC-ase and xylanase production by immobilized Trichoderma were 35 and 30°C, respectively (Fig. 3).
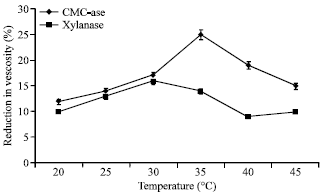 | |
| Fig. 2: | Effect of incubation temperature on CMC-ase and xylanase production by immobilized Trichoderma (T7) |
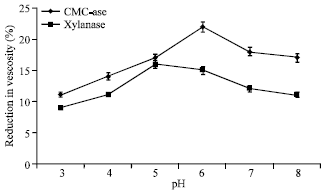 | |
| Fig. 3: | Effect of initial pH of culture medium on CMC-ase and xylanase production by immobilized Trichoderma (T7) |
DISCUSSION
Cellulose is the most abundant renewable organic compound on earth and has received a great attention as a potential substrate for the production of alcohol fuel, chemicals and single cell protein via enzymatic degradation by microbial enzymes (El-Naghy et al., 1991).
Entrapment of microbial cells has been reported to improve the production of proteolytic enzymes (Woodward, 1988). Results showed that immobilized Trichoderma improved CMC-ase and xylanase production compared with free spore suspension especially when CMC and xylan were used as adjuvant. Previous studies indicated that encapsulation technique was further refined by incorporation of nutrient carriers (adjuvant) e.g. wheat bran, milled chitin, corn cobs, soy fibers and peanut hulls into the biopolymers (e.g., alginate) to provide a food base necessarily for proliferation of the entrapped microorganisms (El-Komy, 2001, 2005).
Alginate encapsulation of Trichoderma prolonged the durability of the inoculum and increased in some cases the enzyme production during 4 repetitions. It was observed that the beads became weak and fragile before the last cycle of reuse. This might explain why the enzyme production increased in the last cycle, since the fragile beads allowed the release of more conidia supporting higher growth and enzyme production. The degradation of the pellets has been reported to be due to the presence of certain ions in the medium affecting the stability of the gel (Kennedy and Cabral, 1985).
Optimum alginate concentration for CMC-ase and xylanase production was 2%. It was reported that 3% was the optimal alginate concentration for alkaline proteases produced by Aspergillus flavus ((Woodward, 1988), as well as for the survival of Trichoderma spp. in soil (Lewis et al., 1991). Higher alginate concentration (5%) may reduce microbial growth and enzyme production as a result of limited diffusion of nutrients and oxygen (Van Elsas et al., 1986).
Results of this study showed that pH = 5-7 and pH = 5 were the optimum initial pH values for the production o f CMC-ase and xylanase, respectively. These results are in accordance with previous studies of Ali and Akhand (1992) who reported that pH = 4 was the optimum for cellulase produced by some isolates of Trichoderma. (Gupta et al., 2009) found that xylanase production by Fusarium solani F7 was optimum between pH = 5.0 and 5.5, but the best at pH 5.5 below and above this pH production of xylanase was significantly lower. Moreover, Monti et al. (1991) reported that the pH optimum for Humicola grisea was 5.5 which was close to other microbial xylanases, which were between pH4.0 and 6.0 (Dekker and Richards, 1976).
Data also indicated that optimum temperature for the studied enzymes were 30-35°C. Such results are in agreement with many studies on mesophilic strains of Penicillium spp. and Actinomycetes, which was ranged from 20-40°C (Van Zyl, 1985). Also, Ali and Akhand (1992) reported that maximum cellulase production by Trichoderma isolates was obtained at 28-30°C.
In conclusion alginate immobilization of Trichoderma not only prolonged the metabolic activity of the entrapped organism, but also it promotes slow release of microbial spores into the medium for successful enzyme production. Further studies are needed to clarify the importance of fungal immobilization for the production of such important enzymes in a large scale.
ACKNOWLEDGMENTS
We gratefully acknowledge Professor Lennart Johnsson (MASE laboratory, Uppsala, Sweden), for help with the statistical analyses, helpful comments and discussion; and Dr. Paul Johnson for assistance in interpreting the data, for critical reading and refining of the manuscript; Universiti Sains Malaysia (Malaysia) for inviting me as a scientific visitor and Omar Al-Mukhtar University, Al-Bayda, (Libya) for the financial support.
REFERENCES
- Ahmed, S., A. Bashir, H. Saleem, M. Saadia and A. Jamil, 2009. Production and purification of cellulose-degrading enzymes from a filamentous fungus Trichoderma harzianum. Pak. J. Bot., 41: 1411-1419.
Direct Link - Bashan, Y., J.P. Hernandez, L. Leyva and M. Bacilio, 2002. Alginate microbeads as inoculant carriers for plant growth-promoting bacteria. Biol. Fertil. Soil, 35: 359-368.
CrossRefDirect Link - Dekker, R.F.H. and G.N. Richards, 1976. Hemicellulases: their occurrence, purification, properties and mode of action. Adv. Cabohydr Chem. Biochem., 32: 277-352.
PubMedDirect Link - El-Katatny, M.H., A.M. Hetta, G.M. Shaban and H.M. El-Komy, 2003. Improvement of cell wall degrading enzymes production by alginate encapsulated Trichoderma spp. Food Technol. Biotecnol., 41: 219-225.
Direct Link - El-Katatny, M.S., H.M. El-Komy, G.M. Shaban, A.M.A. Hetta and M.H. El-Katatny, 2004. Effect of benomyl on chitinase and β-1,3-glucanase production by free and alginate encapsulated Trichoderma harzianum. Food Technol. Biotechnol., 42: 83-88.
Direct Link - El-Komy, H.M.A., 2005. Coimmobilization of Azospirillum lipoferum and Bcillus megaterium for successful phosphorus and nitrogen nutrition of wheat plants. Food Technol. Biotechnol., 43: 19-27.
Direct Link - El-Naghy, M.A., M.S. El-Katatny and A.A. Attia, 1991. Degradation of cellulosic materials by Sporotrichum thermophile culture filtrate for sugar production. Int. Biodeterioration, 27: 75-86.
CrossRef - Gupta, V.K., R. Gaur, N. Gautam, P. Kumar, I.J. Yadava and N.S. Darmwal, 2009. Optimization of xylanase production by Fusarium solani F7. Am. J. Food Technol., 4: 20-29.
CrossRefDirect Link - Lewis, J.A., C.C. Papavizas and R.D. Lumsden, 1991. A new formulation system for the application of biocontrol fungi to soil. Biocontrol Sci. Technol., 1: 59-69.
CrossRefDirect Link - Shaban, G.M. and H.M. El-Komy, 2000. Survival and proliferation of alginate encapsulated Trichoderma spp. in Egyptian soil in comparison with allyl alcohol soil fumigation. Mycopathologia, 151: 139-146.
PubMed - Van Zyl, W.H., 1985. A study of the cellulases produced by three mesoophilic actinomycetes grown on bagasse as substrate. Biotechnol. Bioeng., 27: 1367-1373.
CrossRef - Warzywoda, M., J.P. Vandecasleele and J. Pourquie, 1982. A comparison of genetically improved strains of the cellulolytic fungus Trichoderma reesei. Biotechnol. Lett., 5: 243-246.
CrossRef - Woodward, J., 1988. Methods of immobilization of microbial cells. J. Microbiol. Meth., 8: 91-102.
CrossRef - El-Komy, H.M., 2001. Survival of wheat-root colonization by alginate encapsulated Herbaspirllum spp. Folia Microbiol., 46: 25-30.
CrossRef