
M. Keykhahsaber
Sistan Agricultural Biotechnology Institute, University of Zabol, Iran
A.A.V. Sedehi
Sistan Agricultural Biotechnology Institute, University of Zabol, Iran
A. Zakeri
Master of Biology Science, University of Zabol, Iran
H. Khaje
Sistan Agricultural Biotechnology Institute, University of Zabol, Iran
Z. Keykhah
Sistan Agricultural Biotechnology Institute, University of Zabol, Iran
Biotechnology
Year: 2009 | Volume: 8 | Issue: 1 | Page No.: 176-179







ABSTRACT
In order to determine the level of low temperature response genes expression in ten selected barley accessions were obtained from Sistan Agricultural Research at spring of 2008. RNA from each of barley germination was extracted in two cold acclimation and one control condition. The result of real time PCR, using cDNA from three low temperature response gene (bit2, bit14 and bit101), showed that there were significant difference in gene expression between three treatments and in each gene the highest percentage of gene expression belonged to accession 5 while accession 8, 9 and 10 composed one separate. In bit14 gene, the increase in the amount of mRNA was carried out when the maximum level of freezing (4 °C day/2 °C night) apply. All of these genes are shown to be transcriptional regulated and root meristem had maximum level of RNA under cold treatment.
PDF Abstract XML References Citation
How to cite this article
M. Keykhahsaber, A.A.V. Sedehi, A. Zakeri, H. Khaje and Z. Keykhah, 2009. Study of Expression of Low-Temperature-Responsive Genes for Selected Barley Accessions. Biotechnology, 8: 176-179.
DOI: 10.3923/biotech.2009.176.179
URL: https://scialert.net/abstract/?doi=biotech.2009.176.179
DOI: 10.3923/biotech.2009.176.179
URL: https://scialert.net/abstract/?doi=biotech.2009.176.179
INTRODUCTION
In many overwintering temperate plant species a period of low positive temperature will increase tolerance to a subsequent subzero (freezing) temperature. This process is known as cold or frost acclimation and is acknowledged to be complex, involving a number of biochemical and physiological changes. Cellular and metabolic changes that occur during cold acclimation include increased levels of sugars, soluble proteins, proline and organic acids as well as the appearance of new isoforms of proteins and altered lipid membrane composition (Hughes and Dunn, 1990). There is an increasing body of evidence to show that many of these biochemical and physiological changes are regulated by low temperature through changes in gene expression and in recent years a number of Low Temperature-Responsive (LTR) genes have been cloned from a range of both dicotyledon and monocotyledon species (Hughes and Dunn, 1996).
The most extensively studied species is wheat where frost hardiness has been demonstrated to be a multigenic additive trait with major contributions from a few identifiable genes (Doll et al., 1989; Thomashow, 1990). In barley, segregation of frost hardiness and spring growth habit has been studied in doubled haploid progeny of crosses between winter and spring cultivars. Such studies demonstrate that, although the low-temperature response is complex, in cereals a relatively small number of genes are expected to have major effects (Dunn et al., 1994).
In barley, several LTR genes have been described by Hughes and Dunn (1996) and Goddard et al. (1993). Most of the cold-induced barley genes such as bit14, bit101 and bit4 were reported to be small multigene families. bit101 is one of two related genes, which is LT specie and predicted to be a membrane protein. The regulation of bit14 is post-transcriptionally controlled, whereas the expression of bit4.9, the family member of bit4 and bit101 is transcriptionally regulated in barley (Hughes and Dunn, 1996).
Three members of the bit14 gene family have also been cloned from barley and these genes show a pattern of regulation similar to that found in the cognate gene family in rye (Zhang et al., 1993). The genetic analysis of the frost resistance in barley has demonstrated that a QTL for winter survival on barley chromosome 7 (5H) is associated with the Sh2 locus (Hayes et al., 1993). The objective of this study was to asses the relative expression pattern of selected low temperature response, LTR, (bit2, bit14 and bit101) between 10 accessions to understand of the level of expression LTR gene between 10 selected barley accession underlying low temperature acclimation.
MATERIALS AND METHOD
Plant material: This research was conducted in spring of 2008 in Agricultural Biotechnology Institute, University of Zabol. Ten Hordeom vulgaris accessions including landraces and inbred line originated from various region of Iran, obtained from the agricultural research of Zabol, were evaluated in this study (Table 1). Seeds of selected barley (Hordeum vulgare) were germinated in controlled-environment room set at 20/15 °C (day/night), a 10 h photoperiod. This was designated the control environment when the third leaf had emerged in 50 to 75% of plants (about 14 days after sowing). To choose the plant in the same development stage, the vigorous seedling with same height were selected, covered with perlite and grown for approximately 2-3 week at selected conditions. Plants for cold acclimation were transferred to a controlled environment set (growth chamber Wisees g-bh) at 6/2 °C (day/night), 10 h day, 190 μmol m-2 sec-1 photon as negative control one and another part of plant were transferred in growth chamber to for 18 day in 4 °C day/2 °C night, 8 h day and 110 μmol m-2 sec-1 photon as negative control two. Other environmental parameters the same as the control; control plants remained in the 20 °C day/15 °C night environment. Both control and cold-grown plants were analyzed when the fourth leaf was emerging in about 50% of the plants, by which time frost tolerance in the cold-acclimated plants was approaching the maximum for environment. In overwintering grasses and cereals the shoot meristem is at ground level and comprises the meristematic bases of leaves enclosing the stem apex. These parts of the plant are crucial for survival of winter frost and they are the parts of the plants which perceive low temperature for growth acclimation (Hong et al., 1992). The shoot and root meristem was chosen as a source of expressed genes since in Gramineae, it is the part of plant that both perceives low temperature and acclimated for frost tolerance (Thomas and Stoddard, 1984).
| Table 1: | Names and origin of ten accessions of barley used in this study |
 | |
| Table 2: | Sequences of primers used for real-time PCR amplification and the resulting product size |
 | |
RNA extraction and cDNA synthesis: Total RNA from the shoot meristem and the root meristem in low-temperature treatment and control conditions were extracted using RNSy kit according to the manufacturer’s instructions (QIA gene). The total RNA was then quantified on a spectrophotometer (Eppendorf biophotometer). For cDNA synthesis, 1 μg of the DNase-treated RNA was used for first-strand cDNA synthesis using 500 ng of oligodT (12-18), 900 ng of random primer and 200 U of Superscript_III according to the manufacturer’s instructions (Fermentas Revert AidTm first standard cDNA synthesis kit).
Real-time PCR: For real-time PCR, 1 μg of a 1/5 dilution of the cDNA was used as template. The PCR reaction consisted of the respective primers (Table 2), 1xQuantitect_SYBR_Green PCR Master Mix and 30 nM ROX dye in a 25 ll reaction volume (Qiagen, Inc., Mississauga, Ontario, Canada). Real-time PCR was performed in a Roter gene (RG-3000 Corbett Research) under conditions optimized for efficient amplification of the respective genes and reference gene. For bit101 and bit14, amplification was carried out as follows: after the initial activation step at 95 °C for 15 min, 40 cycles each of denaturation at 94 °C for 15 sec, annealing at 60 °C for 1 min and extension at 72 °C for 1 min were performed and for bit2 annealing was carried out at 53 °C for 1 min. For the real-time PCR experiments were conducted on the three biological replicates, with two technical replicates for each sample. Statistical analysis such as compare of pair groups and cluster analysis were analyzed using the Minitab software (Version 14) (Minitab, Inc., Pennsylvania, USA).
RESULT AND DISCUSSION
It has been proposed that the phenotypic adjustment of plants to changes in the environment is result of the expression of different members of multigene families encoding isozymes with different temperature relative properties (Smith, 1990).
Research has been shown that bit2 was induced from 3 h after cold treatment at 4 °C, increased until 24 h after treatment, maximum expression was detected at 24 h and then the level of transcription was slightly reduced at 72 h (Hughs and Dunn, 1996; Goddard et al., 1993).
The level of the expression of bit2 gene was difference between ten accessions; the mRNA accumulated to reach a maximum level within 3 conditions of cold temperature in accession 5 belonged to Semirom while a maximum level reached at the 4 °C day/2 °C night in accession 6 belonged to Esfahan Province. In the accessions 8, 9 and 10 the amount of bit2 expression decrease to a low but steady-state level in bit2 gene expression based on stem RNA. The highest bit2 gene expression was belonged to accession 6 when the temperature on day had hold on 6 to 4 °C. As the other group of charts, there was no significant difference between two negative controls in gene expression while positive control shows a lower gene expression except in accessions 7, 8, 9 and 10. The unpublished research on these accessions had shown that accession 7, 9 and 10 have too much tolerance to cold treatments.
When we used stem meristems cDNA as template in real time PCR reaction, bit2 gene didn’t show difference between three condition including control and two cold treatment (Fig. 1).
In both RNA extraction based of root mersitems of bit101 gene, there were no direction in levels of gene expression for two control and positive treatments while in all accessions negative control show rather level of gene expression (Fig. 2). Same to other LTR gene, in this gene accession 7, 8, 9 and 10 had a lower level of expression. The cluster analysis between 10 accession based on 3 LTR genes show that accession 7, 8, 9 and 10 had closely relationship.
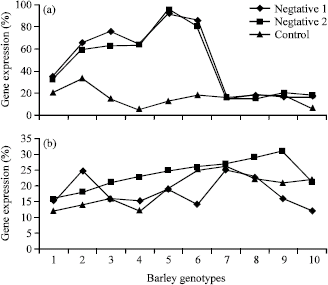 | |
| Fig. 1: | Percentage of bit2 gene expression using mRNA from (a) root meristem and (b) stem meristem of ten barley accession belonged to various region of Iran under three conditions (negative 1: 18 day in 4 °C day/2 °C night, negative 2: 18 day in 4 °C day/2 °C night and control condition 15 day in 20 °C day/15 °C night |
The results of bit14 gene expression for ten accessions of barley show that there was not significance different between two cold treatments (N1 and N2). Accession 6 and accession10 had maximum and minimum expression respectively when root meristem RNA was used for cDNA synthesis. The level of expression of bit101 was monitored closely and compared with the development of freezing tolerance during cold acclimation in barley. The level of freezing tolerance increased after cold treatment (4 °C day/2 °C night) in the all accessions in three groups of bit2 genes. In this study there was no significant difference between tissue-dependent expression in bit14 and bit2 while in bit101, the RNA content under 4 °C day/2 °C night increased when the stem meristem used this result was accorded to another research (Cattivelli and Bartels, 1990; Grossi et al., 1998).
The result of gene expression of bit14 gene, using shoot meristem RNA, was difference from result of percentage of gene expression based on RNA root meristem. In this case the maximum and minimum level of gene expression was belonged to accession 3 and 10, respectively.
Figure 3 shows that in absence of cold treatment bit101 don’t have any difference with cold treatment were applied. Despite of another blt family genes, bit101 didn’t show differences between two cold treatment and two source of mRNA extraction. Accession 7 and 9 belonged to Kerman and Shiraz Province developed RNA expression in bit14 more rapidly than another accessions. A good correlation was observed between the time of cold treatment and development of RNA expression in bit14 and bit2.
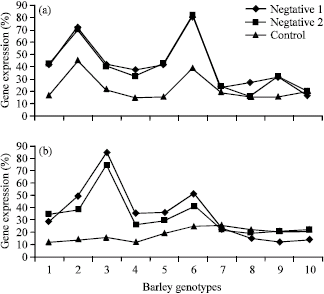 | |
| Fig. 2: | Expression of bit14 gene expression during 3 cold conditions based on (a) root meristem and (b) stem meristem |
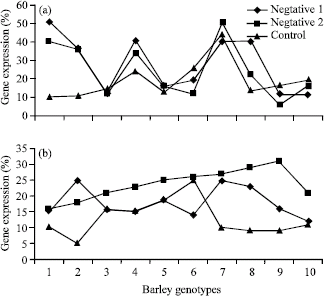 | |
| Fig. 3: | Gene expression in three cold treatment of bit101 gene using (a) root meristem and (b) shoot meristem |
ACKNOWLEDGMENT
This research was funded by Sistan Agricultural Biotechnology Institute, Zabol of University in Iran.
REFERENCES
- Cattivelli, L. and D. Bartels, 1990. Molecular cloning and characterization of cold-regulated genes in barley. Plant Physiol., 93: 1504-1510.
Direct Link - Doll, H., V. Haahr and B. Segaard, 1989. Relationships between vernalization requirement and winter hardiness in doubled haploids of barley. Euphytica, 42: 209-213.
Direct Link - Dunn, M.A., N.J. Goddard, R.S. Pearce and M.A. Hughes, 1994. Low-temperature-responsive barley genes have different control mechanisms. Plant Mol. Biol., 24: 879-888.
Direct Link - Goddard, N.J., M.A. Dunn, L. Zhang, A.J. White, P.L. Jack and M.A. Hughes, 1993. Molecular analysis and spatial expression pattern of a low-temperature-specific barley gene, bit 101. Plant Mol. Biol., 23: 871-879.
Direct Link - Grossi, M., E. Giorni, F. Rizza, A.M. Stanca and L. Cattivelli, 1998. Wild and cultivated barleys show differences in the expression pattern of a cold-regulated gene family under different light and temperature conditions. Plant Mol. Biol., 38: 1061-1069.
Direct Link - Hong, B., R. Barg and T.H.D. Ho, 1992. Developmental and organ-specific expression of an ABA- and stress-induced protein in barley. Plant Mol. Biol., 18: 663-674.
CrossRefDirect Link - Hayes, P.M., T. Blake, T.H.H. Chen, S. Tragoonrung,F. Chen, A. Pan and B. Liu, 1993. Quantitative trait loci on barley (Hordeum vulgare L.) chromosome-7 associated with components of winterhardiness. Genome, 36: 66-71.
Direct Link - Hughes, M.A. and M.A. Dunn, 1990. The effect of temperature on plant growth and development. Biotechnol. Gen. Eng. Rev., 8: 161-188.
Direct Link - Hughes, M.A. and M.A. Dunn, 1996. The molecular biology of plant acclimation to low temperature. J. Exp. Bot., 47: 291-305.
CrossRefDirect Link - Smith, H., 1990. Signal perception differential expression within multigene families and the molecular basis of phenotypic plasticity. Plant Cell Environ., 13: 585-594.
Direct Link - Thomas, H. and J.L. Stoddard, 1984. Kinetics of leaf growth in Lolium temulentum at optimal and chilling temperatures. Ann. Bot., 53: 341-347.
Direct Link - Thomashow, M.F., 1990. Molecular genetics of cold acclimation in higher plants. Adv. Genet., 28: 99-131.
Direct Link - Zhang, L., M.A. Dunn, R.S. Pearce and M.A. Hughes, 1993. Analysis of organ specificity of a low-temperature-responsive gene family in rye (Secale cereale L). J. Exp. Bot., 44: 1787-1793.
Direct Link