
S. Shahrokhi
College of Agriculture Science, Bahonar University of Kerman, Iran
G.H. Shahidi Bonjar
College of Agriculture Science, Bahonar University of Kerman, Iran
I. Saadoun
Jordan University of Science And Technology, Irbid, Jordan
Biotechnology
Year: 2005 | Volume: 4 | Issue: 2 | Page No.: 132-138







ABSTRACT
This is the first report of antifungal activity of Iranian actinomycete isolates against Rhizoctonia solani Kuhn AG-3 (Teleomorph: Thanatephorus cucumeris [(Frank) Donk]). Biological control offers an environmentally friendly alternative to the use of antimicrobials for controlling plant diseases. A collection of about 200 actinomycete strains was screened for the ability to produce metabolites that inhibit R. solani growth in vitro. The Streptomyces olivaceus strain 115 showed strong in vitro antagonistic activity against R. solani in agar disc and Well-diffusion methods by producing extracellular antifungal metabolites. The strain No. 115 was propagated in submerged cultures and active crude was prepared upon which some biological characterization performed. The active metabolite(s) is polar, soluble in H2O and methanol but insoluble in chloroform, dichloromethane or hexane. Thermal inactivation point of active phase of S. olivaceus strain 115 was 80°C. Antifungal active phase of S. olivaceus strain 115 tolerate range of pH (6-9). Antifungal gene from strain 115 may be a useful candidate for genetic engineering of agriculturally important crop for increased tolerance against R. solani.
PDF Abstract XML References
How to cite this article
S. Shahrokhi, G.H. Shahidi Bonjar and I. Saadoun, 2005. Biological Control of Potato Isolate of Rhizoctonia solani by Streptomyces
olivaceus Strain 115. Biotechnology, 4: 132-138.
DOI: 10.3923/biotech.2005.132.138
URL: https://scialert.net/abstract/?doi=biotech.2005.132.138
DOI: 10.3923/biotech.2005.132.138
URL: https://scialert.net/abstract/?doi=biotech.2005.132.138
INTRODUCTION
considerableattentionm cmentyears. Several microorganisms antagonistic to R. solani have been Continued problems antimicrobials, including associated with chemical fnngicides, have stimulated studied for biocontrol of this pathogen including fnngi as some Trichoderma spp., Gliocladium spp., interest in biocontrol strategies and the subsequent identification and characterization of antagonists useful for biocontrol purposes[1]. Advances in the search of biological control of soilborne plant pathogens are accelerating at rapid rate. This phenomenon is partly due to increased knowledge in the production, formulation and delivery of biocontrol agents, which include fnngi, bacteria and actinomycetes[2,3]. The Rhizoctonia disease complex of potatoes comprises two distinct phases: infection of growing plants (Rhizoctonia canker) and infestation of daughter tubers by sclerotia (black scurf). Both are economically important and each has been the objective of much researches. The Rhizoctonia disease is present everywhere in the world that potatoes are grown. The Soilborne pathogenR. solani is ubiquitous and nnder appropriate environmental conditions, can damage the plants severely[4-6].. The application of biocontrol agents to suppress disease caused by R. solani has received Verticillium biguttatum and bacteria as Bacillus spp., Pseudomonas fluorescens and actinomycetes (especially Streptomyces spp)[7-11]. Actinomycetes represent a high proportion of the soil microbial biomass. They have the capacity to produce a wide variety of extracellular hydrolases that give them an important role in the decomposition of organic matter in the soil. They appear to have a high degree of importance among the microbial flora of the rhizosphere. Actinomycetous bacteria have been recognized as sources for several secondary metabolites, antibiotics and lytic enzymes of medical and industrial value but only a few taxa mainly Streptomyces spp. have been studied as potential biocontrol agents against fnngal phytopathogens[12-15].. Several biocontrol agents are commercially available. Mycostop which contains living mycelium and spores of S. griseoviridis is applied as a seed dressing or by soil treatment for control of a number of soilborne plant pathogenic ftmgi as Pythium spp., Fusarium spp. and R. solani, etc.[16].. A group of antibiotics named Validarnycins is produced by S. hygroscopicus. A commertial preparation of Validarnycin A has been used to control disease incited by R. solani[17].This material is used in Japan and the Netherlands to control of black scurf of potato cause by R. solanzil1l. Streptomyces sp. Di944 has bee reported to suppress damping-off of tomato transplants caused by R. solani Kulm rmder controlled envirornnental conditions[18]. Notably, Rothroch and Gottlieb[19] reported direct evidence which showed that the control of Rhizoctonia root rot in pea plants by S. hygroscopicus var. geldanus in an artificially infested sterile soil depended upon the in situ concentration of Geldanamycin (=20 ) μg g-1 of soil) an antibiotic produced by this strain in the soil.
Present investigation was initiated to exploit the ability of actinomycetes to inhibit saprophytic growth of R. solani in vitro. In the present research, 200 isolates of Streptomyces spp. were isolated from agricultural soils of Kerman and Hormozgan provinces, Iran and screened against Rhizoctonia solani through which strain 115 was the most active demonstrated by in vitro studies. Preliminary characterization and biological properties of this strain is being reported here.
MATERIALS AND METHODS
Culture media: A synthetic medium, Casein glycerol (or starch) agar (CGA) was used for screening and isolating of Streptomyces which composed of glycerol or soluble starch, 10 g; casein, 0.3 g; KNO3, 2 g; NaCl, 2 g; K2HPO4, 2 g; MgS04.7H2O, 0.05 g; CaCO3, 0.02 g; FeS04.7H20, 0.01 g and agar, 18 gin 1 L of distilled H2O (pH 7 .2)[20] In submerged cultures, agar was excluded (CG medium). Strptomyces colonies with different morphologies were selected and transferred to CGA slants for further studies[21,22]. R. solani isolate was grown at 20°C and maintained on Potato Dextrose Agar (FDA) (Difco).
Preparation of R solani isolate: A pure culture of potato isolate of R. soloni AG-3 was kindly supplied by Prof Bainhashemi, Mycology Lab., Dept of Plant Pathology, College of Agriculture, Shiraz, Iran. The ftmgus was propagated on FDA and subcultured as needed.
Soil sampling and isolation of Streptomyces: Soil samples were collected from grasslands, orchards and vegetable fields in different localities of Kerman and Hormozgan provinces, Iran. Several samples randomly were selected from mentioned localities using an open-end soil borer (20 em in depth, 2.5 em in diameter) as described by Lee andHwang[23]. Soil samples were taken from a depth of 10-20 em below the soil surface. The soil of the top region (1 0 ern from the surface) was excluded. Samples were air-dried at room temperature for 7-10 days and then passed through a 0.8 mm mesh sieve and were preserved in polyethylene bags at room temperature before use. Samples (10 g) of air-dried soil were mixed with sterile distilled water (100 mL ). The mixtures were shaken vigorously for 1 h and then allowed to settle for 1 h. Portions (1 mL) of soil suspensions (diluted 10-1) were transferred to 9 mL of sterile distilled water and subsequently diluted to 10-2', 10-3, 10-4, 10-5 and 10-6. Inocula consisted of adding aliquots of 10-3 to 10-6 soil dilutions to autoclaved CGA (1 mL-25 ML CGA) at 50°C before pouring the plates and solidification. Three replicates were considered for each dilution. Plates were incubated at 30°C for up to 20 days. From day 7 on, Streptomyces colonies were isolated on CGA, incubated at 28°C for one week and stored refrigerated as pure cultures before use. For screening studies 200 pure Streptomyces isolates were collected.
Screening procedures and in vitro antifungal bioassays agar disk-method: Each Streptomyces isolate was smeared on CGA medium as a single streak and after incubation at 28°C for 4-6 days, from well-grown streaks 6 mm agar disks of Streptomyces colony mass were prepared by using sterile cork borers. Disks were then aseptically transferred to FDA plates having fresh lawn culture ofR. so/ani isolates. Controls included using plain disks from CGA medium. Plates were incubated at 20°C for 4-6 days and bioactivity was evaluated by measuring the diameter of inhibition zones (DIZ, mm)[20-24].
Dual culture bioassay: Ftmgal mycelial-disks (diameter of 6mm) prepared from growing margin of cultures of test R. solani isolate and placed in the center of FDA plates and at 30 mm distance from it, the Streptomyces disks (prepared as mentioned) were placed. Plates incubated at 20°C for 4-6 days[20]. Antifnngal activity was indicative as mycelial grovvth of R. solani isolate was prohibited in the direction of active Streptomyces isolate. The level of inhibition at dual cultures was calculated by subtracting the distance (mm) of fnngal grovvth in the direction of an antagonist colony (Γ) from the fungal growth radius (Γ°) of a control culture to give Δ Γ= Γ°-Γ.The ratings used were modified from those of Lee et al.[23]and El-Tarabily et al[25] where,Δ Γ: 5-9 mm, +(weak inhibition); Δ Γ:10-19 mm, ++(moderate inhibition) and Δ Γ > 20 rnrn, +++(strong inhibition). Controls included R. solani mycelial plugs in center of non-Streptomyces inoculated FDA plates.
Well diffusion method: For evaluation of antiftmgal activity of aqueous samples, by use of sterile cork borer wells (6x4 mm, 2 em apart) were prmctured in fresh la\Vll cultures or at 30 mm distance from plugs of R. solani isolate. Respective concentrations in dimethyl sulfoxide: methanol (1/1: v/v) solvent (DM solvent) were then administered to fullness in each well. Plates were incubated at 20°C for 4-6 days for lawn cultures and dual culture disk-plugs. Bioactivity was determined by measuring inhibitory zones (mm). Each experiment was repeated three times and the mean of inhibitory zones recorded. Controls included use of blank wells and use of DM solvent without test compmmds[20].
Submerged cultures and preparation of crude extract: Strain 115, the most active among other isolated Streptomyces strains, was grownl in submerged cultures ofCG medium on rotary shakers nnder 130rpm at30°C. To monitor the activity versus post seeding time, aseptically small aliquots of culture media were taken every 24 h for 20 days and the activity was evaluated by well diffusion� method[20-24]. To prepare crude extract, after 6-7 days of post seeding which the activity reached its maximum, the cultures were harvested; spores and mycelia were excluded by filtration through two layers of cheese cloth. The clarified sap was then dried to dark crude nnder reduced air at 50°C, pulverized and kept refrigerated before use.
Classification of Streptomyces strain 115: Streptomyces colonies were characterized morphologically and physiologically following the direction mentioned in the methods manual of international cooperative project for description and deposition of cultures of Streptomyces (ISP)[26]
Morphological characterization: Streptomyces colonies on glycerol-nitrate-casein agar were transferred onto oatmeal agar and streaked across the plate and incubated in the dark at 27°C for 21 days.
Color determination: This made for: a) Mass color or mature, sporulating aerial surface grovvth, b) The color of substrate mycelium as viewed from the reverse side and c) Diffusible soluble pigments other than melanin. Observation was made after 21 days and was limited to mature cultures with heavy spore mass surface using code for determining the color of aerial mycelium of Streptomycetes composed by Prauser[27] for color tabs of Baumann Farbtonkarte Atlas.
Determination of morphological characteristic of the spore bearing hyphae: The spore-bearing hyphae characteristics were determined by direct microscopic examination of the culture surface (21 days old) on opened dishes of the crosshatched cultures using 100 x magnification. The species involved in the genus Streptomyces divide into sections: Rectus (R) or straight, flexible (F) or flexeous, Retinaculurn-Aperturn (RA) and spiral (S).
Melanin production: Peptone iron agar was used for the detection of deep brown to black diffusible pigment (+). Absence of the color was recorded as negative(-).
Carbon utilization: The following sugars were tested, L-arabinose, D-xylose, meso-inositol, D-mannitol, D-fructose, rhamnose, raffinose and sucrose. Preparation was done as described in the ISP[26l. Characterization of Streptomyces strain 115 to species level was based on morphological, cultural and physiological characteristics following the directions giVen for the International Streptomyces project (ISP)[26]. General morphology was determined on oatmeal agar plates, incubated in the dark at 27° C for 21 days, by direct light microscopy examination of the surface of crosshatched cultures. Colors were determined according to the scale adopted by Prauser[27] and melanin reactions were detected by growing the isolate on at least one of the ISP media (No. 6 and 7). Strain 115 was identified as a new strain of Streptomyces olivaceus.
Determination of Minimum Inhibitory Concentrations (MIC): To measure the MIC values, two-fold serial dilutions of20, 10, 5, 2.5, 1.25, 0.625 and 0.312mg rnL-1,of the crude extract were prepared in DM solvent and assayed by well diffusion-method as described by Shahidi Bonjar[28]. The MIC was defined as the lowest concentration able to inhibit any visible fnngal grovvth. All data represent average of three replicated experiments.
Solubility studies of active crude in organic solvents: To evaluate the relative polarity of the active principle (s) present in the crude, 2 mL of each of H20, methanol, DMSO: Methanol (1:1, v/v), chloroform, dichloromethane and hexane were added to 20 mg pulverized-crude samples separately and vortexed for 20 min. Each sample was then centrifuged at 3000 rpm for 15 min. Supernatants and pellets were separated, dried nnder reduced air at 50°C and assayed at concentration of 1 0 mg mL-1 by agar diffusion-method[20].
Determination of shelf life or stability of active crude: To measure the stability of the active crude in both soluble
and dry states, 5 mg mL-1 of each sample was prepared in DM solvent and 5 mg dry samples placed in small vials. These samples were kept at room temperature and tested using agar diffusion-method for anti Rhizoctonia activity at 14 days intervals as long as the activity persisted.
Effect of heat on bioactivity: To monitor the effect of temperature on bioactivity, small aliquots (10 mg mL-1) of soluble crude were exposed to each of 30, 40, 50, 60, 70, 80 and 90°C for 10 min and cooled on ice afterwards[29]. Bioactivity of treated samples was evaluated using well diffusion method. Control included incubation of an nntreated sample at 26°C. All samples were tested by well diffusion method as described earlier.
Effect of pH on antifungal bioactivity: Effect of pH on activity and stability of activity was measured at different pH values by the general standard assay methods. The pH of the reaction mixtures was varied using the buffers described by Covington and Davison[29,30]. The pH stability of the active crude sap was evaluated by incubating it for 0.5 h at various pH values at 30°C and evaluating them by agar well diffusion method.
RESULTS
Screening and bioassays: In screening for metabolites of soil Streptomyces spp. having antifnngal activity against isolate of the cosmopolitan pathogen, Rhizoctonia solani Kulm, 200 isolates were screened from which strain 115 showed high level of activity.
Determination of C: In well diffusion-method, MIC of the crude was determined as 2.5 mg mL-1 against R. solani.
Solubility of active crude in organic solvents: Solubility results show, apparently the active principle(s) has a polar nature since activity is recoverable only in H20, methanol supernatants and pellets of chloroform, dichloromethanev and hexane treatments (Table 1).
Shelf life or stability of active crude: Stability of the active crude in DM solvent and dry form determined one month at 15°C and three months at 25°C, respectively, assayed by us1ng agar diffusion-method against R. solani.
Antifungal activity of submerged cultures: Activity versus post seeding time in submerged media cultures is indicated in Fig. 1. Since the activity reaches its maximum after 6-7 of post seeding, this time was used to harvest cultures for preparation of crude extract.
| Table 1: | Bioassay results of solubility tests of the antifungal principle(s) of Streptomyces olivaceus strain 115 against R so/ani in fractions of different solvents indicated by well diffusion-method at 10 mg mL-1 of dry crude |
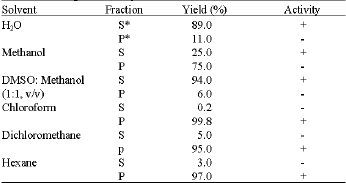 | |
| S*: supernatant, P*: pellet | |
`
| Table 2: | M01phological and physiological characterization of Streptomyces olivaceus strain 115 |
 | |
| S: Spiral; Gy; Sm: Smooth; I: Positive, 0: Negative; -:No utilization; +:Utilization;± Not clear | |
 | |
| Fig. 1: | Activity versus post seeding time in submerged media cultures of Streptomyces olivaceus strain 115 againstR. solani |
Inhibition of mycelial growth and sclerotia formation: As revealed by microscopic examinations, antagonistic activity of S. olivaceus strain 115 against R. solani consists of two inhibitory effects.
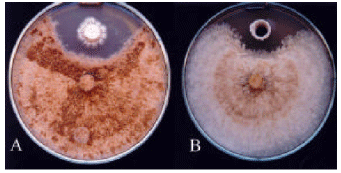 | |
| Fig. 2: | Antagonistic activity of S. olivaceus strain 115 against Rhizoctonia solani revealed by agar disk (A) and well diffusion methods (B). Antifungal activity consists of two inhibitory effects, clear zones adjacent to the plug or well, represent complete mycelial growth (fungicidal activity) and sclerotial inhibition zone in the periphery |
As indicated in Fig. 2, clear zones adjacent to the wells represent complete Antagonistic activity of S. olivaceus strain 115 against Rhizoctonia solani revealed by agar disk (A) and well diffusion methods (B). Antifungal activity consists of two inhibitory effects, clear zones adjacent to the plug or well, represent complete mycelial growth (fungicidal activity) and sclerotial inhibition zone in the periphery mycelial growth (fungicidal activity) and in their periphery, there is sclerotia inhibition zone.
Taxonomy of Streptomyces strain 115: Strain 115 was identified as streptomyces olivaceus strain 115 which is a new record from Iran. Table 2 shows the complete identification of this isolate based on morphological and biochemical characterization.
Effect of heat on activity: Up to 80°C, temperature had no effect on antifungal activity of S. olivaceus. Effect of pH on activity: Effect of pH on antifungal bioactivity of 7-days old aqueous submerged cultures of S. olivaceus strain 115 revealed that the activity was stable at 6-9 pH but drops beyond this range.
DISCUSSION
R. solani Kuhn (releomorph: Thanatephorus cucumeris [(Frank) Donk]) is an important pathogen causing dramatic yield losses worldwide on potato. The universal phase-out of the broad-spectrum fumigant methyl bromide as a control measure for soilborne plant pathogens, alternative control methods for R. solani are urgently needed for commercial potato production. Potential uses of actinomycetes as replacements or supplements for agricultural chemical fungicides have been addressed in many reports[31,32] Significant yield losses due to fungal attacks occur in most of the agricultural species. Genetic engineering has been successful in protecting some of the major crops grown around the world from fungal diseases. Genes encoding many antifungal proteins which can inhibit fungal growth have been exploited to make fungus-resistant transgenic plants[33,37]. The main objective in our study was to identify biologically active streptomyces spp. against R. solani. Present results showed presence of potential antifungal metabolites in S. olivaceous strain 115 against R. solani. Results of these findings may evoke the research for recombinant DNA having antifungal genes cloned from biologically active streptomyces spp. Expression of cloned gene in transgenic plants has provided evidence in plant defense. Thus it may be assumed that the antifungal-metabolite gene from S. olivaceus strain 115 may be a useful candidate for genetic engineering for development of the desired resistant potato cultivars.
ACKNOWLEDGMENTS
Thanks to Head of the Department of Graduate Studies and Research Affairs Office of Bahonar University of Kerman for financial support ofthe project. Helpful information and kind gift of Rhizoctonia solani Kuhn AG-3 isolate by Prof. Banihashemi, Department of Plant Protection, College of Agriculture, Shiraz, Iran is appreciated. This research is dedicated to Mr. A. Afzalipour, the founder of Bahonar University in Kerman.
REFERENCES
- Tsror, L., R. Barak and B. Sneh, 2001. Biological control of black scurf on potato under organic management. Crop Prot., 20: 145-150.
CrossRefDirect Link - Yu, G.Y., J.B. Sinctair, G.L. Hartman and B.L. Bertagnolli, 2002. Production of iturin A by Bacillus amyloliquefaciens suppressing Rhizoctonia solani. Soil Biol. Biochem., 34: 955-963.
CrossRefDirect Link - Kondoh, M., M. Hirai and M. Shoda, 2001. Integrated biological and chemical control of damping-off caused by Rhizoctonia solani using Bacillus subtilis RB14-C and flutolanil. J. Biosci. Bioengineer., 2: 173-177.
PubMedDirect Link - Papavizas, G.C., 1985. Trichoderma and Gliocladium: Biology, ecology and potential for biocontrol. Annu. Rev. Phytopathol., 23: 23-54.
CrossRefDirect Link - Valois, D., K. Fayad, T. Barasubiye, M. Garon, C. Dery, R. Brzezinski and C. Beaulieu, 1996. Glucanolytic actinomycetes antagonistic to Phytophthora fragariae var. rubi, the causal agent of raspberry root rot. Applied Environ Microbiol., 62: 1630-1635.
Direct Link - Saadoun, I. and R. Gharaibeh, 2002. The Streptomycetes flora of Jordan and its potential as a source of antibiotics active against antibiotic resistant Gram negative bacteria. World J. Microbiol. Biotechnol., 18: 465-470.
Direct Link - Saadoun, I., F. Al-Momani, H. Malkawi and M.J. Mohammad, 1999. Isolation, identification and analysis of antibacterial activity of soil streptomycetes isolates from North Jordan. Microbios, 100: 41-46.
Direct Link - Lee, J.Y. and B.K. Hwang, 2002. Diversity of antifungal actinomycetes in various vegetative soils of Korea. Can. J. Microbiol., 48: 407-417.
Direct Link - El-Tarabily, K.A., M.H. Soliman, A.H. Nassar, H.A. Al-Hassani, K. Sivasithamparam, F. McKenna and G.E. Hardy, 2000. Biological control of Sclerotinia minor using a chitinolytic bacterium and actinomycetes. Plant Pathol., 49: 573-583.
Direct Link - Shirling, E.B. and D. Gottlieb, 1966. Methods for characterization of Streptomyces species. Int. J. Syst. Evol. Microbiol., 16: 313-340.
CrossRefDirect Link - Bonjar, G.H.S., 2004. New approaches in screening for antibacterials in plants. Asian J. Plant Sci., 3: 55-60.
CrossRefDirect Link - Fokunang, C.N., J.L. Beynon, K.A. Watson, N.H. Battey, J.M. Dunwell and E.A. Tembe-Fokunang, 2004. Advancement in genetic modification technologies towards disease resistance and food crop production. Biotechnology, 3: 1-20.
CrossRefDirect Link - Aghighi, S., G.H.S. Bonjar and I. Saadoun, 2004. First report of antifungal properties of a new strain of Streptomyces plicatus (Strain 101) against four Iranian phytopathogenic isolates of Verticillium dahliae, a new horizon in biocontrol agents. Biotechnology, 3: 90-97.
CrossRefDirect Link - Bonjar, G.H.S. and A. Sonia, 2005. Chitinolytic and microsclerostatic activity of iranian strains of Streptomyces plicatus and Frankia sp. on olive isolate of Verticillium dahliae. Biotechnology, 4: 108-113.
CrossRefDirect Link - Zamanian, S., G.H. Shahidi Bonjar and I. Saadoun, 2005. First report of antibacterial properties of a new strain of Streptomyces plicatus (strain 101) against Erwinia carotovora from Iran. Biotechnology, 4: 114-120.
CrossRefDirect Link - Nawani, N.N. and B.P. Kapadnis, 2004. Production dynamics and characterization of the chitinolytic system of the Streptomyces sp. NK1057, a well equipped chitin degrader. World J. Microbiol. Biotechnol., 20: 487-494.
CrossRefDirect Link - Berg, G., A. Krechel, M. Ditz, R.A. Sikora, A. Ulrich and J. Hallmann, 2005. Endophytic and ectophytic potato-associated bacterial communities differ in structure and antagonistic function against plant pathogenic fungi. FEMS Microbiol. Ecol., 51: 215-229.
PubMedDirect Link