
Naser Arabzadeh
Assistant Professor of Agricultural and Natural Resources Researches Center, Sodoughi Avenue, P.O. Box 76175-538, Kerman, Iran
Asian Journal of Plant Sciences
Year: 2012 | Volume: 11 | Issue: 1 | Page No.: 28-35







ABSTRACT
Osmotic and elastic parameters of Haloxylon aphyllum branchlets after inducing dryness were studied. The study of changes of water relations parameters with the aim of improving resistance to drought of this species through inducing chronic dryness and clarification of physiologic mechanisms of this plant in response to a low water and dryness are among the objectives of this study. For this purpose, the method of pressure chamber was employed. By this method, the pressure-volume curves were drawn and through their analysis, plant water relations parameters were obtained. A relatively mild dryness was induced to twigs through a lack of irrigation. It was such that after two weeks, the potential of the water of the branchlet of H. aphyllum twigs reached to -16.5 bars. A very intensive dryness was also induced to that but after four weeks of prevention from irrigation; it was such that the potential of the water of H. aphyllum branchlet was diminished to -27.2. A relatively mild dryness was repeated for six terms and a relatively harsh dryness for eleven consecutive periods. In both series of experiment, the potential of water of control twigs were being watered every two days once, remained fixed at about -12.7 bars. Based on the results, though the relatively mild dryness induction to twigs of this type of Haloxylon, made the increase of elastic property of branchlet textures but it had not a meaningful impact on its osmotic potential. Although the application of a relatively intensive dryness reduced the osmotic potential of H. aphyllum branchlets but at the same time, it increased the elasticity of the tissues too.
PDF Abstract XML References Citation
Received: November 14, 2011;
Accepted: January 17, 2012;
Published: February 16, 2012
How to cite this article
Naser Arabzadeh, 2012. Physiologic Responses of Haloxylon aphyllum to Consecutive Tensions of Dryness and Study of Their Role in Improving Resistance to Dryness of Vase Twigs. Asian Journal of Plant Sciences, 11: 28-35.
DOI: 10.3923/ajps.2012.28.35
URL: https://scialert.net/abstract/?doi=ajps.2012.28.35
DOI: 10.3923/ajps.2012.28.35
URL: https://scialert.net/abstract/?doi=ajps.2012.28.35
INTRODUCTION
Haloxylon shrubs as resistant plants are scattered in different growth areas from more or less sporadic masses in natural form in some of the desert and arid zones of Iran (Arabzadeh et al., 2009).
Plants show a wide range of adaptations, at different levels, to drought stress (Xoconostle-Cazares et al., 2010). Reproductive growth of plants is exposed to low rainfall and drought stress in vegetative parts of Iran (Danesh-Shahraki et al., 2008). There are few physiological processes in plants that are not impaired by water deficit (Kumari, 2010) but, stress management is able to more resistant the plants such as Haloxylon against the negative effects of stress (Arabzadeh, 2009). Improving drought tolerance is important for breeding (Khalily et al., 2010) and vigority (Arabzadeh et al., 2009).
Several environmental factors adversely affect plant growth. Drought stress is one of the major factors in limiting production (Omidi, 2010) and growth (Arabzadeh et al., 2009) particularly in arid and semi-arid locations. In such regions, germination and seedling establishment constitute are the most critical periods in the life-cycle of plants (Omidi, 2010).
Drought stress causes a wide variety of physiological and biochemical changes in plants (Omidi, 2010). Drought is one of the most important factors in the ecological field which many crops such as corn (Ngaboyisonga et al., 2009) and other crops such as Haloxylon (Arabzadeh et al., 1992) be able to interact with it.
Drought is the most serious threat to agriculture and is far more important globally (Ali, 2011). Growth of plants such as wheat and others is reduced depending upon degree of water stress and on the rate of stress development, thereby limiting final product (Ali, 2011).
Water stress is a major harmful factor in arid and semi-arid regions worldwide (Ali, 2011). In these regions, it is one of the severe limitations of crop growth as it has a vital role in plant growth and development at all growth stages (Gorji et al., 2011). Drought stress is an important factor in environmental stresses range which limits natural sources and agriculture productions (Imanparast and Hassanpanah, 2009) and controls the plant growth directly (Kufa and Burkhardt, 2011). It is responsible for production losses and is a major factor that limits the growth and reproduction of plants (Naderikharaji et al., 2008).
Plants absorb maximum water from the soil and minimal water loss from their leaves to cope with drought stress (Rad et al., 2011). For drought resistance, several morphological, physiological and biochemical mechanisms are used (Rad et al., 2011). Drought stress influences virtually all physiological functions that are responsible for plants growth (Kufa and Burkhardt, 2011). Haloxylon plants strategy to deal with drought stress and its negative effects is the production of protective substances and increase the elasticity of the tissues (Arabzadeh, 2012). Survival of these plants under drought stress is possible using osmotic adjustment too (Arabzadeh, 2011).
The term resistance to dryness indicates the physiologic and biochemical features of plant which in the condition of limitation of the soil wetness makes it viable. Plants use two mechanisms of bearing drought or escaping from it in confronting with drought (Turner, 1979; May and Milthorpe, 1962). H. aphyllum is among species which with the help of tolerance mechanism is able to spend drought periods. This mechanism has been studied in some of the species like various kinds of pine (Emadian, 1988; Grime, 1979; Bilan et al., 1978, 1977; Youngman, 1965).
The Haloxylon species like many other species, by adjusting the Osmosis or increasing the elastic property of the cellular wall of their own textures in the conditions of water stress, maintain their Turgescence better and consequently bear drought periods better (Emadian, 1988).
H. aphyllum like other multi-functional plants of Haloxylon genus is of great value and importance in protecting and supporting breakable ecosystems like desert. It is such that as a live windbreaker it prevents from erosion of soil (Tokasi et al., 2007; Safarnejad, 2005; Jafari et al., 2004) and as an improving element increases the organic materials of soil and in a long term, it improves the structure of soil (Jafari et al., 2004) and increases the plant enrichment of the area under coverage (Bakhshi and Biroudian, 2008). In addition for those who live in desert, it is an important source for the provision of fuel and fodder for cattle (Tokasi et al., 2007).
However, unfortunately, despite the high importance of Haloxylon in biologic stabilization of sand lands, most of the planted twigs in desert scenes are facing the problem of dryness. Based on the confirmation of authorities and those in charge of desertification projects, after transferring the twigs of H. aphyllum planted in vase to a natural plantation bed and despite observing principles related with post-plantation stages, after sometimes, a high percentage of transferred twigs were afflicted with dryness and they had to be replanted (Arabzadeh et al., 1992; Kerman Natural Resources Office, 1990). Taking action to replant the dried twigs imposes relatively extravagant costs to the executive system of natural resources. According to the existing documents in the File Keeping Department of Ministry of Agriculture of Iran, in average in each period, more than 10% of the vases with H. aphyllum into the scene were fading and became dried and were in need of re-plantation (Arabzadeh et al., 1992). In order this action and aiming at reducing the percentage of mortality of vase based planted twigs in main beds of plantation, it seemed that placing a one-year old twig of H. aphyllum which are exposed to periodical dryness and possibility of their compatibility with unfavorable conditions resulting from reduction of rainfall and periodical droughts can be tested as an appropriate approach. The main motivation in conducting this research was firstly to study the changes of parameters of water relation in order to improve resistance to the dryness of H. aphyllum through induction of periodical dryness and also clarification of physiological mechanisms of this species in response to water shortage and dryness.
MATERIALS AND METHODS
Production and location of twigs: The identified seeds of H. aphyllum were planted in plastic vases and they were taken into care for one year. Plastic vases with an approximate capacity of three liters were selected in order to pave way for a better and a greater growth of the twigs roots. Their soil consisted of wind sand, soil and leaf-soil in proportions of 2, 1 and 1, respectively. By including leaf-soil in the mentioned combination, the aim was to supply necessary nutrition for twigs during the experimental period. At the same time, in order to prevent from unwanted accumulation of water in vases, some fine holes were made in their bottom. After one year, the twigs were transferred to a greenhouse.
Dryness induction: Upon completion of a one-month period of compatibility of twigs in greenhouse, the treatment of dryness induction was applied. For this purpose, a sufficient number of good and healthy twigs were selected for the experiment. Half of them were considered for induction of dryness and the rest as the control and they were being watered every two days once. In this study, two series of experiments were conducted. The twigs of the experiment of the first series received 6 periods of 7-14 days of dryness. During this period, the water potential (before sunrise) of their branchlet was measured every two days once by using the set of pressure chamber and according to Scholander method (Scholander et al., 1965). At this state, the water potential of Haloxylon branchlets at the end of each period diminished in average -16.5 bars. The twigs of the experiment of the second series, in addition to the mentioned dry periods received five periods of dryness of 14-28 days too. In this series of tests, water potential of the H. aphyllum branchlet was diminished at the end of each period in average to -27.2 bars. It is worth mentioning that in order to measure the water potential of branchlets, separate twigs were considered and in each measuring, the branchlets of 5 twigs were cut and used. After the end of dryness period, twigs were watered fully. It was such that their water potential (before sunrise) in the day after irrigation was increased by -5.3 to -4.3 bars. The water potential of control twigs also increased from -12.7 bars in both series of test to about -9.7 to -8.7 bars.
Preparing pressure-volume curves: The impact of dry induction on the elastic and osmotic properties of the branchlet of H. aphyllum twigs became possible through an analysis of pressure-volume curves. These curves were prepared by using Scholander method (Fig. 1).
Analysis of pressure-volume curves
Osmotic potential, turgor potential and water potential: As Fig. 1 also shows, each pressure-volume curve has two outstanding parts: (1) curve part which encompasses about 5 to 8 points and (2) direct part which includes 8 to 11 points. The direct part of curve was used to estimate Ψw, Ψp and Ψs. According to the recommendation of Cutler et al. (1979), this action was performed by drawing a regression curve on at least 7-8 points of the last spots of pressure-volume curves.
The overall equation of the curve is as follows:
In the above relations, (Ψs0)-1 is the inverse of primary osmotic potential of branchlet at full turgescence conditions; m is the slope of regression curve which is under the influence of size of branchlet and osmotic feature of texture and rate of exchange of twig water, (Ψst)-1 is the opposite of Ψs for t times of a pair from the data of pressure-volume and ΣWi is its corresponding accumulated quantity of exited liquid.
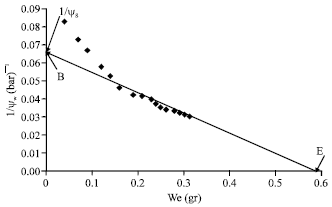 | |
| Fig. 1: | Pressure-volume curve of a branchlet of a H. aphyllum: The horizontal axis is the exit liquid volume (We) and its vertical axis is balancing pressure or (Ψw)-1. Point E, is the place of intersection of osmotic line with We axis showing the rate of water which is exited under the infinite pressure from branchlet known as symplastic water (Ws). Point B is the place of conjunction of osmotic line with the axis of (Ψw)-1 showing that Ψs of branchlet is in the conditions of full turgescence |
With this assumption that the mentioned relation to be fully true in the considered range, by placing each pair of Pi and ΨWi, it is possible to get the Ψs. The continuation of this line crosses the vertical axis in point B which specifies the inverse of osmotic potential in full turgescence (Ψs0)-1 (Tyree and Jarvis, 1982). Subsequently, the turgor potential (Ψp) of the branchlet in each point of pressure-volume curve was obtained through difference of Ψw and Ψs related to the same point. On the other hand, the prolongation of the mentioned regression crosses (cuts) the horizontal axis in point E which determines the volume of exited liquid from branchlet in an infinite pressure [(Ψs0)-1 m-1] (Fig. 1). Active osmotic water of branchlet (WΨs) was calculated in form of Ws (W0-Wd)-1 and its inactive osmotic water in form of 1-Ws (W0-Wd)-1 (Cutler et al., 1979).
Average of absolute value of elastic property (bulk-moduls of elasticity or ![]() ): The existing data in the part of curvature of pressure-volume curve was used to estimate the average of the absolute value of elastic property (
): The existing data in the part of curvature of pressure-volume curve was used to estimate the average of the absolute value of elastic property (![]() ) of the branchlet of H. aphyllum (Tyree and Jarvis, 1982). For the purpose of achieving this parameter, it was necessary that firstly Ψp of the branchlet in the part of curvature of the pressure-volume curve to be calculated. This action was performed by using the osmotic line and method of estimation of osmotic potential, turgor potential and water potential of branchlet. Then the obtained turgor potentials [(Ψp)s] in the mentioned limit with the volume of corresponding condensed exited liquid was drawn in coordinates sheet and their regression equation was calculated (Fig. 2).
) of the branchlet of H. aphyllum (Tyree and Jarvis, 1982). For the purpose of achieving this parameter, it was necessary that firstly Ψp of the branchlet in the part of curvature of the pressure-volume curve to be calculated. This action was performed by using the osmotic line and method of estimation of osmotic potential, turgor potential and water potential of branchlet. Then the obtained turgor potentials [(Ψp)s] in the mentioned limit with the volume of corresponding condensed exited liquid was drawn in coordinates sheet and their regression equation was calculated (Fig. 2).
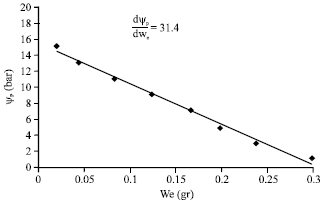 | |
| Fig. 2: | Calculating bulk-moduls of elasticity ( |
The (![]() ) was also calculated from the multiplied by the slope of line [(dΨp) (dWe)-1] in Ws:
) was also calculated from the multiplied by the slope of line [(dΨp) (dWe)-1] in Ws:
In order analyzing the pressure-volume curves and for comparing the impact of dry induction on water parameters between twigs under treatment and control, the statistical method of student’s t-test was used (Snedecor and Cochran, 1980).
RESULTS
Results related to a relatively mild dry induction: At the end of each period of a relatively mild dry induction, Ψw of the branchlet of twigs under the treatment of H. aphyllum reduced in average about -16.5 bars, whereas Ψw of the branchlet of control twigs, remained at a higher limit, i.e., in average -12.7 bars (Fig. 3). Parameters of the active osmotic water (WΨs) and turgor potential at the conditions of wet full saturation (Ψp0) reduced and increased very significantly and reached from 28.8-14% and promoted from 7.3-12.3 bars, respectively. The ability of the elastic of branchlet textures (![]() ) and their water potential (Ψw0) with 99% reliability had a very meaningful increase and accelerated from 30.5-44.8% and from -5.9 to -2.5 bars, respectively but the osmotic potential of stress treatment in full turgescence did not show a meaningful difference as compared with control (Table 1).
) and their water potential (Ψw0) with 99% reliability had a very meaningful increase and accelerated from 30.5-44.8% and from -5.9 to -2.5 bars, respectively but the osmotic potential of stress treatment in full turgescence did not show a meaningful difference as compared with control (Table 1).
The results related to a relatively intensive dry induction: After induction of a relatively intensive drought, the water potential of the twigs branchlets under the treatment of Haloxylon aphyllum diminished in average up to about -27.2 bars whereas, the water potential of the branchlet of control twigs of H. aphyllum remained fixed at the level of mild drought induction (-12.7 bars) more or less (Fig. 4).
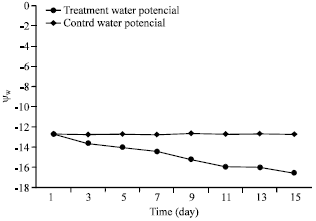 | |
| Fig. 3: | Relationship between shoot (branchlet) water potential (Ψw) and time after seedlings watering. Pre-dawn shoot water potential was measured every other day during a two-week period of water |
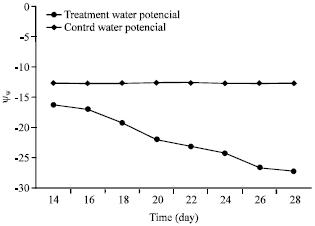 | |
| Fig. 4: | Relationship between shoot (branchlet) water potential (Ψw) and time after seedlings watering. Pre-dawn shoot water potential was measured every other day during a four-week period of water |
| Table 1: | Comparing the results of water relation of the branchlet of H. aphyllum under a relatively mild dry induction with respective control twigs |
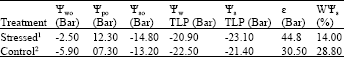 | |
| 1Seedlings, in addition to six early dry periods, five-day dry period 7-14 also received. The average water potential (before sunrise) at the end of their term limit had been reduced to -16.5 bars. Value of each parameter treated seedlings is related to seven seedlings. 2Both seedlings were irrigated once a day. Amount of each parameter of control seedlings is related to five seedlings | |
Water and turgor potentials in fully saturation wet condition had a very meaningful increase (p<1%) and reached to -2 and 18.8 bars, respectively. At the same time, like the mild water stress, the very meaningful increase of elasticity (![]() ) and a very meaningful reduction of osmotic active water (WΨs) in the texture of branchlet were observed. However, concurrent with these changes, the osmotic potential in the condition of wet full saturation (Ψs0) and drought threshold (Ψstlp) diminished in a very meaningful way (p<1%) and decreased from -13.5 to -20.8 bars and from -20.9 to -30.6 bars, respectively.
) and a very meaningful reduction of osmotic active water (WΨs) in the texture of branchlet were observed. However, concurrent with these changes, the osmotic potential in the condition of wet full saturation (Ψs0) and drought threshold (Ψstlp) diminished in a very meaningful way (p<1%) and decreased from -13.5 to -20.8 bars and from -20.9 to -30.6 bars, respectively.
DISCUSSION
Primarily it seemd that in the branchlet of H. aphyllum, the reaction of parameters of water relations like Ψw0, Ψs0, Ψp0, Ψwtlp, ![]() and so on against a very mild relative drought induction to be in conflict with their corresponding parameters at control treatment (Table 1), because in one side, the absolute value of elasticity (
and so on against a very mild relative drought induction to be in conflict with their corresponding parameters at control treatment (Table 1), because in one side, the absolute value of elasticity (![]() ) of twig branchlets under the treatment of Haloxylon were measured to be 44.8 bars, that at the statistical level of 1% in a meaningful rate was more elastic than
) of twig branchlets under the treatment of Haloxylon were measured to be 44.8 bars, that at the statistical level of 1% in a meaningful rate was more elastic than ![]() of branchlets of control twigs (30.5 bars) and on the other side, the osmotic active water of branchlet (WΨs) for treated and controlled twigs were measures to be as 14 and 28.8% and this difference in reliability level of 99% was meaningful. Possibly, the concept of this difference, with regard to the relation of (
of branchlets of control twigs (30.5 bars) and on the other side, the osmotic active water of branchlet (WΨs) for treated and controlled twigs were measures to be as 14 and 28.8% and this difference in reliability level of 99% was meaningful. Possibly, the concept of this difference, with regard to the relation of (![]() ), is that the more elastic branchlet needs a less water to maintain its own turgescence. In addition, in the conditions of full turgescence, the water potentials of twigs which were given drought induction, in a meaningful way were higher than potential of water of controlled twigs.
), is that the more elastic branchlet needs a less water to maintain its own turgescence. In addition, in the conditions of full turgescence, the water potentials of twigs which were given drought induction, in a meaningful way were higher than potential of water of controlled twigs.
Plant physiologists believe that many plants of dry regions through mechanism of elastic property of their own textures are able to maintain turgescence and consequently increase resistance to drought and have continuation of growth. This feature is true for Pinus taeda (Emadian and Newton, 1989), Pseudotsuga menziesii(Joly and Zaerr, 1987), Dubautia ciliotala (Robichaux and Canfield, 1985), Juglans nigra (Parker and Pallardy, 1985). On the other side, though the elastic property of the branchlet of treated twigs increased but their osmotic potential (along with very meaningful increase of water potential) was in lack of any meaningful change. In other words, in the condition of induction of a relatively mild drought induction, no meaningful difference was observed at any statistical level between Ψs0 and Ψstlp of control and drought stress treatments (Table 1). With regard to mentioned conditions, it seems that in the mild drought conditions, H. aphyllum through increase of elastic property of branchlet textures is able to maintain its turgescence and continues it growth.
| Table 2: | Comparing the results of water relation of the branchlet of H. aphyllum under a relatively intensive dry induction with respective control twigs |
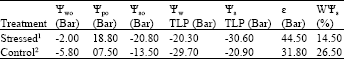 | |
| 1Seedlings, in addition to six early dry periods, five-day dry period 14-28 also received. The average water potential (before sunrise) at the end of their term limit had been reduced to -27.2 bars. Value of each parameter treated seedlings is related to seven seedlings. 2Both seedlings were irrigated once a day. Amount of each parameter of control seedlings is related to five seedlings | |
Apparently, this will be possible only in lieu of an exchange with reduction of osmotic adjustment. In relatively intensive stresses (14-28 days of lack of irrigation), the reaction of different parameters of water relations of H. aphyllum branchlet took place followed by reaction process in the relatively mild stress induction conditions and osmotic adjustment was activated (Table 2). It was such that in addition to elastic property of cellular wall of the textures of branchlet (like the impact of periodical mild drought stresses), the osmotic potential of these textures reduced as compared with controlled textures in a meaningful way and made plants under tension to continue their life in the mentioned conditions. After exposing the twigs of H. aphyllum to a relatively intensive periodical drought stresses, the increase of water potential and turgor potential in the rate of 300 and 250%, respectively and reduction of osmotic potential in the conditions of wet saturation and at the threshold of drought in the rate of more than 50% are considerable and subject to comment. The results showed that the induction of relatively mild and periodical drought was not able to have an impact on the increase of resistance against drought of H. aphyllum and it only changed the strategy of plant in confronting with stresses resulting from change. Instead of that, successive induction of relatively intensive drought could make ideal changes in osmotic parameters and not only cause the viable and freshness of plant but also it enabled twigs to bear the relatively intensive condition of drought and they could maintain their water potential at a very high level. Comparing the parameters of water relation of H. aphyllum in a relatively mild drought stress and those of very intensive ones (Table 3) confirm the importance of the mentioned subject matter. It is such that four important and influential parameters in elucidation of this analysis enjoyed uni-direction changes with an increase of endurance vis-à-vis drought (Ψw0 increased for 0.5 bars, Ψp0 increased by 6.5 bars, Ψs0 decreased by 6 bars, Ψs of branchlet at the threshold of dryness decreased by 7.5 bars).
| Table 3: | Comparing the results of water relation of the branchlet of H. aphyllum under two water regimes: a relatively mild dry and a relatively intensive dry induction |
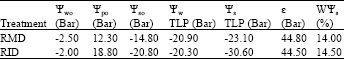 | |
Though under a very relatively intensive and longer drought induction, elastic property of the textures of H. aphyllum was maintained, however, osmotic adjustment was made active to enable twigs maintain their turgescence, to remain alive and continue their physiologic and biochemical activities and consequently their own growth. Reduction of osmotic potential in the conditions of saturation by 6 bars and the fall of this potential at the threshold of wither and dryness by 7.5 bars along with compatibility of twigs to frequent drought stresses might enable them to bear drought periods resulting from factors affecting wet shortage as it is compared with a one-year H. aphyllum, the ones which were not exposed to any kind of intensive drought stress and in a limit of dryness in which similar twigs with controlled twigs face wither, it is possible to be immune against damaging impacts of drought periods. This idea as the symbol of increase of resistance to drought of H. aphyllum (which have received, borne and accustomed to a relatively intensive and periodical drought stresses) can be put forth and to be used as a base for future researches. Anyway, a solid comment on this issue demands further studies and supplementary investigation.
CONCLUSION
The results of this research showed that H. aphyllum bears dryness well. On the other hand, if dryness to be relatively mild and its period is short, by the mechanism of increase of elastic property of textures of branchlet, this species will be able to maintain their turgescence and continue its life. Apparently, this action is possible only in lieu of an exchange with reduction of osmotic adjustment. On the other hands, if dryness is relatively intensive and its period long, for keeping its turgescence, in addition to the mechanism of increase of elastic property uses the mechanism of osmotic adjustment too. In each of the two mentioned conditions, maintaining turgescence in dry conditions cause the continuation of physiologic and biochemical activities of the twigs of this species and make their growth and viability possible despite excessive and long dryness. Based on the results of this research, the executive officials of forestry and production of twigs departments are recommended that if they use H. aphyllum (to have a biologic stability of sand lands in arid and semi-arid zones), prior to transfer of twigs to the main scene of plantation, they should place them under 5 to 8 periods of dry induction for at least 3 to 4 weeks.
ACKNOWLEDGEMENT
I would like to express my sincere gratitude to Organization of Researches, Education and Expansion of Agriculture for supplying me with necessary credits and funds to implement this project. I also wish to thank Dr. Seyed-Fazlallah Emadian, Dr. Ramezanali Khavari-Nejad and Dr. Hossein Heidari Sharifabad for their consultation in developing and implementing this research project. My thanks also go to technicians, drivers and all staff of Center for Agricultural and Natural Resources Researches at Kerman province who has contributed in supplying logistics for the implementation of this project.
REFERENCES
- Ali, M.A., 2011. Pedigree selection for grain yield in spring wheat (Triticum aestivum L.) under drought stress conditions. Asian J. Crop Sci., 3: 158-168.
CrossRefDirect Link - Arabzadeh, N., 2009. The examination of dryness induction effect in growth, some of physiological relations (parameters of water relations), physiological changes and increasing of drought tolerance range in Haloxylon persicum and H. aphyllum. Ph.D. Thesis, Department of Biology Sciences, Teacher Training (Tarbiyat Moallem) University of Tehran, Iran.
- Arabzadeh, N., 2011. A study on the impact of drought stress on changes of free proline in two species of Haloxylon persicum and Haloxylon aphyllum. J. Plant Sci., 6: 190-201.
CrossRef - Arabzadeh, N., 2012. The effect of drought stress on soluble carbohydrates (sugars) in two species of Haloxylon persicum and Haloxylon aphyllum. Asian J. Plant Sci., (In Press).
Direct Link - Arabzadeh, N., R.A. Khavarinejad, S.F. Emadian and H. Heydari-Sharifabad, 2009. A study of the impact of pruning on the growth and vigority of the hand-Planted-haloxylon trees in Kerman, Iran. Asian J. Plant Sci., 8: 474-482.
CrossRefDirect Link - Bilan, M.V., C.T. Hogan and H.B. Carter, 1977. Stomatal opening, transpiration and needle moisture in loblolly pine seedlings from two Texas seed sources. Forest Sci., 23: 457-462.
Direct Link - Cutler, J.M., K.W. Shahan and P.L. Steponkus, 1979. Characterization of internal water relations of rice by a pressure-volume method. Crop Sci., 19: 681-685.
CrossRefDirect Link - Danesh-Shahraki, A., H. Nadian, A. Bakhshandeh, G. Fathi, K. Alamisaied and M. Gharineh, 2008. Optimization of irrigation and nitrogen regimes for rapeseed production under drought stress. J. Argon., 7: 321-326.
CrossRefDirect Link - Emadian, S.F. and R.G. Newton, 1989. Growth enhancement of loblolly pine (Pinus taeda L.) seedlings by silicon. J. Plant Physiol., 134: 98-103.
CrossRef - Gorji, A.H., Z. Zonoori, M. Zolnoori and A. Jamasbi, 2011. Inheritance of antioxidant activity of triticale under drought stress. Asian J. Plant Sci., 10: 220-226.
CrossRefDirect Link - Imanparast, L. and D. Hassanpanah, 2009. Response of onobrychis genotypes to PEG 10000 induced osmotic stress. Biotechnology, 8: 365-369.
CrossRefDirect Link - Jafari, M., H. Azarnivand, H. Tavakoli, G. Zehtabian and H. Esmailzadeh, 2004. Investigation on different vegetation effects on sand dunes stabilization and improvement in Kashan. Pajouhesh Sazandegi, 64: 16-21.
Direct Link - Joly, R.J. and J.B. Zaerr, 1987. Alteration of cell-wall water content and elasticity in douglas-fir during periods of water deficit. Plant Physiol., 83: 418-422.
Direct Link - Khalily, M., M. Moghaddam, H. Kanouni and E. Asheri, 2010. Dissection of drought stress as a grain production constraint of maize in Iran. Asian J. Crop Sci., 2: 60-69.
CrossRefDirect Link - Kufa, T. and J. Burkhardt, 2011. Variations in leaf water potential in the wild Ethiopian Coffea arabica accessions under contrasting nursery environments. J. Agron., 10: 1-11.
CrossRef - Kumari, S., 2010. Cellular changes and their relationship to morphology, abscisic acid accumulation and yield in wheat (Triticum aestivum) cultivars under water stress. Am. J. Plant Physiol., 5: 257-277.
CrossRefDirect Link - Naderikharaji, R., H. Pakniya and A.R. Biabani, 2008. Effect of drought stress on photosynthetic rate of four rapeseed (Brassica napus) cultivars. J. Applied Sci., 8: 4460-4463.
CrossRefDirect Link - Ngaboyisonga, C., K. Njoroge, D. Kirubi and S.M. Githiri, 2009. Effects of low nitrogen and drought on genetic parameters of grain yield and endosperm hardness of quality protein maize. Asian J. Agric. Res., 3: 1-10.
CrossRefDirect Link - Omidi, H., 2010. Changes of proline content and activity of antioxidative enzymes in two canola genotype under drought stress. Am. J. Plant Physiol., 5: 338-349.
CrossRefDirect Link - Parker, W.C. and S.G. Pallardy, 1985. Genotypic variation in tissue water relations of leaves and roots of black walnut (Juglans nigra) seedlings. Physiol. Plantarum, 64: 105-110.
CrossRef - Rad, M.H., M.H. Assare, M.H. Banakar and M. Soltani, 2011. Effects of different soil moisture regimes on leaf area index, specific leaf area and water use efficiency in eucalyptus (Eucalyptus camaldulensis Dehnh) under dry climatic conditions. Asian J. Plant Sci., 10: 294-300.
CrossRefDirect Link - Robichaux, R.H. and J.E. Canfield, 1985. Tissue elastic properties of eight hawaiian Dubautia species that differ in habitat and diploid chromosome number. Oecologia, 66: 77-80.
Direct Link - Safarnejad, A., 2005. Comparison of saxaoul species (Haloxylon spp) for its improvement and expansion in desert areas. Pajouhesh Sazandegi 67: 51-57.
Direct Link - Tokasi, M.V., M. Zahedifar and B. Hemmati, 2007. Determination of digestibility and degradability of tagh (Haloxylon sp.) using in vivo and in vitro and in situ techniques. Pajouhesh Sazandegi, 74: 96-104.
Direct Link - Xoconostle-Cazares, B., F.A. Ramirez-Ortega, L. Flores-Elenes and R. Ruiz-Medrano, 2010. Drought tolerance in crop plants Am. J. Plant Physiol., 5: 241-256.
CrossRef - Scholander, P.F., E.D. Bradstreet, E.A. Hemmingsen and H.T. Hammel, 1965. Sap pressure in vascular plants. Science, 148: 339-346.
CrossRef