
A. Gholamhoseinian Najar
Department of Biochemistry, Afzalipour School of Medicine and Kerman Physiology Research Center
S. Mansouri
Department of Microbiology, Kerman University of Medical Sciences, Kerman, Iran
S. Rahighi
Department of Biochemistry, Afzalipour School of Medicine, Kerman University of Medical Sciences, Kerman, Iran
Asian Journal of Plant Sciences
Year: 2009 | Volume: 8 | Issue: 8 | Page No.: 551-556







ABSTRACT
Myrtus communis L. (Myriaceae) has been reported to have antibacterial activity against Staphylococcus aureus. However, the mechanism of antibacterial activity of this medicinal herb is not so clear. The leaves of M. communis were extracted with chloroform, ethyl acetate and methanol successively and then Total Oligomeric Flavonoids (TOF) were obtained by applying precipitation method. Minimal Inhibitory Concentration (MIC) against Staphylococcus aureus (NTCT 8530) was determined by agar dilution method. Sub MIC (SIC) and 0.1 SIC of different extracts were used for the experiments. Bacteria grown with or without the extracts were lysed with lysophosphatin. The activities of catalase and superoxide dismutase (SOD) were measured by using a spectrophotometric method. Malonedialdehyde (MDA) was determined by thiobarbitoric acid method and the total antioxidant capacity was measured by colorimetry. Specific activity of SOD in bacteria treated by TOF, ethyl acetate, chloroform and methanol extracts at SIC were found to be 0.41, 0.42, 0.38 and 0.51, respectively, compared to 0.93 U mg-1 for the control sample, showing a significant decrease. Catalase specific activity of bacteria in media containing TOF, ethyl acetate, chloroform and methanol extracts were 50.55, 11.97, 46.66 and 46.67 U mg-1, respectively, which showed significant reduction compared to their controls (88.82 U mg-1, p<0.001). All four extracts of Myrtus communis leaves caused a decrease in lipid peroxidation as MDA formation and the total antioxidant activity in S. aureus (p<0.001). It is concluded that the antibacterial effect of Myrtus communis on S. aureus is partly due to induction of free radicals. Ethyl acetate extract showed the highest antibacterial effect comparing to all others. The effects of extracts on free radicals were dose dependent.
PDF Abstract XML References Citation
How to cite this article
A. Gholamhoseinian Najar, S. Mansouri and S. Rahighi, 2009. Effects of Sub-Inhibitory Concentrations of Myrtus communis Leave Extracts on the Induction of Free Radicals in Staphylococcus aureus; A Possible Mechanism for the Antibacterial Action. Asian Journal of Plant Sciences, 8: 551-556.
DOI: 10.3923/ajps.2009.551.556
URL: https://scialert.net/abstract/?doi=ajps.2009.551.556
DOI: 10.3923/ajps.2009.551.556
URL: https://scialert.net/abstract/?doi=ajps.2009.551.556
INTRODUCTION
Infectious diseases and drug resistance to human pathogenic agents have become widespread problems, therefore, therapeutic compounds, that can either inhibit the growth of pathogens or kill them and have no or least toxicity to host cells are considered as candidates for developing new antimicrobial drugs (Robin et al., 1998).
Medicinal plants are known to contain several compounds with antimicrobial properties and the uses of these types of compounds are being increasingly reported from different parts of the world (Saxena and Sharma, 1999). Using plant as medicine represents advantages including low cost, low toxicity, long-time oral consumption and relatively lower toxicity at higher dose. It is expected that plant extracts showing target sites other than those used by antibiotics will be active against drug-resistant microbial pathogens (Ahmad and Beg, 2001; Lee et al., 1998).
Several medicinal plants have antioxidative or oxidative, besides, antimicrobial activities. Antioxidants are considered as possible protective agents reducing oxidative damage of human body, therefore there is a growing interest in the substances exhibiting antioxidant properties that are supplied to human and animal organisms as food components or as specific pharmaceutics (Dawidowicz et al., 2006; Manikandan and Davi, 2005).
The antioxidative properties of plants are related mostly to the presence of phenolic compounds, especially flavonoids (Dawidowicz et al., 2006; Katalinic et al., 2006). Flavonoids, take part very easily in oxidation-reduction processes, both inside and outside the cells (Dawidowicz et al., 2006). The antioxidant power of flavonoids relies on their ability to interact with free radicals which initiate oxidation reactions or which are produced during chain reactions, on the inhibition of oxidation processes (Brown et al., 1998; Cao et al., 1997).
The balance between anti-oxidation and oxidation is believed to be a critical concept in maintaining a healthy biological system. The oxidative stress is the imbalance between oxidants and antioxidants that leads to potential damage and has been suggested to be the cause of aging and various diseases in humans (Dorval and Hontela, 2003; Katalinic et al., 2006). This is the result of an increase in ROS (reactive oxygen species), the impairment of antioxidant defense systems, or incapacity to repair oxidative damage. The main damage induced by ROS results in alterations of cellular macromolecules such as membrane lipids (lipid peroxidation), DNA and/or proteins. The resulting damage may alter cell function through changes in intracellular calcium or intracellular pH, eventually leading to cell death (Dorval and Hontela, 2003; Swann et al., 1991). Main targets of ROS are polyunsaturated fatty acids in cell membranes causing lipid peroxidation and malondialdehyde (MDA) formation, which may lead to damage of the cell structure and function (Kaygusuz et al., 2003).
Myrtus communis L. (Myriaceae) is a perennial shrub belonging to the Myrtaceae family and subfamily Myrtoideae. It is widely distributed in the Mediterranian area (Rotstein et al., 1974) and Iran (Gholamhoseinian et al., 2005). A striking feature of this plant is the pleasant smell its essential oil, present in numerous glands, especially in the leaves. Since ancient times, in traditional medicine, its leaves, fruits and volatile oil were frequently used for several purposes such as food additive, treatment of burns, antidiuretics, antifungal and anti-diabetic effects (Mansouri et al., 2001; Cakir, 2004; Onal et al., 2005).
The antibacterial activity of the leaves of this herb has been previously reported by Sepici et al. (2004) and Mansouri et al. (2001). A potent antioxidant activity of myrtle extracts, mainly due to the presence of galloyl derivatives has been shown and the extract prepared from the leaves of this plant is rich in polyphenols (Romani et al., 2004). Due to antioxidant activity and antimicrobial properties of Myrtus communis leaves, we conducted this study to determine the relation of antioxidant activity to antibacterial activity as a possible mechanism of action.
MATERIALS AND METHODS
Bacterial strain: Staphylococcus aureus (strain NCTC 8530) was obtained from late H.J. Rojers, Biological Laboratory, University of Kent at Canterbury, UK.
Plant material: The plant used in this study was collected from Shahdad City, Kerman Province, in Southeastern part of Iran, during fall 2006. The plant was washed with distilled water and air dried in laboratory at room temperature. The plant was identified by Dr. Mirtajaddini from Department of Biology, Faculty of Science, Shahid Bahonar University of Kerman, Iran. A voucher specimen was preserved and kept at Biochemistry Department of Kerman University of Medical Sciences, Kerman, Iran. Dried leaves were powdered with a blender and subsequently used for the extract preparation.
Preparation of the extracts
Total Oligomeric Flavonoids (TOF) extraction: In order to obtain an extract rich in Total Oligomeric Flavonoid (TOF), the fine air dried powder of Myrtus communis leaves were macerated in a water: acetone mixture (1:2 v/v) for 24 h at 4°C. The extract was filtered with a Buchner funnel. Acetone was evaporated under low pressure to obtain the aqueous phase. Tannins were precipitated and removed after addition of excess NaCl followed by incubation at 4°C for 24 h. The supernatant was extracted with ethyl acetate, concentrated and precipitated with chloroform. The latest precipitate was considered as TOF extract (Hayder et al., 2004).
Methanol, chloroform and ethyl acetate extracts: Methanol, chloroform and ethyl acetate extracts were prepared by cold percolation method (Ramos et al., 2003). Briefly, the air dried powder (100 g) was placed in a modified Buchner funnel. Each of the solvents; chloroform, ethyl acetate or methanol (70%) was poured into the top of the funnel drop wise where it soaked the plant material, leaching out the extract until a clear eluant obtained. The extracts were then concentrated, using a rotatory evaporator and kept in sealed container at 4°C.
All materials were reagent grade and purchased from Merck Co. (Germany).
Determination of Minimum Inhibitory Concentration (MIC) of different extract against Staphylococcus aureus: The Minimum Inhibitory Concentration (MIC) of Myrtus communis leave extract against Staphylococcus aureus was determined using standard agar dilution method (Forbes et al., 2007). Each extract was dissolved in DMSO (dimethyl sulfoxide) and added to the sterile Muller-Hinton Agar (Merck Co., Germany) medium, at 50°C to give the final concentration of 1 up to 150 μg mL-1. Serial two-fold dilutions of different extracts were used for the determination of MIC. Bacterial suspension (10 μL) adjusted to the concentration of 0.5 McFarlands was used for plate inoculation. The plates were incubated at 35°C for 18-24 h. The lowest concentration of the extracts that inhibited the growth of bacterial suspension was regarded as MIC. For all experiments Sub Minimal Concentration (SIC; one dilution lower than MIC) and 0.1 SIC level were used (Gholamhoseinian et al., 2005).
Bacterial lysate preparation: Different methods were used for the lysis of Staphylococcus aureus, such as sonication and freeze-thawing. However, lysostaphin combined with low concentration of Cetyl Trimethyl Ammonium Bromide (CTAB) detergent was finally applied. Briefly, one loop of bacterial colony was inoculated in the Muller Hinton broth in a steady state to reach an optical density at 675 nm equal or near to 1.0. The suspension (1 mL) was centrifuged for 10 min at 5000x g. The precipitate was re-suspended in 400 μL of EDTA solution (100 mM). To this suspension, 2.5 μL lysostaphin solution (1 mg mL-1) and 5 μL of CTAB/NaCl (10% CTAB in 0.7 M NaCl) were added. The mixture was incubated at 60°C for 10 min until the cells were completely lysed. Inability to grow on Muler Hinton agar after 24 h incubation at 37°C was considered as the complete lysis of the sample.
Enzyme activity, total antioxidant measurement and protein determination in bacterial lysate: Ransod and TAS kits (Randox, UK) were used for the determination of Super Oxide Dismutase (SOD) activity and total antioxidant system (TAS) according to the manufacturer protocol.
Catalase was assayed based on the decrease in the absorbance of H2O2 at 240 nm according to Dogruer et al. (2004). Malondialdehyde (MDA) was measured by thiobarbitoric acid method (Van-Ye et al., 1993). One unit of SOD activity was expressed as the amount of lysate which caused 50% dismutation of superoxide radicals under the assay conditions. Catalase activity was defined as the amount of lysate that catalysis the decomposition of 1 μmol H2O2 per min at 25°C. Total protein concentration in cell lysate was determined by method of Lowry (1951) using Bovine serum albumin as the standard. The specific activity of the enzyme activities were expressed as units per milligram of cellular proteins.
Statistical analysis: All experiments were repeated three times. One-way Analysis of Variance (ANOVA) was used for statistical analysis of the effects of different concentration of herb extracts on the antioxidant system using the statistical package SPSS for windows (version 10.5, 2005). Significance of difference was accepted at p<0.05. Data were presented as Mean±SD.
RESULTS
The most active extract of the leaves of M. communis against Staphylococcus aureus was found to be ethyl acetate extract which inhibited the bacterial growth at 5 μg mL-1 concentration, while TOF extract was the least active extract (MIC = 400 μg mL-1). Methanol and chloroform extracts however had similar activities with the MIC of 10 μg mL-1. Therefore, the effect of extracts on the induction of free radicals was evaluated by 200, 5, 5 and 1 μg mL-1 of the TOF, methanol, chloroform and Ethyl acetate extracts (as SIC) respectively and one tenth of these concentrations as 0.1 SIC.
Total antioxidant capacity (Table 1), superoxide dismutase and catalase activities as well as MDA production of the Staphylococcus aureus lysate are presented in Table 2. Total antioxidant activity was reduced significantly in SIC and 0.1 SIC groups of bacteria which were grown in the presence of methanol, TOF, chloroform and ethyl acetate extracts (p<0.001). Superoxide dismutase activity of the bacterial lysate that were grown in the presence or absence of different extracts were 0.38 to 0.93 U mg-1, which were significantly lower than the control group. When SIC and 0.1 SIC were compared for the level of SOD reduction, significant difference was found for TOF and ethyl acetate extracts. Catalase activity was significantly reduced in the SIC treated bacteria, especially in the ethyl acetate extract. When 0.1 SIC was used only TOF and ethyl acetate extracts showed a significant reduction. The level of MDA of SIC group of different extracts were significantly decreased compared to the control group (p<0.001). However, the reduction level of MDA in 0.1 SIC status of methanol extract was not significant when compared to the control (p = 0.74).
| Table 1: | Effect of Myrtus communis leave extracts on total antioxidant capacity of Staphylococcus aureus (Strain NCTC 8530) |
 | |
| Control: Bacteria cultivated under standard condition in the absent of the extract. SIC: Sub minimal inhibitory concentration of the extract. Tabulated values are Mean±SD of 3 experiments. Values on the same column designated with * are significantly different from that of the control at p≤0.05 | |
| Table 2: | Effect of different concentration of extracts prepared from M. communis on superoxide dismutase and catalase activities and malondialdehyde production in S. aureus strain NCTC 8530 |
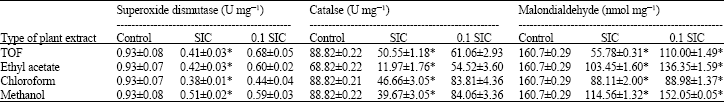 | |
| Control: Bacteria cultivated under standard condition in the absent of the extract. SIC: Sub minimal inhibitory concentration. Tabulated values are Mean±SD of 3 experiments. Values on the same column designated with * are significantly different from that of the control at p≤0.05 | |
DISCUSSION
It has been shown in the pathogenesis of several diseases that free radicals induce tissue damage (Kaygusuz et al., 2003; Dogruer et al., 2001; Inci et al., 2003). Increased lipid peroxidation and decreased antioxidant protection may lead to cytotoxicity, allergy, mutagenicity and carcinogenicity (Mates et al., 1999). Antibacterial activity of Myrtus communis leave extract against Staphylococcus aureus is reported to be very high. This activity was reported to be comparable to that of trimethoprim-sulfamethoxazole (SXT) and inhibited the growth of 99% out of 489 clinical isolate of S. aureus (Mansouri, 1999). The diethyl ether extract fraction of crude methanol extract had the highest activity in comparison with the other fractions and the minimum inhibition concentration ( MIC) for S. aureus was reported to be < 0.05 mg mL-1 (Mansouri et al., 2001).
In this study, the antibacterial activity of the Myrtus communis leaves against Staphylococcus aureus was assessed under two different concentrations of SIC and one tenth of SIC to evaluate the effect of free radical formation as a possible mean of antibacterial action.
The MDA is one of the end products of lipid peroxidation and can inactivate membrane transporters by forming intra- and intermolecular cross links (Kramer et al., 1984). SOD is one of the major intracellular enzymes that catalysis the dismutation of the highly reactive superoxide anion (O2•-) to H2O2 and finally to oxygen which protects cell against oxygen free radicals. Peroxide can be destroyed by catalase and glutation peroxidase (GSP). Catalase has been shown to be responsible for the detoxification of significant amounts of H2O2. Glutathione peroxidase metabolizes peroxides such as H2O2 and protects cell membranes from lipid peroxidation (Madrigal et al., 2001). According to the results of this study the mean specific activity of SOD in the lysate of the standard strain of Staphylococcus aureus at SIC with TOF, ethyl acetate, chloroform and methanol extracts were significantly decreased in comparison with that in the control group grown in the absence of the extracts (p<0.001). These results showed that all forms of the extracts are capable of reducing the activities of the enzymes involved in the detoxification of free radials. Concomitant to this, the total antioxidant capacity of the bacterial cells reduced too and these phenomena would increase free radicals, a situation similar to the hepatocytes of alloxan treated rats (Delibashvili et al., 2002). Addition of sub-minimal inhibitory concentration (SIC) of the extracts to the culture medium decreased MDA production in the bacterial lysate. However, when the antioxidant potency reduces it might be expected to increase the lipid peroxidation. This event was shown in E. coli at high concentration of methanol extract of M. communis, whereas at lower concentration of the extract it has different effect and reduces the MDA production (Gholamhoseinian et al., 2005). However, the chloroform and TOF extracts were almost twice active against lipid peroxidation in comparison to methanol and ethyl acetate extracts in this study. Romani et al. (2004) showed that the myrtle (Myrtus communis L.) extracts are very rich in polyphenols and hydroalcoholic extracts contain galloyl-glucosides, ellagitannins, galloyl-quinic acids and flavonol glycosides whereas; ethyl acetate extract and aqueous residues are rich in flavonol glycosides and hydrolysable tannins. They found a significant difference among hydroalcoholic extracts vs. the ethyl acetate and aqueous residues. On the other hand, the lipids of Staphylococcus aureus membrane grown in the media containing the chloroform or TOF extracts were less protective against peroxidation. This is in contrast to the total antioxidant capacity of the plant extract which was decreasing from hydroalcoholic to ethyl acetate and aqueous extracts (Romani et al., 2004). The phenomenon that was seen at two different concentrations in this study, suggests a protective effect for the membrane of Staphylococcus aureus by the extract. Unexpectedly less MDA was produced under SIC condition compare to the 0.1 SIC.
The biological activities of M. communis leaves had at least been related to myrtucommulone-A and myrtucommulone-B (Kashman et al., 1974; Rotstein et al., 1974). These activities could be through the reduction of total antioxidant potency of the cell extract as well as the reduction of the activities of the related enzymes such as SOD and catalase showed in this study. The hydroalcoholic percolation extracts showed an important free-radical scavenging activity towards 1,1-diphenyl-2-picrylhydrazyl (DPPH) radical and suggested the future utilization of these extracts as additives in chemoprevention studies (Ramos et al., 2003). M. communis has been reported to have similar antibacterial activity as the trimethoprime-sulfamethoxazole, which was active against bacterial DNA (Mansouri, 1999). Flavonoids are the most likely candidates among the compounds known to be present in the ethanol extract, for providing the anti-mutagenic effect and preventing oxidative lesions (Ahmad and Beg, 2001; Dawidowicz et al., 2006). In Melissa officinalis L. extracts the most effective antibacterial activities had been found by petroleum ether and ethyl acetate fractionation which is mostly related to their phenol content (Canadanovic-Brunet et al., 2008). It had been shown that, hydroxy radicals directly trapped by eugenol and is metabolized to a dimer called dipropofol, this compound has strong antibacterial activity against Gram-positive bacteria (Ogata, 2008). Phenol composition of M. communis L. extract might have similar action in reduction of the scavenging free radicals and inhibition of enzymes involved in this system.
No preference was found between various extract preparations studies for their effects on TAS or SOD activities at SIC level. The catalase activity of ethyl acetate extract at SIC level was about four to five times less than that of the other preparations. The effects of all different extracts that screened in this study were dose dependent. This was similar to the dose dependent inhibition of matrix metalloproteinase by crude ethanol extract of Pothomorphe umbellata L.Miq (pariparoba) (Barros et al., 2007).
In conclusion, M. communis showed two opposite effects: a protective effect by reduction of peroxidation of lipids revealed by the rate of MDA production and a destructive action presented by decreasing free radical scavenging activities through reduction of activities of the enzymes involved and total antioxidant capacity. These two effects are not in harmony with each other. The overall effect was in favor of oxidation. As mentioned earlier, since the effect of M. communis extract on Staphylococcus aureus was reported to be similar to trimethoprimsulfamethoxazol (SXT) which was involved in de novo synthesis of DNA (Mansouri et al., 2001), further work on peroxidation in relation to synthesis and structure of DNA is recommended.
ACKNOWLEDGMENTS
This study was supported by the research funds provided by the Vice Chancellor for Research and the Kerman Physiology Research Center, Kerman University of Medical Sciences, Kerman, Iran. We appreciate the assistance of Dr. Nozar Nakhaie from the Department of Epidemiology, Afzali-pour School of Medicine in Kerman for the statistical analysis, Dr. Masoud Mirtajaddini from the Biology Department, Faculty of Science, Bahonar University of Kerman for the identification of the plant samples and Mr. Ali Vaziri Gohar for preparation of first draft of the study.
REFERENCES
- Ahmad, I. and A.Z. Beg, 2001. Antimicrobial and phytochemical studies on 45 Indian medicinal plants against multi-drug resistant human pathogens. J. Ethnopharmacol., 74: 113-123.
CrossRefDirect Link - Barros, L.F.M., P.S.M. Barros, C.D. Ropke, V.V. Silva, T.C.H. Sawada, S.B.M. Barros and R. Belfort, 2007. Dose-dependent in vitro inhibition of rabbit corneal matrix metalloproteinases by an extract of Pothomorphe umbellata after alkali injury. Braz. J. Med. Biol. Res., 40: 1129-1132.
CrossRef - Brown, J.E., H. Khodr, R.C. Hider and C.A. Rice-Evans, 1998. Structural dependence of flavonoid interaction with Cu2+ ions: Implication for their antioxidant properties. Biochem. J., 330: 1173-1178.
Direct Link - Canadanovic-Brunet, J., G. Cetkovic, S. Djilas, V. Tumbas and G. Bogdanovic et al., 2008. Radical scavenging, antibacterial and antiproliferative activities of Melissa officinalis L. extracts. J. Med. Food, 11: 133-143.
PubMed - Cao, G., E. Sofic and R.L. Prior, 1997. Antioxidant and prooxidant behavior of flavonoids: Structure-activity relationships. Free Radical Biol. Med., 22: 749-760.
CrossRefDirect Link - Dawidowicz, A.L., D. Wianowska and B. Baraniak, 2006. The antioxidant properties of alcoholic extracts from Sambucus nigra L. (antioxidant properties of extracts). LWT-Food Sci. Technol., 39: 308-315.
Direct Link - Dogruer, Z., M. Unal, G. Eskandari, Y.S. Pata, Y. Akbas, T. Cevik and M.Y. Burak-Cimen, 2004. Malondialdehyde and antioxidant enzymes in children with obstructive adenotonsillar hypertrophy. Clin. Biochem., 37: 718-721.
CrossRefPubMedDirect Link - Dogru, H., N. Delibas, F. Doner, M. Tuz and K. Uygur, 2001. Free radical damage in nasal polyp tissue. Otolaryngol. Head Neck Surg., 124: 570-572.
PubMed - Dorval, J. and A. Hontela, 2003. Role of glutathione redox cycle and catalase in defense against oxidative stress induced by endosulfan in adrenocortical cells of rainbow trout (Oncorhynchus mykiss). Toxicol. Applied Pharmacol., 192: 191-200.
CrossRefPubMedDirect Link - Hayder, N., A. Abdelwahed, S. Kilani, R.B. Ammar, A. Mahmoud, K. Ghedira and L. Chekir-Ghedira, 2004. Anti-genotoxic and free-radical scavenging activities of extracts from (Tunisian) Myrtus communis. Mutat. Res. Genet. Toxicol. Environ. Mutagen., 564: 89-95.
CrossRefPubMedDirect Link - Katalinic, V., M. Milos, T. Kulisic and M. Jukic, 2006. Screening of 70 medicinal plant extracts for antioxidant capacity and total phenols. Food Chem., 94: 550-557.
CrossRefDirect Link - Kaygusuz, İ., N. İlhan, T. Karlidag, E. Keles, S. Yal�ins and H. �etİner, 2003. Free radicals and scavenging enzymes in chronic tonsillitis. Otolaryngol. Head Neck Surg., 129: 265-268.
PubMed - Kramer, J.H., T. Mak and W.B. Weblicki, 1984. Differential sensitivity of canine cardiac sarcolemmal and microsomal enzymes to inhibition by free radical induced lipid peroxidation. Circ. Res., 55: 120-124.
PubMed - Lee, C.K., H. Kim, K.H. Moon and K.H. Shin, 1998. Screening and isolation of antibiotic resistance inhibitors from herb materials-resistance inhibition of volatile components of Korean aromatic herbs. Arch. Pharm. Res., 21: 62-66.
PubMed - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Madrigal, J.L., R. Olivenza, M.A. Moro, I. Lizasoain, P. Lorenzo, L. Rodrigo and J.C. Leza, 2001. Glutathione depletion, lipid peroxidation and mitochondrial dysfunction are induced by chronic stress in rat brain. Neuropsychopharmacology, 24: 420-429.
PubMed - Manikandan, S. and R.S. Devi, 2005. Antioxidant property of α-asarone against noise-stress-induced changes in different regions of rat brain. Pharmacol. Res., 52: 467-474.
CrossRefDirect Link - Mansouri, S., A. Foroumadi, T. Ghanei and A.G. Najar, 2001. Antibacterial activity of the crude extracts and fractionated constituents of Myrtus communis. Pharm. Biol., 39: 399-401.
Direct Link - Mates, J.M., C. Perez-Gomez and I.N. de Castro, 1999. Antioxidant enzymes and human diseases. Clin. Biochem., 32: 595-603.
CrossRefPubMedDirect Link - Ogata, M., 2008. Antioxidant and antibacterial activities of dimeric phenol compounds. Yakugaku Zasshi, 128: 1149-1158.
CrossRef - Ramos, A., A. Visozo, J. Piloto, A. Garcıa, C.A. Rodrıguez and R. Rivero, 2003. Screening of antimutagenicity via antioxidant activity in Cuban medicinal plants. J. Ethnopharmacol., 87: 241-246.
CrossRefPubMedDirect Link - Huebner, R.E., A. Wasas, A. Mushi, L. Mazhani and K. Klugman, 1998. Nasopharyngeal carriage and antimicrobial resistance in isolates of Streptococcus pneumoniae and Haemophilus influenzae type b in children under 5 years of age in Botswana. Int. J. Infect. Dis., 3: 18-25.
CrossRefPubMedDirect Link - Romani, A., R. Coinu, S. Carta, P. Pinelli, C. Galardi, F.F. Vincieri and F. Franconi, 2004. Evaluation of antioxidant effect of different extracts of Myrtus communis L. Free Radic Res., 38: 97-103.
PubMed - Rotstein, A., A. Lifshitz and Y. Kashman, 1974. Isolation and antibacterial activity of acylphloroglucinols from Myrtus communis. Antimicrob. Agents Chemother., 6: 539-542.
PubMed - Sepici, A., I. Gurbuz, C. Cevik and E. Yesilada, 2004. Hypoglycaemic effects of myrtle oil in normal and alloxan-diabetic rabbits. J. Ethnopharmacol., 93: 311-318.
CrossRefPubMedDirect Link - Swann, J.D., M.W. Smith, P.C. Phelps, J.D. Swann and M.W. Smith et al., 1991. Oxidative injury induces influx-dependent changes in intracellular calcium homeostasis. Toxicol. Pathol., 19: 128-137.
PubMed - Van Ye, T.M., A.M. Roza, G.M. Pieper, J. Henderson, C.P. Johnson and M.B. Adams, 1993. Inhibition of intestinal lipid peroxidation does not minimize morphologic damage. J. Surg. Res., 55: 553-558.
CrossRefPubMedDirect Link