
Jia-Liang Zhang
College of Plant Science and Technology, Huazhong Agricultural University, Wuhan, 430070, China
Dong-Hua Liu
College of Plant Science and Technology, Huazhong Agricultural University, Wuhan, 430070, China
Zhi-Hua Wang
College of Plant Science and Technology, Huazhong Agricultural University, Wuhan, 430070, China
Can Yu
College of Plant Science and Technology, Huazhong Agricultural University, Wuhan, 430070, China
Jin-Hua Cao
College of Plant Science and Technology, Huazhong Agricultural University, Wuhan, 430070, China
Chun-Tai Wang
Biotechnological Laboratory of National Commission for Nationalities, South-Central University for Nationalities, Wuhan, 430074, China
De-Ming Jin
College of Plant Science and Technology, Huazhong Agricultural University, Wuhan, 430070, China
Asian Journal of Plant Sciences
Year: 2009 | Volume: 8 | Issue: 4 | Page No.: 285-292







ABSTRACT
Many studies on mechanisms under drought stress have been based on gene expression. Quantitative Real-Time PCR (QRT-PCR) is the most sensitive method to detect the transcript of interest gene. To avoid bias, expression of the target gene is usually normalized relative to one or multiple reference genes which should not fluctuate in all samples. The most widely used reference genes are those which belong to actin family; however, many researchers make use of paralogous genes for actin without proper validation of their presumed stability of expression. If an unstable housekeeping gene (HKG) is selected to normalize the target, the results obtained may be spurious. So, we evaluated the gene expression of 10 HKGs, including 8 paralogous genes for actin, together with b-tub and eEF-1a in rice. For the entire sample pools analyzed, ACT(X16280)1 and eEF-1a were the most stable genes in the different stages of rice panicle development. In this study, we used ACT(X16280)1, eEF-1a and the geometric mean of ACT(X16280)1 and eEF-1a as internal controls to study the relative expression of gene GS3, controlling grain length and weight, during different stages of panicle development under drought stress. A sharp increase of GS3 expression (up to 4.8 fold) at spikelet primordium differentiation stage was observed under drought stress. The shrinking of grain size under drought stress could be attributed to the over expression of GS3 which is a negative regulator for grain size.
PDF Abstract XML References Citation
How to cite this article
Jia-Liang Zhang, Dong-Hua Liu, Zhi-Hua Wang, Can Yu, Jin-Hua Cao, Chun-Tai Wang and De-Ming Jin, 2009. Expression Pattern of GS3 During Panicle Development in Rice under Drought Stress: Quantification Normalized Against Selected Housekeeping Genes in Real-Time PCR. Asian Journal of Plant Sciences, 8: 285-292.
DOI: 10.3923/ajps.2009.285.292
URL: https://scialert.net/abstract/?doi=ajps.2009.285.292
DOI: 10.3923/ajps.2009.285.292
URL: https://scialert.net/abstract/?doi=ajps.2009.285.292
INTRODUCTION
Abiotic stresses, including drought, cold and high salinity, greatly reduce average yields for most major crop plants by more than 50%. Drought is perhaps the major factor limiting crop production worldwide. The increasing scarcity of water for irrigation is also a major concern in China. For instance, about 45% of the world agricultural lands are subject to continuous or frequent drought.
Grain weight, number of grains per panicle and number of panicles per plant are the most important components of grain yield. GS3, a major Quantitative Trait Loci (QTL) for grain length and weight and also a minor QTL for grain width and thickness has been mapped in a recent study of Fan et al. (2006). This gene consists of five exons and encodes 232 amino acids with a nonsense mutation; this mutation causes a 178-aa truncation in the C-terminus of the predicted protein, suggesting that GS3 may function as a negative regulator for grain size. Rice is particularly sensitive to drought stress during reproductive growth, when even moderate stress can result in drastic reduction in grain yield. The decrease of seeds setting percentage and grain weight contribute a lot to the reduction in grain yield, the change in GS3 gene expression may be related to the decrease of grain weight under drought stress. Thus, the study on the expression of related gene(s) under stress will provide a key to the molecular understanding on the mechanisms of the yield loss resulting from drought.
Quantitative Real-Time PCR (QRT-PCR) is at present the most sensitive method for the analysis of gene expression compared with band densitometry and probe hybridization (Schmittgen et al., 2000). This technique combines accuracy, sensitivity, dynamic range and reproducibility with speed and potential for high throughput (Bustin, 2000). To avoid bias expression of the interest gene is usually normalized relative to that a reference gene which should have constant expression in all samples. If the internal control gene has a large variability or is altered by the conditions of experiment, the results obtained may be spurious. To increase the reliability, the use of multiple reference genes is adopted by Tricarico et al. (2002) and Vandesompele et al. (2002). A number of housekeeping genes (HKGs), such as actin, tubulin, 18S rRNA, eEF-1a, GAPDH, expressed protein, have been used as reference genes in QRT-PCR analysis (Caldana et al., 2007; Jain et al., 2006a; Kim et al., 2003). After searching in 7 different databases (SCIE, Science Direct, Springer, Nature, Oxford journals, Blackwell Synergy, Science Online), 64 related studies in rice were found from January 1994 to August 2008, the most widely used reference genes belong to actin family (Table 1). Most of these studies make use of paralogous genes for actin without validation of their presumed stability of expression. However, the M values analyzed by geNorm show remarkable difference between paralogous genes UBQ5 and UBQ10 (Jain et al., 2006a). Significant differences were also found between GAPDH1 and GAPDH2 and between ACTB1and ACTB2 (Infante et al., 2008). To screen the most stable gene(s) for internal control is crucial to obtain reliable result in QRT-PCR analysis.
In this study, the expression of 10 genes including 8 paralogous genes for actin (most widely used), b-tub and eEF-1a were evaluated to screen the most stable gene(s) for internal control and ACT(X16280)1, eEF-1a and the geometric mean of ACT(X16280)1 and eEF-1a was selected as appropriate internal controls to determine the expression levels of the gene GS3 under drought stress.
MATERIALS AND METHODS
Biological materials and stress treatment: The experimental materials were grown under a rain-out shelter of State Key Laboratory of Crop Genetic Improvement of Huazhong Agricultural University in 2008. Rice (Chuanxian955, 9311) seeds were germinated on wet filter paper in plastic plates for 3 days at 25°C in May 2008. When the radicle reached a length of 2-3 cm, the germinated seeds were then transplanted to a nursery box filled with soil. After 3 weeks, seedlings were transferred to PVC tubes (1 m in depth and 20 cm in diameter), two plants per tube in June 2008. A randomized complete block design of three replications was performed. All tubes were placed under a movable rain-shelter.
The experiment consisted of two treatments: a well-watered control and a drought stressed treatment. For the well-watered control, water was added to the tubes daily until maturity. For drought stressed treatment, tubes were irrigated as the control did before panicle differentiation and were then left for 18 days without irrigation until young panicles were sampled in early developmental stages.
RNA extraction and quantification: The molecular experiment was conducted in central laboratory of Huazhong Agricultural University in 2008. RNA was extracted from 100 mg of rice young panicle using Trizol regent (invitrogen) according to the manufacturer’s instructions. Total RNA (20 μg) was incubated with 10 U of RNase-free DNase I (Takara) to digest contaminating genomic DNA. Purity of the total RNA extracted was determined as the 260/280 nm ratio and the integrity was checked by electrophoresis in 0.8% agarose gel.
Reverse transcription: One microgram of RNA was reverse transcribed using the Rever Tra Ace® kit (ToYoBo, Shanghai, China) in a 20 μL total volume containing 4 μL 5xRT buffer, 4 μL Rnase nhibitor (10 U μL-1), 2 μL dNTP mixture (10 mM), 1 μL ReverTra Ace, Oligo(dt)20 primer. The reaction was incubated at 42°C for 20 min and then for 5 min at 99°C to inactivate the ReverTra Ace and was then cooled to 4°C. The cDNAs were then stored at -20°C until used in QRT-PCR.
Real-time quantitative PCR: QRT-PCR was performed with a Rotor Gene 2000 Real-Time PCR detection system (Corbett, Australia), by using SYBR green dye bound to double strand DNA at the end of each elongation cycle (SYBR Green Real-time PCR Master Mix; TOYOBO, Osaka, Japan). Each 20 μL reaction mixture contained 10 μL of 2xSYBR Green PCR Master Mix, 0.4 μL of each primer (10 μM), 2 μL of sample cDNA and 7.2 μL sterilized ultra pure H2O. PCR conditions were 95°C for 200 sec, followed by 40 cycles of 95°C for 15 sec, 60°C for 15 sec and 72°C for 15 sec. No-template controls were also included to detect any spurious signals arising from amplification of any DNA contamination or primer dimer formed during the reaction (Iskandar et al., 2004). A negative control sample was considered to be negative if the threshold cycle (Ct) for amplification was >5 cycles greater than the corresponding test sample (Fleige et al., 2006).
After amplification, a melting curve was created at 55-99°C to verify the specificity of the PCR reactions and each reaction mixture were analyzed by 2% agarose gel electrophoresis to evaluate amplification of nonspecific products. Standard curves were generated to determine reaction efficiency using serial dilution series of a cDNA pool of all samples (Hoogewijs et al., 2008). The Rotor-Gene 2000 Version 4.6 software was employed to obtain the threshold cycle (Ct) for best fit (least squares method) of the standard curve.
Data analysis: Ct values from the Rotor-Gene 2000 Detection System were exported to qBase Version 1.3.5, a free program (Hellemans et al., 2007) to obtain relative quantities. These data from qBase was analyzed by geNorm 3.4 software (Vandesompele et al., 2002). The relative expression ratios were calculated using the expression 2-ΔΔCT as described by Caldana et al. (2007).
RESULTS
The selection of housekeeping genes: We performed a search of studies in 7 different databases from January 1994 to August 2008. There are 64 related studies using QRT-PCR in rice and the HKGs are shown in Table 1. It was remarkable that actin (27 times; 42.2%) was used as internal control genes in nearly half of the studies.
Some actin genes, however, showed instability under special experimental conditions in plants and were unsuitable as reference genes (Caldana et al., 2007; Jain et al., 2006a; Nicot et al., 2005; Tu et al., 2007). Caldana et al. (2007) found that actin1 was one of the most stable gene in all tested conditions by comparing the expression of 7 HKGs in rice, whereas, act11 only ranked sixth out of 11 HKGs in all 25 cDNA samples (Jain et al., 2006b).
Considering the significant difference of stability among actin family, we selected 10 HKGs: 8 paralogous genes for actin, together with eEF-1a and b-tub to screen the best internal control for present study (Table 2).
PCR efficiency and amplification specificity: Reaction efficiency estimates are derived from standard curve that are generated using a serial dilutions (1, 1: 10, 1: 100 and 1: 1000) of all samples (Jain et al., 2006b; Tsuchiya et al., 2004). The efficiency values of the ten HKGs and the gene of interest, GS3, were in the range 1.94 to 2.01. Amplification of the specific transcript was confirmed by the appearance of a single peak in the melting curve analysis following completion of the amplification reaction. The amplified products were further analyzed by 2% agarose gel electrophoresis and ethidium bromide staining. Only a single band with the expected size was detected in each experiment, indicating good specificity of all the primer sets in QRT-PCR.
Expression profile and expression stability of the housekeeping genes: To analyze the transcription level of each HKG during different stages of panicle development, total RNA was isolated from a variety of samples under drought treatment. With the same threshold setting, the 10 candidate reference genes used for the data analysis displayed a relatively wide range of expression level (Fig. 1), from the lowest mean Ct-value (21.20) in ACT(AK100267) to the highest (26.84) in ACT(XP_475316.1).
| Table 1: | Reference genes used as internal control for gene expression studies in rice |
 | |
| Results of a search of studies from January 1994 to August 2008 with 64 studies in 7 different databases (SCIE, Science Direct, Springer, Nature, Oxford journals, Blackwell Synergy, Science Online) | |
| Table 2: | Reference genes evaluated and target gene used in real-time PCR assay |
 | |
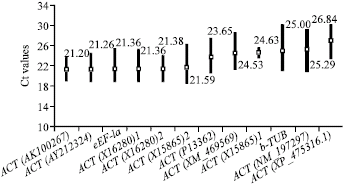 | |
| Fig. 1: | Raw Ct values of the 10 reference genes for drought treatment. Black bars spread from the minimum to the maximum Ct and boxes represent mean Ct-values |
There might be a difference in Ct-values resulted from different primer pairs for the same HKG. ACT(X15865)2 gave a lower mean Ct (21.59) than that of the ACT (X15865)1 (24.63), whereas the primers of ACT(X16280)1 and ACT(X16280)2 show similar mean Ct values (21.36 and 21.38). Ct values give a direct indication of the sensitivity of detecion of a cDNA template.
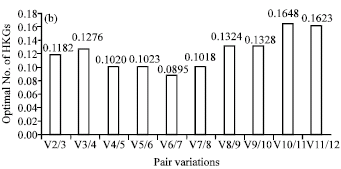
The transcriptional stability of the 10 HKGs was subsequently assessed with the geNorm program. Vandesompele et al. (2002) defined two parameters to quantify the HKG stability: M (average expression stability) and V (pairwise variation). Genes with the lowest M values have the most stable expression. V-value of 0.15 was proposed as the default cut-off value, below which the inclusion of an additional reference gene is not required. For drought stress, the most stable genes were ACT(X16280)1 and eEF-1a. The M value obtained for these two genes was 0.384 (Fig. 2a) and the V2/3 value was 0.118 (Fig. 2b), so there was no need to add a third gene as an internal control according to the standard proposed by Vandesompele et al. (2002).
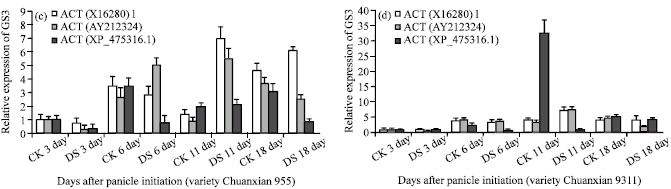
Normalization analysis of GS3 gene expression: Under drought stress, the most stable HKGs were ACT(X16280)1 and eEF-1a assessed by geNorm. The expression pattern of the gene of interest, GS3 was quantified with each reference gene (ACT(X16280)1 or eEF-1a) and two together (ACT(X16280)1 + eEF-1a) as internal control during different stages of rice panicle according to geNorm instructions (Vandesompele et al., 2002). For the rice variety Chuanxian955 (Fig. 3a). The expression levels of gene GS3 under drought stress were slightly lower than that of the control in early stages of panicle development (3-6 day after panicle initiation).
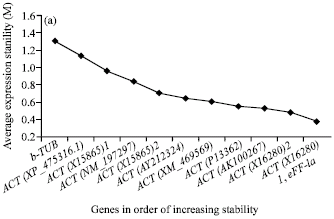 | |
| Fig. 2: | Average expression stability values of HKGs (a) and determination of the optimal number of HKGs for normalization (b) by geNorm analysis |
At 11 day after panicle initiation (spikelet primordium differentiation stage), however, there was a sharp increase of GS3 expression under drought stress, the maximum expression level increased up to 4.8 fold compared with the control. The GS3 expression under drought stress leveled up in later stage but remained higher than that of the control at 18 day after panicle initiation. For the variety 9311 (Fig. 3b), the gene expression of GS3 during different stages showed a similar pattern though the differences between the drought stressed materials and the control were smaller as compared with Chuanxian955.
To show the impact of using the unsuitable paralogous gene for actin (the most widely used HKG) as an internal control gene, three different stable HKG: ACT(X16280)1(the most stable HKG), ACT(AY212324) (the medium stable HKG) and ACT(XP_475316.1) (the least stable HKG) were selected as an internal control for analysis of GS3 expression. For variety 9311, when using the least stable HKG: ACT(XP_475316.1) as a unique internal control, the expression level at 11 or 18 day of treatment is down-regulated, instead of up-regulated relative to the most stable HKG: ACT(X16280)1 (Fig. 3d), moreover, the expression level of GS3 at 11 day of treatment was so lower and decreased 32 fold compared with the control. When normalized only to the medium stable HKG: ACT(AY212324), the expression level of GS3 at 18 day of treatment is significantly down-regulated, instead of up-regulated relative to the most stable HKG (ACT(X16280)1) (Fig. 3d).
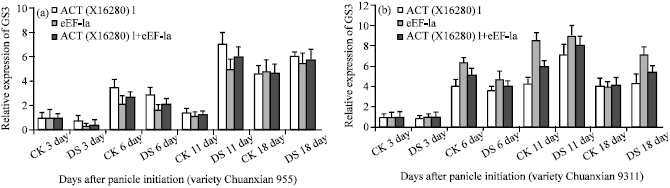 | |
| Fig. 3: | Relative expression of GS3 using different reference genes determined by geNorm as internal controls in different developmental stages of rice panicle under drought stress. The most stable HKG: (a) ACT(X16280)1, eEF-1a and the geometric average of ACT(X16280)1 and eEF-1a were used as internal control for variety Chunxian955, (b) 9311, (c) the most stable HKG: ACT(X16280)1, the medium stable HKG: ACT(XP_475316.1) and the least stable HKG: ACT(XP_475316.1) were used as internal control for variety Chunxian955 and (d) 9311, CK: Control; DS: Drought stress |
| Table 3: | A comparison of the rice grain shape and grain weight under drought stress and control |
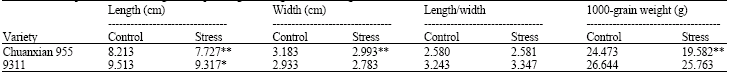 | |
| *,**Significant at p = 0.05 and 0.01, respectively | |
For variety Chuanxian955, the implication of using an unstable reference gene also lead to greatly erroneous results (Fig. 3c).
The change of rice grain shape and grain weight caused by drought stress: The values of grain length, width and 1000 grain weight decreased significantly (p≤ 0.01) under drought stress in Chuanxian 955. The length-width ratio, however, remained unchanged. The shrinking of grain size under drought stress could be attributed to the over expression (starting from the stage spikelet primordium differentiation) of GS3 which is a negative regulator for grain size (Table 3).
Except for a decrease in grain length (significant at p = 0.05), there was no other trait under drought stress that differed significantly from those of control condition in 9311. This is consistent with the relative lower increase of GS3 expression under drought stress (Fig. 3b). These results suggests that 9311 might be a more tolerant variety to drought stress than Chuanxian 955.
DISCUSSION
QRT-PCR has become a popular method to study the expression of interested genes by normalized to an appropriate internal control in a variety of tissues and different treatments (Janovick-Guretzky et al., 2007). A reliable endogenous reference gene should be stable, whereas the expression of interested gene may change greatly over the course of an experiment (Dean et al., 2002). Thus, the selection of appropriate HKG is very important to obtain consistent and reliable results. The majority of studies in the study employ a single gene as internal control and the most widely used reference genes are those belong to actin family (Table 1).
In an earlier study, ACT(X55749) was sfuggested to be an inappropriate internal control gene among the 7 reference genes during biotic and abiotic stresses in potato (Nicot et al., 2005). Another study in cotton also suggested that ACT (AY305733) was not a suitable internal control in a diverse set of 21 cotton samples (Tu et al., 2007). However, ACT (EC969944) was found to be the most stable HKG in a set of samples of berry (Reid et al., 2006). The above reported results suggest that the stability of othologous genes for actin varied greatly in different plant species under a given set of conditions. Therefore, the suitability of a given gene need to be tested even if its orthologue gene in another species is an adequate reference gene.
In order to verify the stability of paralogous genes for actin and select the most stable HKGs in rice. In present study, the variability of expression of 8 paralogous genes for actin, together with b-tub and eEF-1a were studied under drought stressed condition. And geNorm was used to determine the expression stability of HKGs. For all the samples analyzed, ACT(X16280)1, selected as internal control by Toyota et al. (2006), was one of the most stable HKG. This was consistent with the earlier result of a relatively stable expression of the ACT(X16280)1 under several conditions (McElroy et al., 1990). ACT(X15865)2, used to normalize the expression of OsPI1gene in rice by Wasaki et al. (2003), was also found to be an appropriate internal control in this experiment. Other paralogous genes for actin were once used to normalized the expression of interested genes in rice by Duan et al. (2007), Imin et al. (2006), Wasaki et al. (2003), Yu et al. (2007), Zhao et al. (2006), however, the suitability of these actin genes as internal control had not been verified beforehand. The present study indicated that these genes were not stably expressed and might not be suitable to be used as internal control.
Numerous studies suggested that the expression of some HKGs can vary under given conditions (Thellin et al., 1999). Most studies on HKGs selection were made using human or animal tissues. Only a few was focused on plants such as rice (Kim et al., 2003; Jain et al., 2006a; Caldana et al., 2007). Caldana et al. (2007) compared the expression of seven HKGs in rice under salt stress. ACT (P13362) was more stably expressed than ACT (XP_475316.1) in root and shoot. But ACT (XP_475316.1) was a more stable gene than ACT (P13362) under salt stressed condition. However, we found that ACT(XP_475316.1) was the least stably expressed gene and ACT(P13362) ranked second out of 8 paralogous gene for actin in panicle under drought stress. Different experimental conditions may contribute a lot to the variation of stabilily of HKGs. The change of experimental materials (organism or tissue) may be also affecting the stability of the expression of the same reference gene. Singh and Green (1993) and Jain et al. (2006a) pointed out that no one gene has a stable expression under every experimental condition and they thought this may partly be explained by the fact that HKG are not only implicated in the cell metabolism but also participate in other cellular functions in different plant species and tissues. For example, the paralogous gene for actin with stable expression in one organism may be unstable in another organism. And a HKG which is stably expressed in a given condition may not be a stable reference gene in another experiment condition. In addition, eEF-1a was found to be one of the two most stably expressed gene (ACT(X16280)1, eEF-1a) for drought stress. Expression of eEF-1a was previously reported very stable and used as an internal control in animal tissues (Czechowski et al., 2004; Reid et al., 2006). And eEF-1a was also reported to be a stable reference gene in rice under hormone treatments and osmotic stress (Jain et al., 2006a), which was consistent with present study.
Selected reference genes were used to evaluate the gene expression of GS3 under drought stress in this study. The expression level normalized with ACT(X16280)1 was similar to those obtained with eEF-1a or the geometric mean of ACT(X16280)1 and eEF-1a. ACT(X16280)1 has been used as a reference gene to study the expression of starch metabolism-related plastidic translocator genes in rice (Toyota et al., 2006). ACT(X16280)1, eEF-1a, or the geometric mean of ACT(X16280)1 and eEF-1a could be reliable internal control for studying the expression of other target genes controlling grain yield and quality traits, such as GW2 (Song et al., 2007), Ghd7 (Xue et al., 2008), under drought stress in rice.
CONCLUSIONS
The stability of different housekeeping genes vary greatly in rice under drought stress. ACT(X16280)1, eEF-1a were found to be the most reliable reference genes for q PCR studies in rice panicle under drought stress, whereas b-Tub, ACT(XP_475316.1) were less stable genes among the tested HKGs. Appropriate selection of internal control is crucial for analyzing the expression of target gene under drought stress by real-time PCR. The use of unstable HKG may lead to false results. For example, in variety 9311, the GS3 expression level at 11 or 18 days of treatment appeared to be down-regulated when normalized by the least stable HKG: ACT(XP_475316.1); while it showed to be up-regulated when the most stable HKG: ACT(X16280)1 was used as an internal control. To increase the stability of internal control, the geometric mean of two HKGs, ACT (X16280)1 and eEF-1a, could be used in data analysis of real-time PCR experiment.
ACKNOWLEDGMENTS
This research was supported by The Rockefeller Foundation (2004FS070) and State Science and Technology Project on Food Prodution Enhance of China (2004BA520A02). The authors are grateful to Prof. Li-zhong Xiong of National Key Laboratory of Crop Genetic Improvement in Huazhong Agricultural University for his valuable advices and generous help on the research and to prof. Camila Caldana of Max-Planck Institute of Molecular Plant Physiology, Germany for his providing accession number of actin.
REFERENCES
- Bustin, S.A., 2000. Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. J. Mol. Endocrinol., 25: 169-193.
CrossRefPubMedDirect Link - Caldana, C., W.R. Scheible, B. Mueller-Roeber and S. Ruzicic, 2007. A quantitative RT-PCR platform for high-throughput expression profiling of 2500 rice transcription factors. Plant Methods, 3: 7-7.
Direct Link - Czechowski, T., R.P. Bari, M. Stitt, W.R. Scheible and M.K. Udvardi, 2004. Real time RT-PCR profiling of over 1400 Arabidopsis transcription factors: Unprecedented sensitivity reveals novel root and shoot-specific genes. Plant J., 38: 366-379.
Direct Link - Dean, J.D., P.H. Goodwin and T. Hsiang, 2002. Comparison of relative RT-PCR and Northern blot analysis to measure expression of b-1, 3-glucanase in Nicotiana benthamiana infected with Colletotrichum destructivum. Plant Mol. Biol. Rep., 20: 347-356.
Direct Link - Duan, Y.H., Y.L. Zhang, L.T. Ye, X.R. TTTFan, G.H. Xu and Q.R. Shen, 2007. Responses of rice cultivars with different nitrogen use efficiency to partial nitrate nutrition. Ann. Bot., 99: 1153-1160.
Direct Link - Fan, C.C., Y.Z. Xing, H.L. Mao, T.T. Lu and B. Han et al., 2006. GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor. Appl. Genet., 112: 1164-1171.
CrossRefDirect Link - Fleige, S., V. Walf, S. Huch, C. Prgomet, J. Sehm and M.W. Pfaffl, 2006. Comparison of relative mRNA quantification models and the impact of RNA integrity in quantitative real-time RT-PCR. Biotechnol. Lett., 28: 1601-1613.
Direct Link - Hellemans, J., G. Mortier, A. De Paepe, F. Speleman and Vandesompele, 2007. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol., 8: R19-R19.
CrossRef - Hoogewijs, D., K. Houthoofd, F. Matthijssens, J. Vandesompele and J.R. Vanfleteren, 2008. Selection and validation of a set of reliable reference genes for quantitative sod gene expression analysis in C. elegans. BMC Mol. Biol., 9: 9-9.
Direct Link - Imin, N., T. Kerim, J.J. Weinman and B.G. Rolfe, 2006. Low temperature treatment at the young microspore stage induces protein changes in rice anthers. Mol. Cell. Proteomics, 5: 274-280.
Direct Link - Infante, C., M.P. Matsuoka, E. Asensio, J.P. Canavate, M. Reith and M. Manuel, 2008. Selection of housekeeping genes for gene expression studies in larvae from flatfish using real-time PCR. BMC Mol. Biol., 9: 28-28.
Direct Link - Iskandar, H.M., R.S. Simpson, R.E. Casu, G.D. Bonnett, J. Donald, D.J. Maclean and J.M. Manners, 2004. Comparison of reference genes for quantitative real-time polymerase chain reaction analysis of gene expression in sugarcane. Plant Mol. Biol. Rep., 22: 325-337.
Direct Link - Jain, M., A. Nijhawan, A.K. Tyagi and J.P. Khurana, 2006. Validation of housekeeping genes as internal control for studying gene expression in rice by quantitative real-time PCR. Biochem. Biophys. Res. Commun., 345: 646-651.
CrossRefPubMedDirect Link - Jain, M., A.K. Tyagi and J.P. Khurana, 2006. Genome-wide analysis, evolutionary expansion and expression of early auxin-responsive SAUR gene family in rice (Oryza sativa). Genomics, 88: 360-371.
Direct Link - Janovick-Guretzky, N.A., H.M. Dann, D.B. Carlson, M.R. Murphy, J.J. Loor and J.K. Drackley, 2007. Housekeeping gene expression in bovine liver is affected by physiological state, feed intake and dietary treatment. J. Dairy Sci., 90: 2246-2252.
Direct Link - Kim, B.R., H.Y. Nam, S.U. Kim, S.L. Kim and Y.J. Chang, 2003. Normalization of reverse transcription quantitative-PCR with housekeeping genes in rice. Biotechnol. Lett., 25: 1869-1872.
Direct Link - McElroy, D., M. Rothenberg, K.S. Reece and R. Wu, 1990. Characterization of the rice (Oryza sativa) actin gene family. Plant Mol. Biol., 15: 257-268.
Direct Link - Nicot, N., J.F. Hausman, L. Hoffmann and D. Evers, 2005. Housekeeping gene selection for real-time RT-PCR normalization in potato during biotic and abiotic stress. J. Exp. Bot., 56: 2907-2914.
CrossRefDirect Link - Reid, K.E., N. Olsson, J. Schlosser, F. Peng and S.T. Lund, 2006. An optimized grape vine RNA isolation procedure and statistical determination of reference genes for real-time RT-PCR during berry development. BMC Plant Biol., 6: 27-27.
CrossRefDirect Link - Singh, R. and M.R. Green, 1993. Sequence-specific binding of transfer RNA by glyceraldehyde-3-phosphate dehydrogenase. Science, 259: 365-368.
Direct Link - Song, X.J., W. Huang, M. Shi, M.Z. Zhu and H.X. Lin, 2007. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat. Genet., 39: 623-630.
CrossRefDirect Link - Thellin, O., W. Zorzi, B. Lakaye, B.D. Borman and B. Coumans et al., 1999. Housekeeping genes as internal standards: Use and limits. J. Biotechnol., 75: 291-295.
CrossRefDirect Link - Schmittgen, T.D, B.A. Zakrajsek, A.G. Mills, V. Gorn, M.R. Singer and M.W. Reed, 2000. Quantitative reverse transcription±polymerase chain reaction to study mrna decay: Comparison of endpoint and real-time methods. Anal. Biochem., 285: 194-204.
Direct Link - Toyota, K., M. Tamura, T. Ohdan and Y. Nakamura, 2006. Expression profiling of starch metabolism-related plastidic translocator genes in rice. Planta, 223: 248-257.
Direct Link - Tricarico, C., P. Pinzani, S. Bianchi, M. Paglierani and V. Distante et al., 2002. Quantitative real-time reverse transcription polymerase chain reaction: Normalization to rRNA or single housekeeping genes is inappropriate for human tissue biopsies. Anal. Biochem., 309: 293-300.
PubMedDirect Link - Tsuchiya, T., T. Takesawa, H. Kanzaki and I. Nakamura, 2004. Genomic structure and differential expression of two tandem-arranged GSTZ genes in rice. Genetic, 335: 141-149.
Direct Link - Tu, L.L., X.L. Zhang, D.Q. Liu, S.X. Jin and J.L. Cao et al., 2007. Suitable internal control genes for qRT-PCR normalization in cotton fiber development and somatic embryogenesis. Chinese Sci. Bull., 52: 3110-3117.
Direct Link - Vandesompele, J., K. De Preter, F. Pattyn, B. Poppe, N. van Roy, A. De Paepe and F. Speleman, 2002. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol., 3: 1-11.
CrossRefDirect Link - Wasaki, J., R. Yonetani, T. Shinano, M. Kai and M. Osaki, 2003. Expression of the OsPI1 gene, cloned from rice roots using cDNA microarray, rapidly responds to phosphorus status. New Phytologist., 158: 239-248.
Direct Link - Xue, W.Y., Y. Xing, X. Weng, Y. Zhao and W. Tang et al., 2008. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nat. Genet., 40: 761-767.
CrossRefDirect Link - Yu, S.W., Y. Liu and L.J. Luo, 2007. Analysis of relative gene expression using different real-time quantitative PCR. Acta Agronomica Sinica, 33: 1214-1218.
Direct Link - Zhao, S.P., X.Q. Zhao, S.M. Li and W.M. Shi, 2006. Optimization and application of real-time PCR method for detecting the expression levels of nitrogen assimilation-related genes in rice. Russ. J. Plant Physiol., 53: 560-569.
Direct Link