
P. Maneerattanarungroj
Applied Taxonomic Research Center, Department of Biology, Faculty of Science, Khon Kaen University, Khon Kaen 40002, Thailand
S. Bunnag
Applied Taxonomic Research Center, Department of Biology, Faculty of Science, Khon Kaen University, Khon Kaen 40002, Thailand
M. Monthatong
Applied Taxonomic Research Center, Department of Biology, Faculty of Science, Khon Kaen University, Khon Kaen 40002, Thailand
Asian Journal of Plant Sciences
Year: 2007 | Volume: 6 | Issue: 8 | Page No.: 1235-1240







ABSTRACT
Protocorm cryopreservation has been performed for ex situ preservation of Cleisostoma areitinum (Rchb.f.) Garay, rare Thai orchid species. Cryopreservation of protocorms was investigated by an encapsulation-dehydration method. The highest regrowth percentage of cryopreserved explants was observed when they were precultured on semi-solid ND medium with 0.25 M sucrose in the dark for 1 week then transferred to liquid ND medium containing 0.75 M sucrose for 2 days. The treated explants were dehydrated with silica gel in laminar air-flow for 5 h prior to immersion in liquid nitrogen for 1 h. Protocorm viability was tested by the TTC (2, 3, 5-triphenyltetrazoliumchloride) assay and regrowth ability was assessed by determining the survival percentage after recovery for 2 weeks. The survival rate of cryopreserved protocorms was 49% while percentage of TTC assay was 77%. Consequently, the simple and reliable protocol for cryopreservation of C. areitinum was clearly revealed in this study.
PDF Abstract XML References Citation
How to cite this article
P. Maneerattanarungroj, S. Bunnag and M. Monthatong, 2007. In vitro Conservation of Cleisostoma areitinum (Rchb.f.) Garay, Rare Thai Orchid Species by an Encapsulation-Dehydration Method. Asian Journal of Plant Sciences, 6: 1235-1240.
DOI: 10.3923/ajps.2007.1235.1240
URL: https://scialert.net/abstract/?doi=ajps.2007.1235.1240
DOI: 10.3923/ajps.2007.1235.1240
URL: https://scialert.net/abstract/?doi=ajps.2007.1235.1240
INTRODUCTION
Orchids are well known as important ornamental plants in Thailand, especially wild orchids. Cleisostoma arietinum is an epiphyte orchid and native to Thailand. It is classified as an endangered species that should be conserved (Kew, 1999). According to in situ preservation, to conserve plant species only in field is also risky, as valuable germplasm can be lost (genetic erosion) because of pets, diseases and adverse weather conditions. Moreover, the maintenance of clonal orchids is labour-intensive and expensive. The maintenance of in vitro collections (established for some vegetatively propagated species) is also labour-intensive and there is the risk of losing accessions due to contamination, human errors or somaclonal variations (i.e., mutations that occur spontaneously in tissue culture, with a frequency that increases with repeated sub-culturing) (Panis and Lambardi, 2005). Thus, cryopreservation has attracted attention as a method for storage of variable plant germplasm (Hirano et al., 2005). Cryopreservation involves the storage of live plant cells, tissues or organs at ultra-low temperature (-196°C, liquid nitrogen) with a supposed low risk of genetic and physiological changes even over long periods of time (Gonzalez-Benito and Perez, 1997). It is particularly useful for the conservation of precious and rare species due to global environment problems (Hirata et al., 1998). This effective method also offers a safe and cost-effective option for a long-term conservation of genetic resources in many plant species; for example, Vanda pumila (Na and Kondo, 1996), Bletilla striata (Ishikawa et al., 1997; Hirano et al., 2005), Geodorum densiflorum (Datta et al., 1999), Doritaenopsis orchid (Tsukazaki et al., 2000), Dendrobium orchid (Dendrobium Walter Oumae) (Lurswijidjarus and Thammasiri, 2004), sweet potato (Ipomoea batatas [L.] Lam.) (Bhatti et al., 1997), horseradish (Armoracia rusticana) (Phuchindawan et al., 1997) and Gentian (Gentiana scabra Bunge var. buergeri Maxim.) (Suzuki et al., 2005).
There are many different techniques of cryopreservation used to develop cryogenic protocols. The encapsulation-dehydration method is our selected protocol in this research. It is easy to handle, avoids the use of costly programmable freezers and uses sucrose as the only cryoprotectant (Martinez et al., 1999). Moreover, encapsulation of explants in alginate beads for cryopreservation has some benefit compared with the use of non-encapsulated samples. The alginate beads provide enhanced protection of dried materials from mechanical and oxidative stress during storage and ease the handling of small samples during pre- and post-cryopreservation procedures (Niino and Sakai, 1992). Encapsulation-dehydration technique have been used in the successful cryopreservation of Melia azedarach (Scocchi et al., 2004), Centaurium rigualii (Gonzalez-Benito and Perez, 1997) and Hop (Dumet et al., 2002).
This study reports on experiments undertaken on encapsulated protocorms to examine the effects of dehydration time on water content, percentage of electrolyte leakage, percentage of TTC staining and survival rate of cryopreserved explants. This study provides the first report of a simple and reliable encapsulation-dehydration method for the cryopreservation of C. areitinum protocorms.
MATERIALS AND METHODS
Plant materials: The capsules of C. areitinum were surface sterilized in 70% (v/v) ethanol for 5 min and then shaken in 25% (v/v) sodium hypochlorite for 20 min prior to washing 3 times in sterile distilled water. The capsules were then aseptically cut open and seeds transferred to New Dogashima (ND) medium (Tokuhara and Mii, 1993) which containing 2 mg L-1 BA and 2 mg L-1 NAA with 1% (w/v) sucrose and 0.85% (w/v) agar (pH 5.4). Seeds were cultured at 25±2°C under a photoperiod of 16 h light/ 8 h dark with a light intensity at 40 μmol m-2 sec-1. Protocorms reached an average diameter of 1-3 mm were used as explants in all experiments.
Encapsulation, preculture and desiccation: Protocorms were immersed in a 3% (w/v) alginate solution prepared with ND medium free of calcium, growth regulator and iron. Each protocorm was dropped into 100 mM CaCl2 solution and allowed to polymerize for 10 min. The resulting beads (3-5 mm in diameter) were washed in sterile distilled water for 3 times. The encapsulated protocorms were precultured in a semi-solid ND medium containing 0.25 M sucrose in the dark for 1 week at 4°C then transferred into liquid ND medium containing 0.75 M sucrose for 2 days. Following osmotic desiccation, encapsulated protocorms were surface dried on filter paper in Petri dishes (30 beads/Petri dish containing 20 g of silica gel) and dried in a laminar air-flow (Fig. 1). Various desiccation periods from 0 to 6 h were tested. There were 10 encapsulated protocorms for each treatment and the experiments were repeated three times.
Freezing and thawing: The desiccated beads were transferred to 1.8 mL cryotubes (ten encapsulated protocorms per cryotube) and directly immersed into liquid nitrogen for 1 h in the dark. Cryotubes were then warmed rapidly in a water bath at 38±2°C for 2 min and cryopreserved explants were transferred to the establishment medium; ND medium supplemented with 2 mg L-1 BA, 2 mg L-1 NAA, 1% (w/v) sucrose and 0.85% (w/v) agar (pH 5.4).
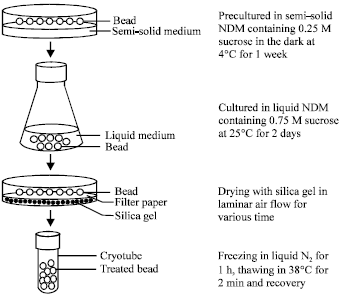 | |
| Fig. 1: | Preculture pattern of encapsulated protocorms |
Survival rates, water content and statistical analysis: The cryopreserved protocorms were cultured in liquid ND medium supplemented with 1% (w/v) sucrose, 1% (w/v) potato extract, 2 mg L-1 BA and 2 mg L-1 NAA. Survival rates were determined by regrowth of cryopreserved protocorms about 2 weeks after recovery from liquid nitrogen. Protocorms were recorded as viable if they showed signs of growth (greening and expansion). Water content (in grams of water per gram of dry weight [g H2O g-1 DW]) was determined from the weight loss of beads when dried in an oven at 60°C for 24 h (Dumet et al., 2002). Experiments were replicated three times with at least ten samples for all treatments. Analysis of variance and mean comparison were conducted. The comparison of means was performed by LSD test.
Viability test by TTC staining: Cryopreserved explants were placed in test tubes and 500 μL of TTC (2,3,5 Triphenyltetrazoliumchloride) solution then added prior to incubation in the dark at 25±2°C for 24 h. The living plant cells should show red colour because dehydrogenase enzyme in living plant cells reduced the colorless triphenyltetrazoliumchloride to triphenylformazan (reduced TTC) (Fokar et al., 1998).
Electrolyte leakage (EL): After dehydrated, freezing and thawing, plant samples were transferred in test tube which containing 10 mL of deionized water. The tissue samples were incubated at 32°C for 2 h and initial electrical conductivity of the medium (EC1) was measured using an electrical conductivity meter (WTW TetraCon®325, inoLab pH/Cond Level 1). After EC1 measurements were made on all treatments, all samples were autoclaved for 20 min (121°C, 1.06 kg cm-2) to completely kill the tissue and release all electrolytes. The final electrical conductivity of the medium (EC2) was measured after cooled sample in a water bath at 25°C. Freeze damage of the tissue was calculated as a percentage of total electrolyte leakage (EL) (EL = EC1/EC2x100).
RESULTS AND DISCUSSION
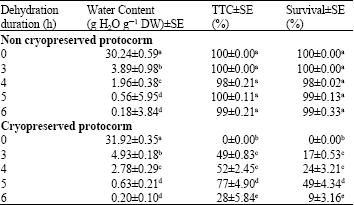
Water content: Water content of treated explants in all condition was dramatically decreased when drying in laminar flow cabinet for 3 to 6 h. Drying with silica gel, water content of beads were significantly lower than water content of dehydrated bead that drying alone without silica gel (p<0.05). This result indicated that drying with silica gel enhance effect on desiccation of plant sample before freezing (Table 1).
According to the result, water content of dehydrated beads was depended on the desiccation period. Most of encapsulated protocorms that contained high water content were dead after freezing in liquid nitrogen. In contrast, beads with the lowest water contents showed the highest levels of viability (approximately 0.63 g H2O g-1 DW, Table 1). Water content of treated explant was also important for survival rate after recovery from liquid nitrogen. Bian et al. (2002) reported that the optimal water content of protocorm-like bodies (PLBs) of Dendrobium candidum before freezing seemed to be at the range of 0.1 -0.5 g H2O g-1 DW. However, optimal water content for cryopreserved explants was depended on many conditions such as plant species, developmental stage, preculture regime and recovery protocol.
In fact, water is the important driving force in cellular organization; the assembly of phospholipids into biological membranes and, in part, for the conformation of many proteins. If water completely dissipates from living matter, the driving force for cellular organization is lost. Membranes then undergo structural changes and proteins denature (Hoekstra et al., 2001). However, in cryopreservation, high water contents in normal cells can produce intracellular ice crystals during freezing and subsequent thawing (Bian et al., 2002; Hoekstra et al., 2001; Popov et al., 2006).
It is concluded that desiccation encapsulated protocorms in laminar flow cabinet with silica gel for 5 h is the optimal dehydration duration for avoid water content before freezing these plant samples.
| Table 1: | Comparison of water content of dehydrated samples with and without silica gel after preculture and drying for various time |
 | |
| Means with the same letter(s) are not significantly different at p<0.05 (LSD test) | |
| Table 2: | Comparison of water content, percentage of TTC assay after recovery from liquid nitrogen and survival rate of cryopreserved protocorms after regrowth for 2 weeks |
 | |
| Means with the same letter(s) are not significantly different at p<0.05 (LSD test) | |
Survival rate and TTC assay: We studied the effect of dehydration time on protocorm recovery after the beads had been immersed in liquid nitrogen. The encapsulated protocorms were precultured in semi-solid ND medium in the dark for 1 week and then transferred to liquid ND medium with 0.75 M sucrose for 2 days. Five different dehydration periods (0 to 6 h) were assessed. The best results in terms of protocorm recovery were obtained with 5 h dehydration (Table 2). Appreciably lower recovery levels were observed with 3, 4 and 6 h dehydration periods and no protocorms recovered when there was no dehydration period.
In this report, protocorms were dehydrated using sucrose and air-drying. Many reports have shown the effect of sucrose on osmotic dehydration in plant species (Bian et al., 2002; Hoekstra et al., 2001; Popov et al., 2006). Following membrane protection mechanism, sucrose fulfills two important roles in protecting the membranes of the dry organism: reduction of the dry membrane phase transition temperature (Tm) and formation of a carbohydrate glass with a high melting temperature (Oliver et al., 2002). Moreover, Buitink and Leprince (2004) also found that the formation of a glass during the dehydration step prevented serious damage to cells.
 | |
| Fig. 2a-h: | Plant samples of C. areitinum; fruit stage (a), protocorm clump (b) scale = 5 mm, protocorms used in this study (c) scale = 5 mm, Plantlets derived from non cryopreserved protocorms (d) scale = 5 mm, encapsulated protocorms (e) scale = 5 mm, TTC assay of non treated bead, control (left) and treated bead (right) (f) scale = 1 mm, cryopreserved plantlets after recovery for 3 months and (g-h) scale = 5 mm, plantlets derived from non cryopreserved and cryopreserved protocorms after regrowth in ND medium containing 2 mg L-1 NAA and 2 mg L-1 BA for 3 months scale = 5 mm |
Some types of sugar can protect cells or tissues from freezing damage. The high viscosity of sugar solutions inhibits chemical reactions within the cells. Although vitrification during freezing is a key factor for cryopreservation but in the present work we did not measure glass formation and thus do not know whether the water in the protocorms entered into the glass state.
In our experiment, viability tests were investigated by the TTC assay and regrowth ability after 2 weeks of treatments. The highest regrowth was observed when encapsulated protocorms were precultured in semi-solid ND medium containing 0.25 M sucrose in darkness for 1 week and then transferred to liquid ND medium with 0.75 M sucrose for 2 days and subsequently desiccated for 5 h. The survival rate was up to 49 %.
Following this result, the regrowth ability of cryopreserved protocorms appeared lower than the viability test by TTC assay (TTC percentage) (Table 2). After recovery from liquid nitrogen, some explants were still living, but others were damaged and finally dead. Therefore, some cryopreserved protocorms showed a brown color after testing with the TTC solution. This brown colour may be the source of confusion due to its similarity to the red colour (reduced TTC). This phenomenon probably occurred because of damage caused by oxidative reactions in the freezing step (Verleysen et al., 2004). However, TTC staining allowed us to assess the viability of the explants in this experiment, TTC values after thawing provide a guide to predict the success of experiments (Florin et al., 2000). For regeneration, plantlets were developed 3 months after cryopreservation (Fig. 2). There was no morphological difference between plantlets from non-cryopreserved protocorms and cryopreserved protocorms. Nevertheless, the recovery rate of cryopreserved protocorms after freezing in liquid nitrogen in this experiment was poor and slow when comparing with non-cryopreserved protocorms. This may be caused by non-optimised conditions such as light intensity or culture medium. Grout (1995) also found that a high level of light might cause photobleaching with adverse effects on viability during early regrowth of thawed explants.
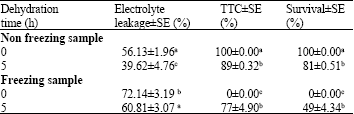
| Table 3: | Electrolyte leakage percentage, percentage of TTC assay and survival of cryopreserved samples when dehydrated for 0 and 5 h |
 | |
| Means with the same letter(s) are not significantly different at p<0.05 (LSD test) | |
Therefore, it may be helpful to reduce this non-optimised condition.
Electrolyte leakage: Test in dehydration time between 0 and 5 h had a significant (p<0.05) effect on EL. For dehydrated explants, EL was 39.63±4.76% (non-freezing) and 60.81±3.07% (freezing) whereas non-dehydrated explants showed high level of EL, 53.13±1.96% (non-freezing) and 72.14±3.19% (freezing). This result indicated that dehydrated explant showed less electrolyte leakage than the control explant (Table 3). The increases in EL with cold storage of dehydrated beads indicate that the chilling requirement was supplied in storage, resulting in cold hardiness with continued exposure. During dehydration, cold storage and thawing, cell membrane also sensitive to these stresses. Damaged membranes allow electrolyte from injured cells to flow into surrounding tissue (Wilson and Jacobs, 2004). The electrolyte leakage assay is widely used to evaluate the degree of cold injury in many plant species (Campos et al., 2003; Mirdehghan et al., 2007; Plazek and Zur, 2003; Wang and Li, 2006). Although dehydration treatment increased freezing tolerance but dehydration duration and optimal concentration of sucrose also important to preserve all of explants. It is concluded that EL levels in treated explants could indicate that dehydrate and freezer storage regimes are effective in maintaining a lower degree of physiological activity for this orchid species.
CONCLUSIONS
In conclusion, preservation of C. areitinum protocorms is possible through a cryogenic procedure after preculture with 0.25 and 0.75 M sucrose and drying in laminar air flow with silica gel for 5 h. However, more experiment is needed to optimize high survival rates of protocorms after recovery from liquid nitrogen, especially the effect of sucrose concentration on water content and protocorm recovery of cryopreserved explants. Moreover, DNA analysis or genetic fidelity of non-cryopreserved and cryopreserved explant will be studied in the future. This study will provide useful information for planning and implementing related projects in the future. Moreover, it may be useful for other plant species, although other cell lines will need special preliminary studied on cultivation conditions, cold acclimation, specific nutrients and other supplementation.
ACKNOWLEDGMENTS
This study was partly supported by Grant No. ATRC_R4902 from Applied Taxonomic Research Center (ATRC) and Graduate School, Khon Kaen University, Thailand. Authors are grateful to Dr. Bernard A. Goodman for reviewing the manuscript.
REFERENCES
- Bhatti, H.M., T. Percival, M.D.C. Davey, G.C. Henshaw and D. Blakesley, 1997. Cryopreservation of embryogenic tissue of a range of genotypes of sweet potato (Ipomoea batatas [L.] Lam.) using an encapsulation protocol. Plant Cell Rep., 16: 802-806.
Direct Link - Bian, H.W., J.H. Wang, W.Q. Lin, N. Han and M.Y. Zhu, 2002. Accumulation of soluble sugars, heat-stable proteins and dehydrins in cryopreservation of protocorm-like bodies of Dendrobium candidum by air-drying method. J. Plant Physiol., 159: 1139-1145.
Direct Link - Buitink, J. and O. Leprince, 2004. Glass formation in plant anhydrobiotes: Survival in the dry state. Cryobiology, 48: 215-228.
CrossRefDirect Link - Campos, P.S., V. Quartin, J. Ramalho and M.A. Nunes, 2003. Electrolyte leakage and lipid degradation account for cold sensitivity in leaves of Coffea sp. plants. J. Plant Physiol., 160: 283-292.
Direct Link - Dumet, D., A. Grapin, C. Bailly and N. Dorion, 2002. Revisiting crucial steps of an encapsulation/ desiccation based cryopreservation process: Importance of thawing method in the case of Pelargonium meristems. Plant Sci., 163: 1121-1127.
Direct Link - Florin, B., E. Brulard, P.J. Ducos, H. Tessereau and V. Petiard, 2000. Development of a Simplified Method for the Routine Cryopreservation of Coffee Germplasm Collection. In: Cryopreservation of Tropical Plant Germplasm, Engelmann, F. and T. Hiroko (Eds.). Japan/International Plant Genetic Resources Institute, Rome, Italy, pp: 496.
- Fokar, M., T.H. Nguyen and A. Blum, 1998. Heat tolerance in spring wheat. I. estimating cellular thermotolerance and its heritability. Euphytica, 104: 1-8.
CrossRef - Hirano, T., T. Godo, M. Mii and K. Ishikawa, 2005. Cryopreservation of immature seeds of Bletilla striata by vitrification. Plant Cell Rep., 23: 534-539.
CrossRefDirect Link - Hirata, K., S. Goda, M. Phunchindawan, D. Du, M. Ishio, A. Sakai and K. Miyamoto, 1998. Cryopreservation of horseradish hairy root cultures by encapsulation-dehydration. J. Ferment. Bioeng., 86: 418-420.
Direct Link - Hoekstra, F.A., E.A. Golovina and J. Buitink, 2001. Mechanisms of plant desiccation tolerance. Trends Plant Sci., 6: 431-438.
CrossRefPubMedDirect Link - Ishikawa, K., K. Harata, M. Mii, A. Sakai, K. Yoshimatsu and K. Shimomura, 1997. Cryopreservation of a Japanese terrestrial orchid (Bletilla striata) by vitrification. Plant Cell Rep., 16: 754-757.
Direct Link - Lurswijidjarus, W. and K. Thammasiri, 2004. Cryopreservation of shoot tips of Dendrobium walter oumae by encapsulation/dehydration. Sci. Asia, 30: 293-299.
Direct Link - Martinez, D., S.R. Tames and A.M. Revilla, 1999. Cryopreservation of in vitro grown shoot-tips of hop (Humulus lupulus L.) using encapsulation/ dehydration. Plant Cell Rep., 19: 59-63.
Direct Link - Mirdehghan, H.S., M. Rahemi, S. Castillo, D. Martinez-Romero, S. Maria and Valero, 2007. Pre-storage application of polyamines by pressure or immersion improves shelf-life of pomegranate stored at chilling temperature by increasing endogenous polyamine levels. Posthar. Biol. Technol., 44: 26-33.
Direct Link - Na, Y.H. and K. Kondo, 1996. Cryopreservation of tissue-cultured shoot primordia from shoot apices of cultured protocorms in Vanda pumila following ABA preculture and desiccation. Plant Sci., 118: 195-201.
CrossRefDirect Link - Nino, T. and A. Sakai, 1992. A Cryopreservation of alginate-coated in vitro grown shoot tips of apple, pear and mulberry. Plant Sci., 87: 199-206.
Direct Link - Oliver, E.A., K.D. Hincha and H.J. Crowe, 2002. Looking beyond sugars: The role of amphiphilic solutes in preventing adventitious reactions in anhydrobiotes at low water contents. Comp. Biochem. Physiol. Part A, Mol. Integrative Physiol., 131: 515-525.
Direct Link - Plazek, A. and I. Zur, 2003. Cold-induced plant resistance to necrotrophic pathogens and antioxidant enzyme activities and cell membrane permeability. Plant Sci., 164: 1019-1028.
Direct Link - Popov, S.A., V.E. Popova, V.T. Nikishina and N.O. Vysotskaya, 2006. Cryobank of plant genetic resources in Russian academy of sciences. Int. J. Refrigeration, 29: 403-410.
Direct Link - Scocchi, A., M. Faloci, R. Medina, S. Olmos and L. Mroginski, 2004. Plant recovery of cryopreserved apical meristem-tips of Melia azedarach L. using encapsulation/dehydration and assessment of their genetic stability. Euphytica, 135: 29-38.
Direct Link - Suzuki, M., T. Akihama and M. Ishikawa, 2005. Cryopreservation of encapsulated gentian axillary buds following 2 step-preculture with sucrose and desiccation. Plant Cell Tissue Organ Cult., 83: 115-121.
CrossRef - Tokuhara, K. and M. Mii, 1993. Micropropagation of phalaenopsis and doritaenopsis by culturing shoot tips of flower stalk buds. Plant Cell Rep., 13: 7-11.
Direct Link - Tsukazaki, H., M. Mii, K. Tokuhara and K. Ishikawa, 2000. Cryopreservation of Doritaenopsis suspension culture by vitrification. Plant Cell Rep., 19: 1160-1164.
Direct Link - Verleysen, H., G. Samyn, E. Van Bockstaele and P. Debergh, 2004. Evaluation of analytical techniques to predict viability after cryopreservation. Plant Cell Tissue Organ Cult., 77: 11-21.
Direct Link - Wang, L.J. and S.H. Li, 2006. Salicylic acid-induced heat or cold tolerance in relation to Ca2+ homeostasis and antioxidant systems in young grape plants. Plant Sci., 170: 685-694.
Direct Link