
Hala Abdel Wahab
Laboratory of Molecular Diagnostic of Plant Diseases, Department of Plant Pathology, Faculty of Agriculture, Ain Shams University, Cairo, Egypt
LiveDNA: 20.16685
Elsayed E. Wagih
Department of Plant Pathology, College of Agriculture, University of Alexandria, El-Shatby, Alexandria, Egypt
LiveDNA: 20.524
Mohamed R.A. Shehata
Department of Plant Pathology, College of Agriculture, University of Alexandria, El-Shatby, Alexandria, Egypt
Magda M. Fahmy
Department of Plant Pathology, College of Agriculture, University of Alexandria, El-Shatby, Alexandria, Egypt
Mahmoud A. Gaber
Department of Plant Pathology, College of Agriculture, University of Alexandria, El-Shatby, Alexandria, Egypt
Asian Journal of Plant Pathology
Year: 2020 | Volume: 14 | Issue: 1 | Page No.: 1-10







ABSTRACT
Background and Objective: Botrytis is a serious fungal pathogen which infects most ornamental plants and its resistance to effective fungicide is a continuous challenge facing disease management. The current research aimed to identify resistant Botrytis spp. from different ornamental plants against fenhexamid. Materials and Methods: About 64 single-spore isolates were collected from different symptomatic and asymptomatic ornamental plants using the modified Kerssies medium, m1KERS. Phenotypic variability was shown in some features including growth texture, sclerotia pattern and conidial dimensions. Pathogenicity and mycelial growth rate tests were statistically carried out using one way analysis of variance (ANOVA). Molecular characterisation was demonstrated by transposable elements (TEs) analysis and fenhexamid resistance was revealed using mycelial growth assay. Results: All isolates were identified as Botrytis spp. according to their morphological and molecular characteristics. Based on phylogenetic analysis, BCLi1 isolate seemed to be distantly related to B. cinerea and other Botrytis species. Significant variation in aggressiveness among isolates was observed on detached lettuce leaves. Four TE genotypes, transposa, boty, flipper and vacuma, were detected in Botrytis populations with TE distribution reached 50, 29.7, 10.9 and 9.4%, respectively. The low virulence level of some Botrytis spp. isolates seemed to be related to vacuma isolates suggesting a possible correlation between virulence and TE genotype. Moreover, a low to moderate resistance of Botrytis spp. isolates was revealed towards the hydroxyanilide fungicide, fenhexamid, in three transposa isolates, BCT6, BCGL2 and BCGL5, with a mean EC50 value of 0.2, 0.3 and 0.5 μg mL–1, respectively. Conclusion: The results demonstrated the first observation of the emergence of resistant isolates of Botrytis from ornamental plants against the highly effective fungicide in Egypt, providing a serious attention towards Botrytis management.
PDF Abstract XML References Citation
Received: August 25, 2019;
Accepted: November 18, 2019;
Published: December 15, 2019
How to cite this article
Hala Abdel Wahab, Elsayed E. Wagih, Mohamed R.A. Shehata, Magda M. Fahmy and Mahmoud A. Gaber, 2020. Studies on Botrytis spp. Infected Ornamental Plants and Emergence of Resistant Isolates Against Fenhexamid in Egypt. Asian Journal of Plant Pathology, 14: 1-10.
URL: https://scialert.net/abstract/?doi=ajppaj.2020.1.10
URL: https://scialert.net/abstract/?doi=ajppaj.2020.1.10
INTRODUCTION
Botrytis spp. and B. cinerea Pers.: Fr. (teleomorph, Botryotinia fuckeliana (de Bary) Whetzel) are necrotrophic pathogens that attack flowers at any stage, especially new tender and senescent tissues which are exceptionally susceptible1 at any time proper conditions prevail in greenhouses, fields or during shipping. B. cinerea has the widest host range and can infect more than 1000 plant species2. This pathogen is found worldwide and causes disease in many fruit, flower and leafy crops1,3. One of the molecular variation factors implicated in fungal genomic development is the transposable elements (TEs) which were characterized by their ability to transpose within the genome resulting in a huge impact on genome function and evolution4,5. Based on the presence of TEs, flipper and boty, Botrytis sp. isolates have been classified to four TEs genotypes, namely, transposa, boty, flipper and vacuma6-8. For Botrytis management, growers use many fungicides such as fenhexamid (a non-systemic fungicide with both protective and curative activity), chlorothalonil and iprodione to which Botrytis spp. strains have shown various degrees of resistance. Previously, no resistant isolate of Botrytis to the hydroxyanilide fungicide, fenhexamid was observed on various plants in Egypt6,8. As Botrytis spp. attack a broad range of plant species in various geographic conditions, we should study different characterization aspects especially on those isolated from ornamental plants which were not, in our knowledge, well studied in Egypt. Additionally, Botrytis spp. known to make adaptation towards many fungicides including fenhexamid as it was highly efficient, leading to emergence of new resistant isolates. Naturel resistance to fenhexamid was also evident in Botrytis isolates due to their genetic plasticity. Despite this fact, low reduction of fenhexamid efficacy had been recorded in various countries for many years after its registration. At the same time, the other toxic fungicides, such as dicarboximides are nearly banned in many countries as high frequency of resistant Botrytis isolates had arisen3. In addition, a widespread resistance to the benzimidazole fungicides has been reported9 and therefore they are no longer suggested for use in Botrytis management. The aims of the present investigation were to identify and characterise Botrytis spp. isolated from symptomatic and asymptomatic ornamental plants and to determine the baseline sensitivity of Botrytis isolates to fenhexamid.
MATERIALS AND METHODS
Plant and fungal sample collection: Samples from ornamental plants were collected from rose (Rose sp.), begonia (Begonia rex), narcissus (Narcissus tazetta), gladiolus (Gladiolus communis), lily (Lilium sp.) and tulip (Tulipa gesneriana) grown in different Egyptian governorates (Table 1) during 2015-2016. These samples were either symptomatic, showing typical symptoms of Botrytis blight or asymptomatic, with no symptoms whatsoever. The symptomatic and asymptomatic plant samples were cut into small pieces (0.5 cm), dipped separately in sterilised water for 5 min, dried on sterilised filter paper, then plated onto the modified Kerssies medium, m1KERS10 and incubated at 23°C for 3-21days.
Fungal identification: Single spore of each isolate was grown on a PDA medium and the resulting culture was preserved under sterilised paraffin oil at 4°C. Botrytis isolates were identified based on morphological and cultural characteristics according to previous studies6,8.
Molecular identification of Botrytis isolates using PCR: Genomic DNA was extracted and purified from harvested mycelia using the mini preparation procedure according to Moller et al.11. Botrytis identification was carried out using specific primers12 (Table 2) and then DNA sequence was revealed using ITS1/ITS4 primers13 (Table 2). The PCR programme was performed in a total volume of 25 μL consisting of 2 μL genomic DNA (50 ng μL–1), 0.5 μL of each primer "Bio-search Technologies" (10 μM), 12.5 μL Red PCR master mix (Bio-line) and 9.5 μL H2O. Amplification was conducted in a thermocycler (Techne-Progene) as follows: Initial denaturation at 94°C for 4 min, followed by 35 cycles of denaturation at 94°C for 1 min, annealing at 55-64°C for 1 min, extension at 72°C for 1 min and a final extension step at 72°C for 10 min.
Evolutionary analysis by maximum likelihood method: The maximum likelihood method and Tamura-Nei model were used according to Tamura and Nei14 to produce evolutionary analyses which were conducted by MEGA X15. From 1000 replicates, a bootstrap consensus tree was produced to represent the evolutionary taxa analysed16. Initial tree consisting of 21 nucleotide sequences was obtained using Neighbor-Join and BIONJ algorithms.
| Table 1: | Collection of Botrytis isolates from symptomatic and asymptomatic ornamental plants |
 | |
| Table 2: | Molecular identification and characterisation of Botrytis isolates collected from ornamental plants using different primers |
 | |
Detection of transposable elements (TEs): The PCR amplification of the two transposable elements, boty and flipper, was done using their specific primers17,18 (Table 2). The PCR preparation and programme were performed according to Abdel Wahab6.
Pathogenicity assay using detached lettuce leaves: Pathogenicity of Botrytis isolates was carried out using the detached lettuce (Lactuca sativa L. cv. ‘Baladi’) leaf technique6-8. Isolate aggressiveness was determined by measuring the lesion diameter of each inoculated leaf.
Determination of EC50 of fenhexamid for Botrytis isolates: Thirty isolates of Botrytis collected from the current ornamental samples: BCN1, BCN2, BCN4, BCLi1, BCLi2, BCLi3, BCLi4, BCGL1, BCGL2, BCGL3, BCGL4, BCGL5, BCGL6, BCGL7, BCGL8, BCGL9, BCT1, BCT2, BCT3, BCT4, BCT5, BCT6, BCT7, BCT8, BCT9, BCT10, BCT11, BCT12, BCT13 and BCT14 were tested for their sensitivity to the hydroxyanilide fungicide, fenhexamid (Teldor SC, 500 g L–1, Bayer) using PDA medium amended with 2 concentrations of fenhexamid (0.1 and 1 μg mL–1). Each isolate was represented by 3 replicate plates and the entire experiment was conducted 2 times. The diameters of the resulting colonies were measured in 2 perpendicular directions after 3 days of incubation at 23°C. Isolates that were able to grow on PDA containing >0.2 μg mL–1 fenhexamid were putatively considered fenhexamid resistant, while those that became inhibited were classified as fenhexamid sensitive. The 50% effective concentration (EC50), 50% mycelial growth inhibition, should be determined by regressing the relative growth rate (% control) against the log of the fungicide concentration.
Statistical analysis: Results were statistically tested using one way analysis of variance (ANOVA) to determine the significant difference. Data means were compared at p = 0.05 level using the least significant difference test.
RESULTS
Characterisation of Botrytis isolates collected from ornamental plants: The formation of brown halos was observed around many Botrytis spp. colonies resulting from symptomatic and asymptomatic ornamental plant samples after 3 days of incubation at 23°C using m1KERS selective media indicating their infection with Botrytis spp. The fungal colonies which grew on such selective medium were sub-cultured on PDA (Fig. 1) and then identified microscopically as Botrytis. Data presented in Table 3 show the isolate characterisation after collection from rose, tulip, gladiolus, begonia, narcissus and lily. The results demonstrated that Botrytis isolates varied in growth texture which was categorised as follows: compact, heavy warty, light warty, watery and fluffy (Table 3). The conidial dimensions differed among isolates and ranged from 10.6-12.9 μm in length, 7.1-8.9 μm in width and 276.9-539.4 μm3 in volume (Table 3). Conidia of all isolates were described as oval shape.
| Table 3: | Phenotypic characteristics of Botrytis spp. isolates collected from ornamental plants |
 | |
*Data are means of triplicate measurements±standard deviation (SD) at LSD 0.05, **All data are means of 20 conidial measurements±standard deviation (SD) at LSD 0.05 | |
 | |
| Fig. 1(a-c): | Cultural characteristics of Botrytis isolates collected from ornamental plants (a) BCLi4, (b) BCN4 and (c) BCGL7 |
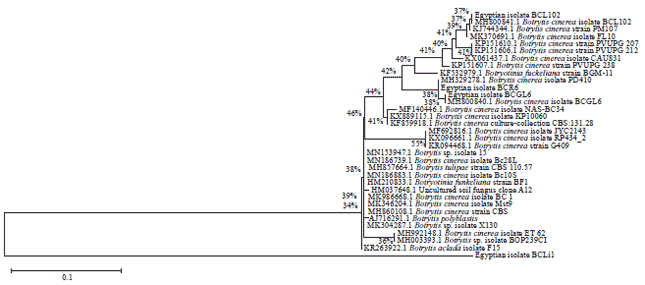 | |
| Fig. 2: | Phylogenetic tree of the tested sequences based on maximum likelihood method, bootstrap support values are written for each clade |
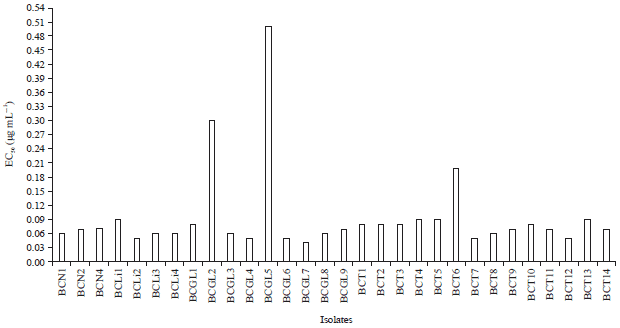 | |
| Fig. 3: | Resistance of some Botrytis spp. isolates obtained from different ornamental plants to fenhexamid, expressed as EC50 (μg mL–1) |
| Table 4: | Pathological, growth rate and TE patterns of Botrytis spp. collected from ornamental plants |
 | |
*MGR: Mycelial growth rate expressed as means of triplicate measurements±standard deviation (SD) at LSD 0.05, **data for lesion diameter were means of triplicate measurements±standard deviation (SD) at LSD 0.05 after 72 h of infection, ***Transposable element type, B: Boty, F: Flipper, T: Transposa, V: Vacuma, means followed by the same letter are not significantly different (p = 0.05) | |
The sclerotia which were cerebriform in all tested isolates showed varied characteristics in their number which ranged from 12.7-196.7 sclerotia/plate (9 cm).
Molecular identification of Botrytis spp.: PCR produced a single specific band of 700 bp long (data not shown) using a pair of specific primers corresponding to the expected amplicon length. Sequence analysis of the amplified product revealed that the three isolates tested, BCR6, BCLi1 and BCGL6 were Botrytis spp. based on the BLAST identity.
Pathological, MGR and TE patterns of Botrytis spp.: Virulence of 64 isolates of Botrytis spp. collected from the ornamental plants subjected to the present study was tested on detached lettuce leaves. Isolates showed significant differences in their aggressiveness (Table 4). Lesion diameter ranged from 0.2-4.7 cm and could be divided into 3 categories, namely highly, moderately and low virulent with a lesion diameter >2.5, >1-2.5 and 0.2-1 cm, respectively (Table 4). As to mycelial growth rate, it ranged from 0.3-4.2 cm/day (Table 4). TE typing of Botrytis isolates used in this study was achieved using the specific primers quoted in the material and methods section for separately amplifying boty and flipper elements and four TEs genotypes could be recognised (Table 4). PCR generated 510 bp and/or 1250 bp long products for boty and flipper elements, respectively. About 32 out of the 64 isolates tested demonstrated the co-presence of the two TEs, boty and flipper (transposa type). While, 19 isolates showed the presence of only boty element (boty type), 7 revealed the presence of only flipper element (flipper type) and 6 have neither boty nor flipper element (vacuma type) (Table 4).
Phylogenetic analysis of Botrytis spp. isolates: The result of the amplified region (ITS1-5.8s-ITS4) revealed that, the tested isolates were Botrytis spp. according to the percentage of sequence identity imported from GenBank using BLAST search. Considerable variation was observed using a multiple sequence alignment of amplified region. Phylogenetic tree branches which corresponded to partitions produced in less than 50% bootstrap replicates were collapsed. The replicate tree percentage was put next to tree branches in which taxa clustered together in the bootstrap test (1000 replicates)16. Sequence analysis of the amplified products revealed that the isolate BCLi1 was clearly distinguished from other Botrytis isolates deposited in the GenBank and those under study (Fig. 2).
Fenhexamid resistance of some Botrytis spp. isolates: Thirty isolates under study: BCN1, BCN2, BCN4, BCLi1, BCLi2, BCLi3, BCLi4, BCGL1, BCGL2, BCGL3, BCGL4, BCGL5, BCGL6, BCGL7, BCGL8, BCGL9, BCT1, BCT2, BCT3, BCT4, BCT5, BCT6, BCT7, BCT8, BCT9, BCT10, BCT11, BCT12, BCT13 and BCT14 were tested for resistance to fenhexamid. The EC50 of all sensitive isolates was <1 μg mL–1. The EC50 values ranged from 0.06-0.07, 0.05-0.09, 0.05-0.2 and 0.04-0.5 μg fenhexamid mL–1 for BCN, BCLi, BCT and BCGL isolates, respectively (Fig. 3). The highest sensitive isolates were BCGL7, BCLi2 and BCT7, having had EC50 values at 0.04, 0.05 and 0.05 μg fenhexamid mL–1, respectively and the highest resistant isolates were BCGL5, BCGL2 and BCT6, having expressed EC50 value at 0.5, 0.3 and 0.2 μg fenhexamid mL–1, respectively (Fig. 3).
DISCUSSION
The current study provides informative results about the morphological, cultural, molecular and fungicide-resistance features of Botrytis spp. infecting a number of ornamental plants in Egypt. The results showed a phenotypic and pathological diversity among isolates regardless their host plant and location as previously reported19. Similar observations had been documented in numerous isolates obtained from a number of host plants from California20 and Tunisia21. All conidial and sclerotial morphology demonstrated the same shape (oval and cerebriform, respectively) as was previously recorded for vegetable and grape8 while other study results on strawberry isolates had showed sclerotia with a flat shape8. Molecular characterisation showed different frequencies of the four TE genotypes with transposa having been represented by 50%, boty by 29.7%, flipper by 10.9% and vacuma by 9.4% in accordance with those reported in previous studies6,8,22-25 but contradictory to others26,27. Interestingly, this evidence also showed the predominance of transposa in the Botrytis population investigated here and this was consistent with data reported elsewhere6,8,20,22,23,26,28,29-32, followed by boty, flipper and vacuma type. These findings are also compatible with those reported in previous studies24,25. Furthermore, the existence of the four TEs genotypes in the isolates investigated here was demonstrated20-22,33-37 in group 2 of Botrytis population. No correlation was found between isolate phenotype and TE type as the conidial dimensions of all genotypes did not significantly vary between transposa and vacuma isolates and this was consistent with those obtained in other studies conducted on grape and strawberry isolates8. While, a significant variation in virulence was observed on detached lettuce leaves as a lower virulence level of some Botrytis isolates seemed to be correlated with vacuma type as previously reported by many other studies6,8,26-28,30,36,38. Moreover, the results determined a low to moderate resistance towards the hydroxyanilide fungicide, fenhexamid in the three transposa isolates, BCT6, BCGL2 and BCGL5 with average EC50 values of 0.2, 0.3 and 0.5 μg mL–1, respectively. No correlation was found between sensitivity to fenhexamid and the TE type and this was consistent with that reported by other studies28,39 as we did not find any resistant vacuma isolate. In addition, the high resistance to the fungicide fenhexamid was not detected in our current nor previous studies8 indicating that group 1 isolates may not be widely existing at least in Egypt. In fact, Botrytis population of group 1 may exist at low frequency level of 0.7-15%30,31,35,40 or may not exist at all21,32,34. Similar results derived from 33 isolates from Germany41, 55 isolates from Chile, South Asia and Australia32,34 along with 99.3% of many isolates in other studies40 demonstrated that they all belonged to group 2. These findings may confirm that group 1 isolates do exist but in a small number42,43. Although a number of studies reported that Botrytis spp. belonging to group 1 differed from B. pseudocinerea in morphology and phylogeny40,44, BLAST analysis, in the present study, showed a high sequence similarity (98-100%) with that of the GenBank for Botrytis spp. The low frequency of vacuma genotype did not permit to conclude whether the absence of group 1 (B. pseudocinerea) was due to the strategy of sampling used or to the intrinsic absence of vacuma isolates in Egypt. Further study on more vacuma isolates remains to be needed to elucidate this observation. Moreover, phylogenetic analysis revealed that BCLi1 isolate was distantly related to B. cinerea and other Botrytis species. Further molecular analysis of the current isolates is still needed to investigate their phylogenetic relationship and the effect of transposa/vacuma types on their infection severity, through the host-pathogen interaction and fenhexamid resistance of Botrytis isolates, both in vitro and in vivo.
CONCLUSION
In order to investigate whether resistant isolates of Botrytis against fenhexamid started to emerge in Egypt, different symptomatic and symptomless plant samples were collected from various ornamental plants and locations. Based on morphological and molecular characteristics, the pathogen isolates were identified as Botrytis spp. Significant variation in aggressiveness among Botrytis isolates was observed and correlated to vacuma type suggesting a possible effect of TE genotype on Botrytis virulence. The results also revealed a low to moderate resistance of some Botrytis isolates towards fenhexamid using the mycelial growth assay. Such resistance was not found in any vacuma population under study suggesting that the current Botrytis population does not belong to group 1.
SIGNIFICANCE STATEMENT
The current study showed a new emergence of resistant isolates of Botrytis spp. against fenhexamid in Egypt. This is a serious alarm for gray mold control. The results will help the researchers to make attention with fenhexamid application using alternative measures in order to decrease resistance probability to such fungicide.
ACKNOWLEDGMENT
The authors would like to thank Dr. Hala B. Khalil, Department of Genetics, Faculty of Agriculture, Ain Shams University, for the sequence analysis and also Mohamed Gamal, Neveen Sameh, Noran Ashraf and Sarah Osama, the undergraduate students, Department of Plant Pathology, Faculty of Agriculture, Ain Shams University, for their assistance in carrying out some of the study. This research was partially supported by the Science and Technology Development Fund (STDF), Egypt, Grant No. 2131. The developed modified selective medium had been registered as a patent (No. 2012091520) from Academy of Scientific Research and Technology (ASRT), Ministry of Scientific Research, Cairo, Egypt.
REFERENCES
- Elad, Y., B. Williamson, P. Tudzynski and N. Delen, 2007. Botrytis spp. and Diseases They Cause in Agricultural Systems-An Introduction. In: Botrytis: Biology, Pathology and Control, Elad, Y., B. Williamson, P. Tudzynski and N. Delen (Eds.)., Chapter 1. Springer, Netherlands, pp: 1-8.
Direct Link - Dufresne, M., A. Hua-Van, H. Abdel Wahab, S.B. M'Barek and C. Vasnier et al., 2007. Transposition of a fungal miniature inverted-repeat transposable element through the action of a Tc1-Like transposase. Genetics, 175: 441-452.
CrossRefPubMedDirect Link - Lopez-Berges, M.S., A. Di Pietro, M.J. Daboussi, H. Abdel Wahab and C. Vasnier et al., 2009. Identification of virulence genes in Fusarium oxysporum f. sp. lycopersici by large-scale transposon tagging. Mol. Plant Pathol., 10: 95-107.
CrossRefPubMedDirect Link - Abdel Wahab, H., 2015. Characterization of Egyptian Botrytis cinerea isolates from different host plants. Adv. Microbiol., 5: 177-189.
CrossRefDirect Link - Aboelghar, M., M.S. Moustafa, A.M. Ali and H. Abdel Wahab, 2019. Hyperspectral analysis of Botrytis cinerea infected lettuce. Int. J. Agric. Environ. Res., 5: 26-42.
Direct Link - Wagih, E.E., H. Abdel Wahab, M.R.A. Shehata, M.M. Fahmy and M.A. Gaber, 2019. Molecular and pathological variability associated with transposable elements of Botrytis cinerea isolates infecting grape and strawberry in Egypt. Int. J. Phytopathol., 8: 37-51.
CrossRefDirect Link - Fan, F., M. Hahn, G.Q. Li, Y. Lin and C.X. Luo, 2019. Rapid detection of benzimidazole resistance in Botrytis cinerea by loop-mediated isothermal amplification. Phytopathol. Res., Vol. 1, No. 1.
CrossRefDirect Link - Abdel Wahab, H. and N.A.S. Helal, 2013. Evaluation of preharvest bioagent applications for both production and biological control of onion and strawberry under natural Botrytis infections. Afr. J. Plant Sci. Biotechnol., 7: 64-69.
Direct Link - Moller, E.M., G. Bahnweg, H. Sandermann and H.H. Geiger, 1992. A simple and efficient protocol for isolation of high molecular weight DNA from filamentous fungi, fruit bodies and infected plant tissues. Nucleic Acids Res., 20: 6115-6116.
CrossRefDirect Link - Rigotti, S., K. Gindro, H. Richter and O. Viret, 2002. Characterization of molecular markers for specific and sensitive detection of Botrytis cinerea Pers.: Fr. in strawberry (Fragaria × ananassa Duch.) using PCR. FEMS Microbiol. Lett., 209: 169-174.
CrossRefDirect Link - White, T.J., T.D. Bruns, S.B. Lee and J.W. Taylor, 1990. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In: PCR Protocols: A Guide to Methods and Applications, Innis, M.A., D.H. Gelfand, J.J. Sninsky and T.J. White (Eds.), Academic Press, San Diego, CA, USA, ISBN-13: 9780123721808, pp: 315-322.
CrossRefDirect Link - Tamura, K. and M. Nei, 1993. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol., 10: 512-526.
CrossRefPubMedDirect Link - Kumar, S., G. Stecher, M. Li, C. Knyaz and K. Tamura, 2018. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol., 35: 1547-1549.
CrossRefDirect Link - Felsenstein, J., 1985. Confidence limits on phylogenies: An approach using the bootstrap. Evolution, 39: 783-791.
CrossRefDirect Link - Diolez, A., F. Marches, D. Fortini and Y. Brygoo, 1995. Boty, a long-terminal-repeat retroelement in the phytopathogenic fungus Botrytis cinerea. Applied Environ. Microbiol., 61: 103-108.
PubMedDirect Link - Levis, C., D. Fortini and Y. Brygoo, 1997. Flipper, a mobile fot1-like transposable element in Botrytis cinerea. Mol. Gen. Genet., 254: 674-680.
CrossRefPubMedDirect Link - Leyronas, C., F. Bryone, M. Duffaud, C. Troulet and P.C. Nicot, 2015. Assessing host specialization of Botrytis cinerea on lettuce and tomato by genotypic and phenotypic characterization. Plant Pathol., 64: 119-127.
CrossRefDirect Link - Ma, Z. and T.J. Michailides, 2005. Genetic population structure of Botrytis cinerea from different hosts in California. Plant Dis., 89: 1083-1089.
CrossRefDirect Link - Karchani-Balma, S., A. Gautier, A. Raies and E. Fournier, 2008. Geography, plants, and growing systems shape the genetic structure of Tunisian Botrytis cinerea Populations. Phytopathology, 98: 1271-1279.
CrossRefDirect Link - Vaczy, K.Z., E. Sandor, L. Karaffa, E. Fekete and E. Fekete et al., 2008. Sexual recombination in the Botrytis cinerea populations in Hungarian vineyards. Phytopathology, 98: 1312-1319.
CrossRefDirect Link - Johnston, P.R., K. Hoksbergen, D. Park and R.E. Beever, 2014. Genetic diversity of Botrytis in New Zealand vineyards and the significance of its seasonal and regional variation. Plant Pathol., 63: 888-898.
CrossRefDirect Link - Amiri, A., A.I. Zuniga and N.A. Peres, 2018. Prevalence of Botrytis cryptic species in strawberry nursery transplants and strawberry and blueberry commercial fields in the Eastern United States. Plant Dis., 102: 398-404.
CrossRefDirect Link - Zhang, Y., X. Li, F. Shen, H. Xu, Y. Li and D. Liu, 2018. Characterization of Botrytis cinerea isolates from grape vineyards in China. Plant Dis., 102: 40-48.
CrossRefDirect Link - Samuel, S., T. Veloukas, A. Papavasileiou and G.S. Karaoglanidis, 2012. Differences in frequency of transposable elements presence in Botrytis cinerea populations from several hosts in Greece. Plant Dis., 96: 1286-1290.
CrossRefDirect Link - Kumari, S., P. Tayal, E. Sharma and R. Kapoor, 2014. Analyses of genetic and pathogenic variability among Botrytis cinerea isolates. Microbiol. Res., 169: 862-872.
CrossRefDirect Link - Giraud, T., D. Fortini, C. Levis, C. Lamarque, P. Leroux, K. Lobuglio and Y. Brygoo, 1999. Two sibling species of the Botrytis cinerea complex, transposa and vacuma, are found in sympatry on numerous host plants. Phytopathology, 89: 967-973.
CrossRefDirect Link - Muñoz, G., P. Hinrichsen, Y. Brygoo and T. Giraud, 2002. Genetic characterisation of Botrytis cinerea populations in Chile. Mycol. Res., 106: 594-601.
CrossRefDirect Link - Martinez, F., B. Dubos and M. Fermaud, 2005. The role of saprotrophy and virulence in the population dynamics of Botrytis cinerea in Vineyards. Phytopathology, 95: 692-700.
CrossRefDirect Link - Kretchmer, M. and M. Hahn, 2008. Fungicide resistance and genetic diversity of Botrytis cinerea isolates from a vineyard in Germany. J. Plant Dis. Prot., 115: 214-219.
CrossRefDirect Link - Esterio, M., G. Muñoz, C. Ramos, G. Cofré, R. Estévez, A. Salinas and J. Auger, 2011. Characterization of Botrytis cinerea isolates present in thompson seedless table grapes in the central valley of Chile. Plant Dis., 95: 683-690.
CrossRefDirect Link - Fournier, E., T. Giraud and Y. Brygoo, 2005. Partition of the Botrytis cinerea complex in France using multiple gene genealogies. Mycologia, 97: 1251-1267.
CrossRefDirect Link - Isenegger, D.A., P.K. Ades, R. Ford and P.W.J. Taylor, 2008. Status of the Botrytis cinerea species complex and microsatellite analysis of transposon types in South Asia and Australia. Fungal Divers, 29: 17-27.
Direct Link - Martinez, F., M.F. Corio-Costet, C. Levis, M. Coarer and M. Fermaud, 2008. New PCR primers applied to characterize distribution of Botrytis cinerea populations in French Vineyards. Vitis, 47: 217-226.
Direct Link - Muñoz, C., S.G. Talquenca, E. Oriolani and M. Combina, 2010. Genetic characterization of grapevine-infecting Botrytis cinerea isolates from Argentina. Rev. Iberoam. Micol., 27: 66-70.
CrossRefDirect Link - Rajaguru, B.A.P. and M.W. Shaw, 2010. Genetic differentiation between hosts and locations in populations of latent Botrytis cinerea in Southern England. Plant Pathol., 59: 1081-1090.
CrossRefDirect Link - Pande, S., M. Sharma, G.K. Kishore, L. Shivram and U.N. Mangala, 2010. Characterization of Botrytis cinerea isolates from chickpea: DNA polymorphisms, cultural, morphological and virulence characteristics. Afr. J. Biotechnol., 9: 7961-7967.
CrossRefDirect Link - Albertini, C., G. Thebaud, E. Fournier and P. Leroux, 2002. Eburicol 14α-Demethylase gene (CYP51) polymorphism and speciation in Botrytis cinerea. Mycol. Res., 106: 1171-1178.
Direct Link - Saito, S., D. Margosan, T.J. Michailides and C.L. Xiao, 2016. Botrytis californica, a new cryptic species in the B. cinerea species complex causing gray mold in blueberries and table grapes. Mycologia, 108: 330-343.
CrossRefDirect Link - Kecskeméti, E., A. Brathuhn, K.H. Kogel, B. Berkelmann-Löhnertz and A. Reineke, 2014. Presence of transposons and mycoviruses in Botrytis cinerea isolates collected from a German grapevine growing region. J. Phytopathol., 162: 582-595.
CrossRefDirect Link - Saito, S., T.J. Michailides and C.L. Xiao, 2014. First report of Botrytis pseudocinerea causing gray mold on blueberry in North America. Plant Dis., 98: 1743-1743.
CrossRefDirect Link - Zhang, W., J.A. Corwin, D. Copeland, J. Feusier and R. Eshbaugh et al., 2017. Plastic transcriptomes stabilize immunity to pathogen diversity: The jasmonic acid and salicylic acid networks within the Arabidopsis/Botrytis pathosystem. Plant Cell, 29: 2727-2752.
CrossRefDirect Link - Zhou, Y.J., J. Zhang, X.D. Wang, L. Yang and D.H. Jiang et al., 2014. Morphological and phylogenetic identification of Botrytis sinoviticola, a novel cryptic species causing gray mold disease of table grapes (Vitis vinifera) in China. Mycologia, 106: 43-56.
CrossRefDirect Link