
Suvigya Sharma
Department of Biological Sciences, CBSH, G.B. Pant University of Agriculture & Technology, 263145 Pantnagar, India
Ram Prasad
Amity Institute of Microbial Technology, Amity University Uttar Pradesh, Sector 125, Noida, 201313 UP, India
Ajit Varma
Amity Institute of Microbial Technology, Amity University Uttar Pradesh, Sector 125, Noida, 201313 UP, India
Anil Kumar Sharma
Department of Biological Sciences, CBSH, G.B. Pant University of Agriculture & Technology, 263145 Pantnagar, India
LiveDNA: 91.5122
Asian Journal of Plant Pathology
Year: 2017 | Volume: 11 | Issue: 4 | Page No.: 199-202







ABSTRACT
Background and Objective: Arbuscular mycorrhizal fungi (AMF) form symbiotic association with most plants and are known to play significant role in higher nutrient uptake and soil aggregation as well as, carbon sequestration through production of specific glycoprotein characterized as glomalin or glomalin related soil protein (GRSP) contained in their hyphae and spores. The objective of this study was to find out the fungi static effects of this glycoprotein in suppression of growth of two soil borne plant pathogens in vitro. Materials and Methods: Sorghum was used as the test plant, Funneliformis coronatum (F. coronatum), Acaulospora scrobiculata (A. scrobiculata) and Gigaspora margarita (G. margarita) were used as AMF inoculums, Rhizoctonia solani (R. solani) and Colletotrichum falcatum (C. falcatum) were used as test pathogens. The GRSP was extracted from the soil of sorghum pots after harvesting, two different concentrations of GRSP was supplied to the agar medium and test pathogens were inoculated, control plates received extractant from non-AMF inoculated soil. The experiment was performed in completely randomized design with 4 replicates each. The data were analyzed by one-way ANOVA. Results: The level of GRSP production varied with different AMF species. In vitro testing of suppression of Rhizoctonia solani and Colletotrichum falcatum by GRSP extracted from Funneliformis coronatum inoculated soil was higher followed by Gigaspora margarita and Acaulospora scrobiculata inoculated soil. Conclusion: Production of GRSP differs with AMF species and was found to suppress the growth of pathogens in vitro.
PDF Abstract XML References Citation
Received: August 05, 2017;
Accepted: September 23, 2017;
Published: December 13, 2017
How to cite this article
Suvigya Sharma, Ram Prasad, Ajit Varma and Anil Kumar Sharma, 2017. Glycoprotein Associated with Funneliformis coronatum, Gigaspora margarita and Acaulospora scrobiculata Suppress the Plant Pathogens In vitro. Asian Journal of Plant Pathology, 11: 199-202.
URL: https://scialert.net/abstract/?doi=ajppaj.2017.199.202
URL: https://scialert.net/abstract/?doi=ajppaj.2017.199.202
INTRODUCTION
Arbuscular mycorrhizal fungi directly participate in the process to accumulate soil aggregate stability by physically forming a network around soil particles and indirectly by the hyphal exudation of an iron-containing, heat stable glycoprotein (extracted at 121°C) named glomalin as an aggregate binding agent1,2. Glomalin has been operationally defined as glomalin-related soil protein (GRSP) by extraction and detection conditions from soil and it is detected in large amounts in different ecosystems1. The sticky GRSP acts as biological glue, helping to bind soil tiny particles into small aggregates of different sizes3. Well-aggregated soil is stable enough to stand against calamities such as heavy winds and water erosion and has better air and water infiltration rates which work in favor of plants and microbial growth4,5. Additionally, GRSP is recalcitrant enough to have a long residence time in soils (7-42 years)6,7 and plays a pivotal role in long-term carbon/nitrogen storage and heavy metal sequestration2,8. Therefore, the release and accumulation of GRSP in soils can be a very important mechanism for ecological restoration of soils.
Improved plant nutrition, competition for colonization sites and activation of plant defense mechanism are some of the mechanisms proposed in controlling diseases9. Though, role of AM fungi in plant tolerance to disease has already been proven10. The role of GRSP in suppression of Rhizoctonia solani and Colletotrichum falcatum has been worked out probably for the first time in this study.
MATERIALS AND METHODS
The study was carried at Rhizosphere Biology Lab, Department of Biological Sciences, G.B Pant University of Agriculture & Technology, Pantnagar (U.K., India) between January-July, 2017.
GRSP production: The seeds of sorghum were sterilized with 0.1% clorox solution (water, sodium hypochlorite, sodium chloride, sodium carbonate, sodium chlorate, sodium hydroxide and sodium polyacrylate) followed by four rinsing with sterilized water. The infective propagules were estimated from separately maintained AM cultures11. Two hundred infective propagules each of Funneliformis coronatum, Acaulospora scrobiculata and Gigaspora margarita were used to inoculate sorghum in 500 mL pots containing steam sterilized soil: sand (1:1) mixture. AMF inoculum was provided in holes and one germinated sorghum seed was kept on to the inoculum and covered with the same pot soil. Plants were watered 3 times a week with autoclaved distilled water and Hoagland’s solution was added weekly having ¼ dose of phosphorus. AMF inoculum was produced separately for two cycles of 60 days each. Plants without inoculation of any AMF served as control, however they did receive microbial wash from 1 g inoculum of AMF which was obtained by filtering 1g inoculum through Whatman filter paper No. 1. The experiment was conducted in greenhouse with 18 h of light (600 μmol photons m–2 sec–1) and 27±2°C temperature. Each treatment (inoculated and uninoculated) was replicated 4 times. After every harvest, the shoot was harvested from both AMF and control treatments. Shoot was dried in oven at 65°C for 48 h and weighed.
Extraction, precipitation and dialysis of protein: After harvesting the plants, soil from each replicate was mixed thoroughly for extraction of GRSP, 0.25 g samples of dry-sieved 1–2 mm aggregates extracted with 2 mL of extractant (20 mM citrate, pH 7.0 at 121°C for 30 min), following the method of Wright and Upadhyaya12. Extraction process was repeated twice from the same sample. After extraction, precipitation was carried out in 1 N HCl and reconstituted in 100 mM sodium borate (pH 8.0). Dialysis was done with 10 mM sodium borate at pH 8.0. GRSP samples were lyophilized and again reconstituted in distilled water. Reconstituted protein was stored at -20°C.
Estimation of GRSP: GRSP was estimated by modified Bradford dye binding assay13 with BSA as the standard and concentration was extrapolated to mg g–1 soil.
In-vitro evaluation and estimation of inhibition of Rhizoctonia solani and Colletotrichum falcatum: Poisoned food technique was used to see the impact of GRSP on two pathogens i.e., Rhizoctonia solani and Colletotrichum falcatum. In control, extractant was taken from control pot grown in absence of AMF. Two concentrations; 0.4 and 0.8 mg/petri plate of GRSP, were used from three species of F. coronatum, A. scrobiculata and G. margarita, separately. Discs of 5 mm diameter was taken from full grown petri plate of either R. solani or C. falcatum and was placed over GRSP containing petri plate as well as control petri plates. Each treatment was replicated 4 times. Growth inhibition was measured as reduction in the radial growth of pathogen with GRSP over control. Percent inhibition was calculated using formula14:
![]()
Where, C and T is radial growth of pathogen in control and treatment, respectively.
Statistical analysis: The data were analyzed by one-way ANOVA. All treatment means were tested for significant differences at p<0.05.
RESULTS
Plant growth and production of GRSP: Significant growth enhancement of sorghum was observed in either of the AMF inoculated plants. Sorghum plants were inoculated with F. coronatum showed significantly (p<0.05) higher shoot dry weight followed by A. scrobiculata and G. margarita.
Color of GRSP from pot soil varied from dark brown to yellow brown with different AMF species, however, no color was detected in control. Significantly higher (p<0.05) amount of GRSP was recorded in case of F. coronatum followed by G. margarita and A. scrobiculata (Table 1).
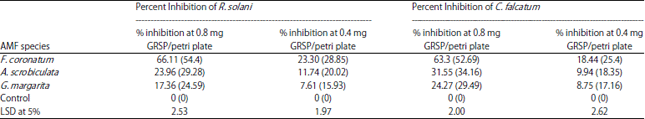
Percent inhibition of R. solani and C. falcatum: Percent inhibition of R. solani and C. falcatum was highest by GRSP extracted from pot inoculated with F. coronatum followed by A. scrobiculata and G. margarita (Table 2). The percent inhibition of pathogen was higher at higher concentration of GRSP in petri plates (Table 2). All the data were significantly different at p<0.05.
DISCUSSION
In the current study, results support the possible role of AM in suppression of soil borne plant pathogens. Research on plant mycorrhizal interactions has mostly focused on the physiology of nutrient/carbon exchange and plant signal-transduction mechanisms controlling the bidirectional interaction. Comparatively less is reported about the mechanisms conferring non-nutritional contributions by mycorrhiza, such as suppression of soil-borne diseases and enhancing plant resistance to pests and diseases15. There are evidences obtained from the decomposition studies that accumulation of GRSP in soil is of AMF origin7. It has been reported that AMF can reduce disease incidence and propagule number of several soil borne pathogens including Aphanomyces, Fusarium, Phytophthora, Pythium and Verticillium species in the plant and mycorrhizosphere10,16,17. Although the mechanisms implicated are still not well characterized, direct and indirect interactions between AMF and pathogens have been put forward as a plausible hypothesis to explain the role of AMF in biological control of root diseases3. Present finding is additional information proving the role of GRSP in suppression of soil borne pathogens.
In this experiment, shoot and root growth of sorghum plants varied with different mycorrhizal species: F. coronatum, A. scrobiculata and G. margarita. It indicates that different species of AMF shows different effects on same variety of plant. GRSP production was found significantly higher in soils used to grow plants with F. coronatum, followed by G. margarita and A. scrobiculata. This could be the result of the performance of an AMF species to produce extra radical hyphae which is ultimately responsible for the release of GRSP in soil. Significant plant growth enhancement by F. coronatum reflects more functionality of extra radical hyphae and therefore, there is a possibility that the amount of extra radical hyphae is more in this case and so, the amount of GRSP. Absence of GRSP in the control pots assures the production of this glycoprotein from AMF only. This describes implode of GRSP from different species of AMF is variable. In vitro studies showed that at different concentrations of GRSP percent, inhibition of R. solani and C. falcatum was significantly higher at higher concentration of GRSP showing antifungal activity.
| Table 1: | Concentration of GRSP from different AMF pure culture |
 | |
| Angularly transformed values are given in parenthesis | |
| Table 2: | Percent inhibition of Rhizoctonia solani and Colletotrichum falcatum at different concentration of GRSP |
 | |
| Angularly transformed values are given in parenthesis | |
The result indicated a direct antibiosis of GRSP in the suppression of pathogen under in vitro condition.
A new putative focusing result was found, which provide a new possible and promising explanation to the involvement of AM fungi in plant protection against soil borne pathogens and point to their use as biological control agents.
CONCLUSION
The current study suggests that the amount of GRSP production varies greatly with different AM species. F. coronatum showed the highest level of GRSP production in the soil. The study reveals alternative roles of GRSP other than soil particle aggregation.
SIGNIFICANCE STATEMENTS
This study discovers the plausible explanation of involvement of AMF in contributing towards plant protection against soil borne plant pathogens that can be beneficial for understanding its mechanism of mycorrhizae induced resistance. This study will help the researchers to uncover the critical areas of mycorrhizae induced resistance related transcriptomics. Thus, a new theory on glomalin related soil protein (GRSP), acting as ‘shield glycoprotein’ against soil borne plant pathogen is being presented.
ACKNOWLEDGMENT
The authors acknowledged the funds received by ICAR, AMAAS to carry out this study. (Grant No: NBAIM/AMAAS/2014-15/BG (1)/144).
REFERENCES
- Rillig, M.C., 2004. Arbuscular mycorrhizae, glomalin and soil aggregation. Can. J. Soil Sci., 84: 355-363.
CrossRefDirect Link - Gonzalez-Chavez, M.C., R. Carillo-Gonzelez, S.F. Wright and K.A. Nichols, 2004. The role of glomalin, a protein produced by arbuscular mycorrhizal fungi, in sequestering potentially toxic elements. Environ. Pollut., 130: 317-323.
CrossRefDirect Link - Rillig, M.C. and D.L. Mummey, 2006. Mycorrhizas and soil structure. New Phytol., 170: 41-53.
CrossRefPubMedDirect Link - Bronick, C.J. and R. Lal, 2005. Soil structure and management: A review. Geoderma, 124: 3-22.
CrossRefDirect Link - Wu, Q.S., R.X. Xia and Y.N. Zou, 2008. Improved soil structure and citrus growth after inoculation with three arbuscular mycorrhizal fungi under drought stress. Eur. J. Soil Biol., 44: 122-128.
CrossRefDirect Link - Rillig, M.C., S.F. Wright, B.A. Kimball, P.J. Pinter, G.W. Wall, M.J. Ottman and S.W. Leavitt, 2001. Elevated carbon dioxide and irrigation effects on water stable aggregates in a Sorghum field: A possible role for arbuscular mycorrhizal fungi. Global Change Biol., 7: 333-337.
CrossRefDirect Link - Steinberg, P.D. and M.C. Rillig, 2003. Differential decomposition of arbuscular mycorrhizal fungal hyphae and glomalin. Soil Biol. Biochem., 35: 191-194.
CrossRefDirect Link - Malekzadeh, E., N. Aliasgharzad, J. Majidi, J. Abdolalizadeh and L. Aghebati-Maleki, 2016. Contribution of glomalin to Pb sequestration by arbuscular mycorrhizal fungus in a sand culture system with clover plant. Eur. J. Soil Biol., 74: 45-51.
CrossRefDirect Link - Pozo, M.J. and C. Azcon-Aguilar, 2007. Unraveling mycorrhiza-induced resistance. Curr. Opin. Plant Biol., 10: 393-398.
CrossRefDirect Link - Veresoglou, S.D. and M.C. Rillig, 2012. Suppression of fungal and nematode plant pathogens through arbuscular mycorrhizal fungi. Biol. Lett., 8: 214-217.
CrossRefDirect Link - Sharma, M.P., A. Gaur, N.P. Bhatia and A. Adholeya, 1996. Growth responses and dependence of Acacia nilotica var. cupriciformis on the indigenous arbuscular mycorrhizal consortium of a marginal wasteland soil. Mycorrhiza, 6: 441-446.
CrossRefDirect Link - Wright, S.F. and A. Upadhyaya, 1996. Extraction of an abundant and unusual protein from soil and comparison with hyphal protein of arbuscular mycorrhizal fungi. Soil Sci., 161: 575-586.
CrossRefDirect Link - Wright, S.F. and A. Upadhyaya, 1998. A survey of soils for aggregate stability and glomalin, a glycoprotein produced by hyphae of arbuscular mycorrhizal fungi. Plant Soil, 198: 97-107.
CrossRefDirect Link - Al-Reza, S.M., A. Rahman, Y. Ahmed and S.C. Kang, 2010. Inhibition of plant pathogens in vitro and in vivo with essential oil and organic extracts of Cestrum nocturnum L. Pestic. Biochem. Physiol., 96: 86-92.
CrossRefDirect Link - Cameron, D.D., 2010. Arbuscular mycorrhizal fungi as (agro)ecosystem engineers. Plant Soil, 333: 1-5.
CrossRefDirect Link - Campos-Soriano, L., J. Garcia-Martinez and B.S. Segundo, 2012. The arbuscular mycorrhizal symbiosis promotes the systemic induction of regulatory defence‐related genes in rice leaves and confers resistance to pathogen infection. Mol. Plant Pathol., 13: 579-592.
CrossRefDirect Link - Azcon-Aguilar, C. and J.M. Barea, 1997. Arbuscular mycorrhizas and biological control of soil-borne plant pathogens-an overview of the mechanisms involved. Mycorrhiza, 6: 457-464.
CrossRefDirect Link