
Ibrahim A.A. Adss
Division of Genetics, Faculty of Agriculture, Damanhur University, Al-Beheira, Egypt
Muhammad A. Abdel-Gayed
Department of Onion and Garlic Diseases Research, Plant Pathology Research Institute, Agriculture Research Center, Giza, Egypt
William Botros
Department of Nucleic Acids, Genetic Engineering and Biotechnology Research Institute, City for Scientific Research and Technology Applications, Alexandria, Egypt
Elsayed E. Hafez
Department of Plant Protection and Biomolecular Diagnosis, Arid Lands Cultivation Research Institute (ALCRI), City for Scientific Research and Technology Applications, Alexandria, Egypt
LiveDNA: 20.3216
Asian Journal of Plant Pathology
Year: 2017 | Volume: 11 | Issue: 1 | Page No.: 18-27







ABSTRACT
Background: Tomato belongs to the Solanaceae family alongside other economically important crops such as pepper, eggplant and potato. Fruit rot disease caused by Alternaria solani is the most severe disease of tomato. Materials and Methods: Seven isolates of Alternaria solani were collected from different tomato fields in diverse localities of Al-Behiera Governorate. Pathogenicity tests of the seven isolates of A. solani were performed on fresh tomato fruits of the 1077 cv. The isolates differed in their virulence as evidenced by the diameters of rotted areas in the inoculated tomato fruits. Results: The isolates 7, 5 and 6 were highly virulent, while the isolates 2 were moderately virulent. The polygalacturonase enzyme activity was examined for all isolates and isolate 7 revealed the maximum activity (1.13 U mL–1 min–1), followed by isolates 5 (1.1 U mL–1 min–1), while the activity was low with isolate 1 (0.42 U mL–1 min–1), isolates 2 and 4 (0.6 and 0.7 U mL–1 min–1), respectively. On the other hand, both of isolates 3 and 6 gave a moderate activity (0.89 and 0.95 U mL–1 min–1), respectively. Genetic diversity was studied using RAPD-PCR and ISSR markers. Results exposed that inclusive genetic variations between the isolates were observed and the polymorphic percentage was ranged from 5.8-80%. While, the ISSR analysis grouped the 7 fungal isolates into two main clusters; the first cluster include isolates 1 and 2 but cluster two was divided into two groups; group one contains isolates 4-6 while group two include isolate 7. Conclusion: The polygalacturonase activity and ISSR gave the same results but the RAPD gave completely different results. That means, using functional gene for differentiation between plentiful closed isolates is more preferable than RAPD. Moreover, the ISSR-PCR is more confident than the RAPD-PCR.
PDF Abstract XML References Citation
Received: November 01, 2016;
Accepted: November 25, 2016;
Published: December 15, 2016
How to cite this article
Ibrahim A.A. Adss, Muhammad A. Abdel-Gayed, William Botros and Elsayed E. Hafez, 2017. Multilocus Genetic Techniques, RAPD-PCR and ISSR-PCR Markers and Polygalacturonase Activity as Tools for Differentiation Among Alternaria solani Isolates on Tomato Fruits and Relation to their Pathogenicity in Egypt. Asian Journal of Plant Pathology, 11: 18-27.
URL: https://scialert.net/abstract/?doi=ajppaj.2017.18.27
URL: https://scialert.net/abstract/?doi=ajppaj.2017.18.27
INTRODUCTION
The tomato belongs to the Solanaceae family alongside other economically important crops such as pepper, eggplant and potato. The tomato was classified by Miller1 as Lycopersicon esculentum and renamed by Child2 and Peralta et al.3 as Solanum lycopersicum. Tomato crop is regarded as one of the most important economic vegetables in Egypt as well as in many countries of the whole world. Actuality, it is a widely cultivated crop and the top one in value among the vegetables4. Tomatoes are attacking with several fungal pathogens in the field and marketing, which cause serious diseases affecting the quality and quantity of fruit yield5.
Fruit rot disease caused by Alternaria solani is the most severe disease of tomato6. Considerable effort is invested in protecting the agricultural production against Alternaria before and after harvest. Under Egyptian conditions tomato plants are susceptible to infection by Alternaria solani, which causes a great reduction in the quantity and quality of fruit yield. Alternaria fungus can cause numerous diseases on all parts of tomato plant (leaf blight, stem collar rot and fruit lesions) and results in severe damage during all stages of plant growth and development7.
The cell wall degradation enzymes can be secreted by phytopathogens to facilitate their penetration and entrance into the plant cell and use the components of the host cell walls as nutrient source8. Most of phytopathogens are secreted different pectolytic enzymes to degrade the middle lamella of the plant cell wall, these pectolytic enzymes are an important tools of the virulence of invaded pathogens9. Alternaria solani causes tomato fruit rot, this pathogen has the ability to produce polygalacturonase enzyme and other cell wall degrading enzymes9,10.
For most genetic variation studies, a good genetic marker is defined by high genetic variability and the ability to generate multi locus data from the genome under study11. The generation of ISSR markers makes use of micro satellite sequences that are highly variable and ubiquitously distributed across the genome, at the same time achieving higher reproducibility compared to using RAPDs and costs less in terms of time and money compared to using AFLPs. All these make ISSR an ideal genetic marker for various studies, most notably on genetic variation/diversity11-13 DNA fingerprinting and phylogenetics6.
A great number of molecular markers (RFLP, AFLP, RAPD, CAPs and ISSR) are mapped onto the genetic map of fungi. Multi locus techniques such as RAPD Random Amplified Polymorphic DNA (RAPD) and Inter Simple Sequence Repeats (ISSR) are convenient and allow ready scoring of numerous polymorphic loci but are limited by features including difficulties of reproducibility and the assumption that co-migrating bands are identical. Random amplified polymorphic DNA (RAPD) and Inter Simple Sequence Repeats (ISSR) have been applied to evaluate the genetic diversity and typing of Alternaria solani strains14. However, each method has advantages and disadvantages.
The RAPD-PCR and ISSR-PCR have been developed since more than 30 years ago. The two techniques have wide range of applications. Studying the diversity of various genetic resources with RAPD and ISSR is important due to their greater stability, efficiency and accuracy in comparison to morphology15,16.
The Random Amplified Polymorphic DNA (RAPD) which is a Polymerase Chain Reaction (PCR)-based technique in which amplification of random DNA fragments with single primers of arbitrary nucleotide sequence16,17, RAPD Polymerase Chain Reaction (PCR) is considered to be a useful approach to obtain genetic data18,19. The RAPD-PCR is conducted and used as powerful tool for genetic studies involving molecular genetic diversity of microorganisms. It can be applied to differentiate races, strains and pathogenic or nonpathogenic isolates of fungi. Generally very short (10 or fewer bases) pieces of DNA from the selected source are used as primers. The RAPD markers are robust genetically and have been used widely for genetic mapping in plants20 and other organisms21. The RAPD method is simple, sensitive and rapid for the detection, differentiation and determination of phylogenetic relationship between isolates of a morphological species of several pathogens22. However, RAPD-PCR has a tendency for a reproducibility problem in the band patterns23. For this reason, the RAPD reaction conditions should be highly standardized18,23.
We can use the RAPD-PCR to study the fungal taxonomy24. Many of studies used RAPD marker to assess the genetic diversity of Alternaria isolates such as15, assessment of genetic diversity between Alternaria alternate isolates were collected in Rajasthan.
The ISSR is being able to create polymorphism patterns among near-organisms and their reproducibility25. Guo et al.26 used ISSR primers to study the genetic diversity of 112 isolates of Alternaria alternata isolated from pine in China.
De la Torre et al.27 reported that 13 ISSR primers yielded a reproducible banding pattern, with 701 amplified loci and 98% of polymorphism among 35 genotypes of Calibrachoa caesia. The ISSR primers 5’CT, 5’CA, 5’GA, 5’GACA, 3’CAC, 3’TG and 3’TC generated 100% polymorphic patterns. The Rp values ranged from 23.20-10.29 for 5’GACA and 3’AG primers, respectively, while the average values for MI and PIC were 0.367 and 0.231, respectively. The more informative primers were 5’GACA and 5’GA and the less informative was 3’AC. Simple matching coefficient of similarity varied from 0.8875-0.6659, indicating high levels of genetic similarity among the genotypes examined.
The main objective of the present study is using polygalacturonase activity and genetic diversity (RAPD and ISSR marker) as tools for differentiation among 7 isolates of A. solani and their pathogenic capability.
MATERIALS AND METHODS
Collection of diseased tomato fruits samples: Sampling of blighted tomato fruits was carried out from different tomato fields at diverse localities in Al-Behiera Governorate of Egypt. The samples were examined carefully in the field, kept in sterilized polyethylene bags and brought to the laboratory for isolation and purification of the causal pathogen and complete the study.
Isolation and purification of causal pathogen: For isolation and purification of the causal pathogen, the diseased portions from tomato fruits were cut into small pieces (5 mm2). The pieces were washed thoroughly with tap water, then air dried and surface sterilized by immersing in 2% sodium hypochlorite (NaOCl) solution for 2 min, rinsed in sterilized distilled water several times and gently blotted dry between two sterilized filter papers then plated individually, on Potato Dextrose Agar (PDA) medium supplemented with streptomycin (50 μg cm–3). The plates were incubated at 28°C and examined for the pathogen growth. After 4 days, the young growing hyphal tips were transferred to freshly prepared PDA plates by hypha tip technique according to Karr and Albersheim28.
Identification of the isolated pathogen: To identify the pathogen isolates; the cultures were tentatively identified as Alternaria solani based on standard mycological parameters according to Subramanian29 and maintained on PDA slants at 4°C for further study.
Pathogenicity tests: To confirm pathogenicity, healthy tomato fruits semi of the 1077 cv were surface sterilized by immersing in 3% sodium hypochlorite (NaOCl) solution for 3 min, rinsed in sterilized distilled water several times. After surface sterilization, the fruits were inoculated with 8.0 mm diameter inoculum discs by cavity method30. All the inoculated fruits were kept in moist chamber (80-100% relative humidity) and then incubated at 28°C for 10 days. This experiment was performed in three replicates for each isolate. At the end of incubation period, the diameter of rotted area was measured in millimeter.
Production of polygalacturonase enzyme: To estimate the difference among 7 isolates of A. solani using the polygalacturonase (PG) enzyme activity, we used semi ripe tomato fruit medium6. To study polygalacturonase (PG) enzyme production in vitro, A. solani isolates were grown in 150 mL Erlenmeyer flasks containing 25 mL of sterilized semi ripe tomato fruit medium. The flasks were inoculated by a disc (0.5 mm) taken from the edge of 4 days old culture for each isolates. The inoculated flasks were incubated for 10 days at 28°C. Three replicates were done for each isolate. At the end of incubation period, fungal mats were removed and the culture fluids were diluted with 35 mL of distilled sterilized water. The obtained filtrates were centrifuged for 15 min at 8000×g and the supernatant was used as crude enzyme extract.
Assay and determination of polygalacturonase enzyme activity: Polygalacturonase (PG) activity was determined using the thiobarbituric acid (TBA) method31,32 with some modifications. The PG activity was assayed by adding the crude enzyme to tubes containing 2.5 mL of 0.5% polygalacturonic acid in 0.05 M sodium acetate buffer, pH 5.5 and 0.1 M NaCl33. The reaction mixture was incubated for 2 h at 30°C and the reaction was stopped by heating the tubes at 100°C for 10 min. After cooling at room temperature the reaction mixture was centrifuged at 10,000×g for 10 min and 2 mL of the supernatant were mixed with an equal volume of TBA reagent34 and heated at 100°C for 30 min. The activity was determined by the increase in the absorbance (optical density) at 515 nm (OD: 515). Zero time reaction mixture containing active enzyme was used as control.
The amount of galacturonic acid released per milliliters per minute was calculated from standard curve of galacturonic acid. About 1 U of PGase activity was defined as the amount of enzyme required to release 1 μmol mm–1 min–1 galacturonic acid under standard assay conditions.
DNA extraction and RAPD-PCR procedure: Mycelia of 7 A. solani isolates grown in Potato Dextrose Broth (PDB) medium were harvested washed and were kept at -20°C. The DNA was extracted and performed using DNA extraction kit (QIAgene, Germany) according to the manufacture procedures.
| Table 1: | Primers used in this study |
 | |
The DNA quantity and quality of extracted DNA were determined by spectrophotometer and agarose gel electrophoresis.
Four random primers each consists of 10 mers were used to differentiate between the obtained A. solani isolates (Table 1). The RAPD-PCR reaction was performed according to Abd El-Gayed35.
ISSR-PCR: Four ISSR primers were used to fingerprint, the 7 isolates of A. solani and the primers are illustrated in Table 1. The PCR amplification was carried out according to Bagherabadi et al.36 in total volume of 25 μL containing; 2 μL DNA, 2 μL of primer, 2.5 μL 1x buffer, 2.5 μL 3.2 mM MgCl2, 2.5 μL 0.6 mM dNTPs and 0.2 μL (5 U μL–1) Taq DNA polymerase (Promega Germany). The PCR conditions were: 94°C for 5 min, followed b y 40 cycles of 1 min at 94°C for 75 sec (Depending on the type of primer used), 2 min at 72°C and 10 min of final extension at 72°C. The PCR products were separated by gel electrophoresis on 1% (w/v) agarose in TBE buffer and photographed after staining with ethidium bromide.
RAPD and ISSR analysis: Pair wise genetic comparisons based on RAPD and ISSR fingerprints among isolates were calculated using Jaccard’s similarity coefficient embedded in NTSYS-pc software37. The polymorphic bands were scored as; 1 for the presence and 0 for absence of the band. It was assumed that the bands with the same size were identical (monomorphic). Cluster analysis of the data matrix was performed by the unweighted pair group method with arithmetic means (UPGMA) with Jaccard’s similarity coefficient38.
RESULTS
Isolation, purification and identification of the causal pathogen: The pathogen was successfully isolated on PDA medium through 6 days, the isolation trials were leaded to 7 fungal isolates. The obtained isolates of the causal pathogen from blighted tomato were identified as Alternaria solani based on morphological, microscopical and cultural characteristics of mycelia, conidiophores according to Subramanian29.
| Table 2: | Pathogenicity tests of 7 isolates of Alternaria solani on fruits of 1077 tomato cultivar |
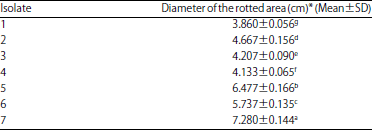 | |
*All data are averages of triplicate measurements±standard deviation, LSD0.05 for A. solani isolates = 0.017, means followed by the different letter(s) are significantly different at p = 0.05 | |
| Table 3: | Enzyme activity of polygalacturonase produced by 7 isolates of A. solani |
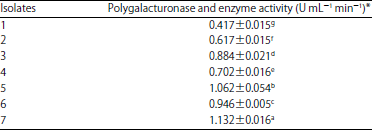 | |
*All data are averages of triplicate measurements±standard deviation, LSD0.05 for fungal isolates = 0.03, means followed by the different letter(s) are significantly different at p = 0.05 | |
Pathogenecity tests: Pathogenicity tests of the 7 isolates of A. solani were performed on fresh tomato fruits of the 1077 cv. Symptoms of the disease on tomato fruits were examined 10 days post inoculation. The results indicated that all tested isolates contrasted in their virulence as evidenced by the diameters of rotted areas in the inoculated tomato fruits. Data in Table 2 and Fig. 1 demonstrated that all isolates of A. solani were virulent. The isolates 7, 5 and 6 were highly virulent on tomato fruits produced the highest rotten area diameter with average 7.280, 6.477 and 5.737 cm, respectively. The isolates 2 was moderately virulent (4.667 cm), however, the isolates 3, 4 and 1 were weakly virulent produced the lowest rotten area diameter with average 4.207, 4.133 and 3.860 cm, respectively.
Production of polygalacturonase and enzyme activity: The results attained in Table 3 showed that the all A. solani isolates were able to produce polygalacturonase enzyme and the levels of polygalacturonase activity were varied among the obtained 7 isolates. It was observed that isolate 7 showed high productivity of polygalacturonase enzyme (1.13 U mL–1 min–1) followed by isolate 5 (1.06 U mL–1 min–1), while low production activity was (0.417, 0.617 and 0.702 U mL–1 min–1) and occasioned by isolates 1, 2 and 4, respectively.
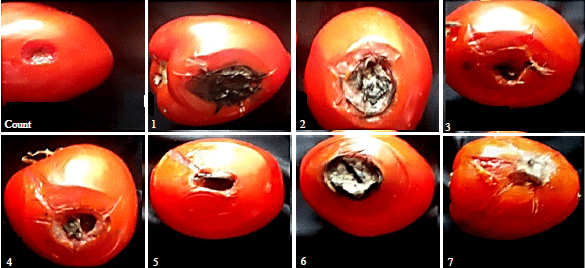 | |
| Fig. 1: | Pathogenicity tests of 7 isolates of Alternaria solani on fruits of 1077 tomato cultivar |
| Table 4: | Numbers of polymorphic and monomorphic bands and percentage of polymorphism detected by each RAPD primers among 7 isolates of A. solani varied in their virulence on tomato fruits |
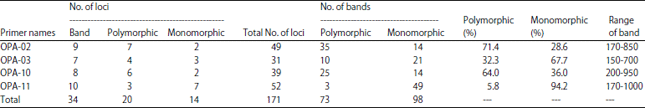 | |
| Polymorphism = No. of polymorphic bands/total No. of bands×100 | |
But the production activity of the two isolates 3 and 6 were moderated 0.884 and 0.946 U mL–1 min–1, respectively.
Random Amplified Polymorphic DNA (RAPD-PCR) of A. solani isolates: Results obtained by RAPD-PCR and presented in Table 4 and Fig. 2 revealed that all the four used primers produced wide range of fragments with different molecular sizes (150-1000 bp). The high number of bands (49 bands) were obtained by primer OPA-02 produced 49 loci most of them are polymorphic, while primer OPA-03 produced a low number of bands 31 loci. The high number of polymorphic bands obtained by OPA-02 was 35 loci (71.4%), while the low number of polymorphic bands was 3 loci by primer OPA-11 (5.8%). Jaccard similarity coefficient was calculated from the RAPD data and it was detected that the similarity between the 7 examined fungal isolates is ranged from 67.2-80.6%. The RAPD-PCR divided the seven A. solani isolates into two clusters (Fig. 3). Isolate one formed the cluster 1 but the other 6 isolates formed the second cluster which grouped into two main groups (A and B). Group A contains isolate 5, while group B were divided into two subgroups; subgroup 1 include isolates 2, 3 and 4 but subgroup 2 contains isolates 6 and 7.
Inter Simple Sequence Repeats (ISSR-PCR) polymorphism of A. solani isolates: The fragment size produced by ISSR primers were ranged from 100-3000 bp (Table 5, Fig. 4). Two hundred bands were obtained by the four used primers (88 polymorphic and 112 monomorphic bands). The polymorphic percentage of the 200 bands ranged from 55.3-25.53%. The number of bands which obtained by each primer was 54 with UBC807 primer, 52 with the UBC35 primer, 47 with both UBC809 and UBC834 primers. According to the Fig. 4 and 5, UBC809 primer had the highest polymorphism (55.3%) but UBC834 primer gave the lowest polymorphism (25.53%). Finally, 38 bands loci were amplified by ISSR primers which 22 loci of them were polymorphic and 16 loci were monomorphic.
The UPGMA cluster analysis based on pairwise genetic similarity coefficients revealed that the similarity between the 7 isolates ranged from 65.7-80.6% (Fig. 5). Two major clusters were observed, the first cluster include isolates 1 and 2 but the rested isolates formed cluster two.
| Table 5: | Number of polymorphic and monomorphic bands and percentage of polymorphism detected by each ISSR primers among seven isolates of A. solani varied in their virulence on tomato fruits |
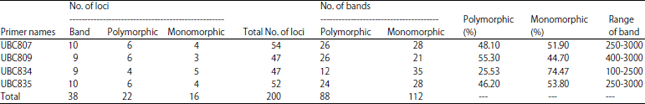 | |
| Polymorphism = No. of polymorphic bands/Total No. of bands ×100 | |
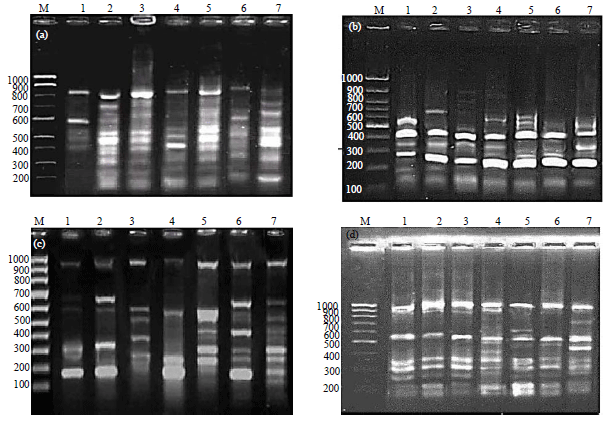 | |
| Fig. 2(a-d): | Electrophoretic patterns of RAPD-PCR products of 7 A. solani isolates with four primers, (a) OPA-2, (b) OPA-3, (c) OPA-10 and (d) OPA-11. Lane on far left side of each gel labeled M is loaded with DNA marker with uppermost band 1000 bp. Samples from 1-7 are the 7 A. solani isolates isolated from different localities and varied in their virulence on tomato fruits |
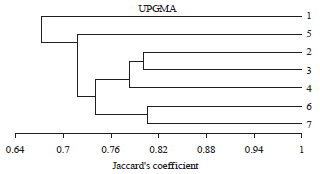 | |
| Fig. 3: | A dendreogram illustrates the similarity and the relationship between the 7 examined isolates of A. solani based on the RAPD analysis of Jaccard’s similarity coefficient and UPGMA cluster analysis |
Cluster two was divided into two groups; group one include isolates 3-6 while group two contains only isolate 7.
DISCUSSION
The main objective of this study was to investigate and demonstrate the variation among 7 isolates of A. solani the causal pathogen of tomato fruit blight collected from different localities in Al-Behiera Governorate of Egypt. The variation was studied based on pathogenicity test, polygalacturonase enzyme activity and multi-locus genetic markers (RAPD-PCR and ISSR-PCR). The isolates of A. solani were isolated from blighted tomato fruit and it was grown and purified on PDA medium.
Pathogenicity tests of the 7 isolates of A. solani were performed on fresh tomato fruits of the 1077 cv. The isolates differed in their virulence as evidenced by the diameters of rotted areas in the inoculated tomato fruits. The isolates 5-7 were highly virulent, while the isolates 2 was moderately virulent. The same results were obtained by Shafiei-Astani et al.13 and Chaurasia and Chaurasia39 while A. solani was isolated as a pathogen from the fruit rot disease of tomato.
Pectic substances play an important role in maintaining the architecture of plant cell walls. Depolymerization of pectin has been shown to be a prerequisite for degradation of other cell wall polymers40. The polygalacturonase is apectic enzyme involved in cell wall-degradation. The importance of poygalacturonase produced by many plant pathogens has been strongly implicated in contributing to pathogenicity and virulence12.
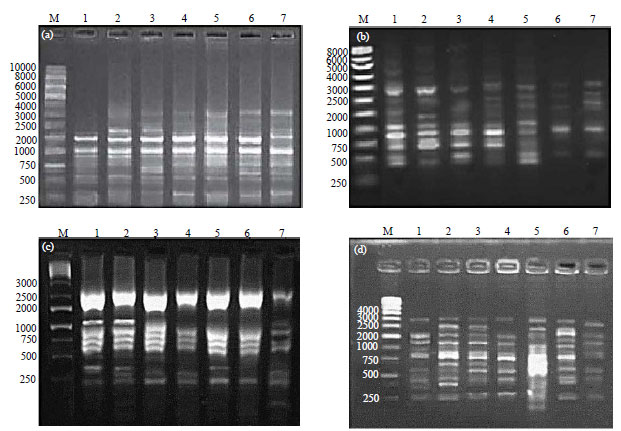 | |
| Fig. 4(a-d): | Electrophoretic patterns of ISSR-PCR products of 7 A. solani isolates with four primers, (a) UBC 807, (b) UBC 809, (c) UBC 834 and (d) UBC 835. Lane on far left side of each gel labeled M is loaded with DNA marker. Samples from 1-7 are the 7 A. solani isolates isolated from different localities and varied in their virulence on tomato fruits |
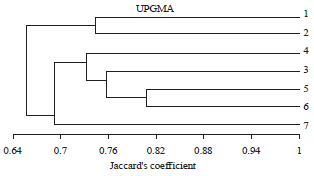 | |
| Fig. 5: | A dendreogram illustrates the similarity and the relationship between the seven examined isolates of A. solani based on the ISSR analysis of Jaccard’s similarity coefficient and UPGMA cluster analysis |
Moreover, the enzyme is supposed to act before other pectic enzymes41 and is the first pectolytic enzyme secreted by pathogenic fungi when cultured on isolated cell walls42. Data showed that the all A. solani isolates were able to produce polygalacturonase enzyme. The levels of polygalacturonase activity were different among the isolates, high activity by isolates 7 and 5, while the isolates 1, 2 and 4 were low activity. The isolates 3 and 6 were moderately of polygalacturonase activity. These results agree with the results obtained by Chaurasia et al.9, who reported that A. solani isolate was able to produce polygalacturonase enzyme with different levels of activity. Clearly, there is a strong relationship between the ability of A. solani isolates to produce polygalacturonase enzyme and their virulence and pathogenicity on tomato fruits. Therefore, the highly pathogenic isolate 7 had the highly capability of polygalacturonase enzyme production and activity.
The development of molecular techniques has revolutionized and energized fungal population genetics by providing numerous readily available genetic markers12. The Random Amplified Polymorphic DNA (RAPD) technique is a PCR procedure involving arbitrary primers. It can be applied to differentiate races, strains and pathogenic or nonpathogenic isolates of the fungi. Generally, very short (10 or fewer bases) pieces of DNA from the selected source are used as primers. Multi locus techniques such as RAPDs and ISSR are convenient and allow ready scoring of numerous polymorphic loci but are limited by features including difficulties of reproducibility and the assumption that co-migrating bands are identical. The RAPD and ISSR markers can demonstrate similarities and dissimilarities between different isolates of same species even when a morphological description is severely limited.
In the present study, RAPD polymerase chain reaction and ISSR-PCR procedures were used to study the genetic diversity among 7 isolates of A. solani varied in their morphology and their virulence, on tomato cv 1077, using genomic DNA of all tested isolates and four arbitrary primers for each technique. It was observed that a wide genetic variation between A. solani isolates were evident due to the high number of polymorphic bands. In case of RAPD-PCR, primer OPA-02 gave polymorphic with 71.4%, while it was decreased into 5.8% with the primer OPA-11. The frequencies of the resulted polymorphic bands varied from primer to primer. Data obtained by RAPD-PCR grouped the A. solani isolates were divided into two large clusters. The RAPD was come up to differentiate between Alternaria alternate isolates26.
In this study the percentage of polymorphic bands obtained by ISSR-PCR were ranged from 55.3-25.53%. Primer UBC809 gave the highest polymorphism (55.3%) but the lowest was obtained by primer UBC834 (25.53%). Genetic similarity coefficients were divided the isolates into two major cluster the first cluster include isolates 1 and 2 but the second cluster contains the rest of the isolates which are differed into two main groups. Analysis of ISSR polymorphism showed a higher variability for A. solani isolates. Differences in ISSR polymorphism were observed in strains isolated from either tomato or cabbage43. It can conclude that, the isolate fingerprinting using polygalacturonase activity and ISSR gave the same results but the differentiation with RAPD was completely different. That means, using functional gene for differentiate between plentiful closed isolates is more preferable than RAPD. Moreover, the ISSR-PCR is more confident than the RAPD-PCR. Boccara et al.44 assessed the genetic diversity among 52 strains of Erwinia chrysanthemi. The similarity coefficients obtained with a probe encoding pathogenicity genes of E. chrysanthemi gave similar results compared to those obtained with randomly selected probes from a cosmid library of E. chrysanthemi 3937.
CONCLUSION
The polygalacturonase activity and ISSR gave the same results but the RAPD gave completely different results. That means, using functional gene for differentiation between plentiful closed isolates is more preferable than RAPD. Moreover, the ISSR-PCR is more confident than the RAPD-PCR.
ACKNOWLEDGMENTS
We do thank all the studies in the Department of Genetics, Faculty of Agriculture, Damanhur University. Also, the engineers are studying in the green house of the Faculty of the Agriculture and the Agricultural Research Center, Giza, Egypt as well.
REFERENCES
- Peralta, I.E., S. Knapp and D.M. Spooner, 2006. The taxonomy of tomatoes. A revision of wild tomatoes (Solanum L. section Lycopersicon (Mill.) Wettst.) and their out group relatives (Solanum sections Juglandis folium (Rydb.) child a Lycopersicoides (Child) Peralta). Monogr. Syst. Bot. Missouri Bot. Garden: Syst. Bot. Monogr., 84: 1-186.
- Iruela, M., J. Rubio, J.I. Cubero, J. Gil and T. Millan, 2002. Phylogenetic analysis in the genus cicer and cultivated chickpea using RAPD and ISSR markers. Theor. Applied Genet., 104: 643-651.
CrossRefPubMedDirect Link - Abada, K.A., S.H. Mostafa and M.R. Hillal, 2008. Effect of some chemical salts on suppressing the infection by early blight disease of tomato. Egypt. J. Applied Sci., 23: 47-58.
Direct Link - Al-Hindi, R.R., A.R. Al-Najada and S.A. Mohamed, 2011. Isolation and identification of some fruit spoilage fungi: Screening of plant cell wall degrading enzymes. Afr. J. Microbiol. Res., 5: 443-448.
Direct Link - Chaurasia, A.K., S. Chaurasia, S. Chaurasia and S. Chaurasia, 2014. In vitro and in vivo production of polygalacturonase, polymethylgalacturonase and cellulase enzymes by Alternaria solani at different incubation periods. Afr. J. Plant Sci., 8: 248-253.
CrossRefDirect Link - Anne, C., 2006. Choosing the right molecular genetic markers for studying biodiversity: From molecular evolution to practical aspects. Genetica, 127: 101-120.
CrossRefPubMedDirect Link - Wang, X., R. Yang, S. Feng, X. Hou, Y. Zhang, Y. Li and Y. Ren, 2012. Genetic variation in Rheum palmatum and Rheum tanguticum (Polygonaceae), two medicinally and endemic species in China using ISSR markers. PLOS One, Vol. 7.
CrossRefDirect Link - Shafiei-Astani, B., A.H.K. Ong, A. Valdiani, S.G. Tan and C.Y.S. Yien et al., 2015. Molecular genetic variation and structure of Southeast Asian crocodile (Tomistoma schlegelii): Comparative potentials of SSRs versus ISSRs. Gene, 571: 107-116.
CrossRefPubMedDirect Link - Troncoso-Rojas, R., M.E. Baez-Flores, B. Pryor, H.S. Garcia. and M. Tiznado-Hernandez, 2013. Inter simple sequence repeat polymorphism in Alternaria genomic DNA exposed to lethal concentrations of isothiocyanates. Afr. J. Microbiol. Res., 7: 838-852.
Direct Link - Guo, L.D., K.D. Hyde and E.C.Y. Liew, 2001. Detection and taxonomic placement of endophytic fungi within frond tissues of Livistona chinensis based on rDNA sequences. Mol. Phylogenet. Evol., 20: 1-13.
CrossRefPubMedDirect Link - Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski and S.V. Tingey, 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res., 18: 6531-6535.
CrossRefPubMedDirect Link - Hilton, A.C., J.G. Banks and C.W. Penn, 1997. Optimization of RAPD for fingerprinting Salmonella. Lett. Applied Microbiol., 24: 243-284.
CrossRefPubMedDirect Link - Ramos, J.R., M.P. Telles, J.A. Diniz-Filho, T.N. Soares, D.B. Melo and G. Oliveira, 2008. Optimizing reproducibility evaluation for random amplified polymorphic DNA markers. Genet. Mol. Res., 7: 1384-1391.
PubMedDirect Link - Singh, S., P. Goswami, R. Singh and K.J. Heller, 2009. Application of molecular identification tools for Lactobacillus, with a focus on discrimination between closely related species: A review. LWT-Food Sci. Technol., 42: 448-457.
CrossRefDirect Link - Grimmer, M.K., S. Trybush, S. Hanley, S.A. Francis, A. Karp and M.J.C. Asher, 2007. An anchored linkage map for sugar beet based on AFLP, SNP and RAPD markers and QTL mapping of a new source of resistance to Beet necrotic yellow vein virus. Plant Breed., 114: 1151-1160.
CrossRefPubMedDirect Link - Abd El-Salam, K.A., J.R. Guo, F. Schneider, A.M. Asran-Amal and J.A. Verreet, 2004. Comparative assessment of genotyping methods for study genetic diversity of Fusarium oxysporum isolates. Polish J. Microbiol., 53: 167-174.
Direct Link - Sharma, R.L., 1994. Prevalence of postharvest diseases of [tomato in] Himachal Pradesh. Plant Dis. Res., 9: 195-197.
Direct Link - Guo, L.D., G.R. Huang, Y. Wang, W.H. He, W.H. Zheng and K.D. Hyde, 2003. Molecular identification of white morphotype strains of endophytic fungi from Pinus tabulaeformis. Mycol. Res., 107: 680-688.
CrossRefPubMedDirect Link - Zietkiewicz, E., A. Rafalski and D. Labuda, 1994. Genome fingerprinting by Simple Sequence Repeat (SSR)-anchored polymerase chain reaction amplification. Genomics, 20: 176-183.
CrossRefPubMedDirect Link - Guo, L.D., L. Xu, W.H. Zheng and K.D. Hyde, 2004. Genetic variation of Alternaria alternate, an endophytic fungus isolated from Pinus tabulaeformis as determined by random amplified microsatellites (RAMS). Fungal Diversity, 16: 53-65.
Direct Link - De la Torre, M.P., M. Garcia, R. Heinz and A. Escandon, 2012. Analysis of genetic variability by ISSR markers in Calibrachoa caesia. Electron. J. Biotechnol. Vol. 15, No. 5.
CrossRefDirect Link - Karr, A.L. and P. Albersheim, 1970. Polysaccharide-degrading enzymes are unable to attack plant cell walls without prior action by a wall-modifying enzyme. Plant Physiol., 46: 69-80.
PubMedDirect Link - Lei, S.P., H.C. Lin, L. Heffernan and G. Wilcox, 1985. Cloning of the pectate lyase genes from Erwinia carotovora and their expression in Escherichia coli. Gene, 35: 63-70.
PubMedDirect Link - Lei, S.P., H.C. Lin, L. Heffernan and G. Wilcox, 1985. Evidence that polygalacturonase is a virulence determinant in Erwinia carotovora. J. Bacteriol., 164: 831-835.
PubMedDirect Link - Nasuno, S. and M.P. Starr, 1967. Polygalacturonic acid trans-eliminase of Xanthomonas campestris. Biochem. J., 104: 178-185.
PubMedDirect Link - Bagherabadi, S., D. Zafari and M.J. Soleimani, 2015. Genetic diversity of Alternaria alternate isolates causing potato brown leaf spot, using ISSR markers in Iran. J. Plant Pathol. Microbiol., Vol. 6, No. 7.
CrossRefDirect Link - Schlee, D., 1975. Review: Numerical Taxonomy. The Principles and Practice of Numerical Classification by P. H. A. Sneath, R. R. Sokal, 1973, W.H. Freeman and Company, San Francisco. Systemat. Zool., 24: 263-268.
CrossRefDirect Link - Chaurasia, A.K. and S. Chaurasia, 2013. Studies on the development of fruit rot of tomato caused by Alternaria solani (Ellis & Mart.) Jones & Grout. Int. J. Pharmacy Life Sci., 4: 2713-2716.
Direct Link - Bauer, W.D., D.F. Bateman and C.H. Whalen, 1977. Purification of an endo-13-1,4 galactanase produced by Sclerotinia sclerotiorum: Effects on isolated plant cell walls and potato tissue. Phytopathology, 82: 1185-1193.
Direct Link - Mankarios, A.T. and J. Friend, 1980. Polysaccharide-degrading enzymes of Botrytis allii and Sclerotium cepivorum. Enzyme production in culture and the effect of the enzyme on isolated onion cell walls. Physiol. Plant Pathol., 17: 93-104.
CrossRefDirect Link - Brown, W., 1965. Toxins and cell-wall dissolving enzymes in relation to plant disease. Ann. Rev. Phytopathol., 3: 1-21.
CrossRefDirect Link - Boccara, M., R. Vedel, D. Lalo, M.H. Lebrun and J.F. Lafay, 1991. Genetic diversity and host range in strains of Erwinia chrysanthemi. Mol. Plant-Microbe Interact., 4: 293-299.
Direct Link