
Mahfut
Department of Biology, Faculty of Mathematics and Natural Sciences, Universitas Lampung, Lampung, Indonesia
Faculty of Agriculture, Universitas Gadjah Mada, Yogyakarta, Indonesia
Tri Joko
Faculty of Agriculture, Universitas Gadjah Mada, Yogyakarta, Indonesia
Budi Setiadi Daryono
Faculty of Biology, Universitas Gadjah Mada, Yogyakarta, Indonesia
Asian Journal of Plant Pathology
Year: 2016 | Volume: 10 | Issue: 1-2 | Page No.: 9-14







ABSTRACT
Orchids are one of the important ornamental plants that were cultivated in tropical countries, including in Indonesia. Virus infections has been important limiting factor in orchids cultivation because it could decrease the orchids quality. Odontoglossum ringspot virus (ORSV) is one of the most reported viruses infecting orchids, which spread widely in the world. During 2010-2011 surveys of viral diseases were conducted in Jawa and Bali, Indonesia. The orchids were found infected by virus, showed symptoms of mosaic, mottle, chlorotic, necrotic, streak, wilting leaf and ringspot on leaf surface. Detection with Reverse Transcription-Polymerase Chain Reaction (RT-PCR) showed that only 5 from 88 samples were infected by ORSV with total incidence of 5.7%. Three leaf samples of Phalaenopsis sp., were infected, then called ORSV BOC, ORSV KRB and ORSV TNBB isolates, respectively. The results obtained by amplification of DNA band with 474 bp in length as expression of Coat Protein (CP) gene. Phylogenetic analysis based on nucleotide sequences of CP gene showed that ORSV BOC have similarity to ORSV Germany, whereas, ORSV KRB and ORSV TNBB lead to speciation that possibly to be a new strain. This study was proved that ORSV have entered and spread widely by infected orchids in orchids landscape (nursery), semi-natural forests (botanical gardens) and natural forest (national park) in Java and Bali, Indonesia.
PDF Abstract XML References Citation
Received: December 17, 2015;
Accepted: February 09, 2016;
Published: March 15, 2016
How to cite this article
Mahfut, Tri Joko and Budi Setiadi Daryono, 2016. Molecular Characterization of Odontoglossum Ringspot Virus (ORSV) in Java and Bali, Indonesia. Asian Journal of Plant Pathology, 10: 9-14.
URL: https://scialert.net/abstract/?doi=ajppaj.2016.9.14
URL: https://scialert.net/abstract/?doi=ajppaj.2016.9.14
INTRODUCTION
Orchids are one kind of plants that have a high aesthetic value (Pant, 2013) and most desirable by communities as ornamental plant (Griesbach, 2002). This plant is commonly found in the tropics (Pant, 2013), including Indonesia (Sandara, 2002). Orchids, more than most cultivated plants, are affected by virus diseases reducing their commercial value considerably in several countries (Pataky, 1990; Koh et al., 2014). Odontoglossum ringspot virus (ORSV) or also called tobbaco mosaic virus orchid strain (TMV-O) belongs to the family Virgaviridae (Ong et al., 2016), being the most prevalent and economically important (Navalinskiene et al., 2005; Zheng et al., 2010). The ORSV cause severe damage to orchid plants and are distributed throughout the world (Ryu and Park, 1995; Zettler et al., 1990; Sherpa et al., 2006).
The ORSV was first isolated from Odontoglossum grande orchid (Jensen and Gold, 1951). The virus particle has elongated, 300×18 nm rigid rod-shape, which are non-enveloped, similar with that of Tobacco Mosaic Virus (TMV). The virion contains a single molecule of positive sense linear ssRNA, 6 kb in size (Choi et al., 2002). The virus is mechanically and wound transmissible. The complete nucleotide sequence of ORSV RNA genome have been reported by Ryu and Park (1995). The 3’-terminal region encoded for Coat Protein (CP) was found highly conserved among various kinds of orchids cultivated in Asia. Some studies have demonstrated that ORSV caused streak or stripped mosaic, diamond mottle, or ringspot on leaves (Corbett, 1967; Matthews, 1991), also crinkle symptom (Zheng et al., 2010), color break, malformation, distortion and necrotic streak on petals (McMillan and Vendram, 2005; Lakani et al., 2010).
This virus was firstly discovered in the United States (Corbett, 1967) and has spread to other countries including Indonesia since Suseno (1976). Based on the results of field surveys, ORSV has a wide distribution area that were in Java, Ujung Pandang and Bali (Inouye and Gara, 1996). All this time, previous studies were only done on the orchids in the nursery or in semi-natural forests. Nevertheless, it was needed to study the orchids in the nursery, semi-natural forests and natural forests to find out appropriate management strategies of the disease. Survey and samples collection of this study were conducted in seven locations of orchids cultivation in Java and Bali, Indonesia.
MATERIALS AND METHODS
Virus source: Samples were collected during May-August, 2010 and April-June, 2011 from 7 locations cultivating orchids in Java and Bali: Purwodadi botanical garden (East Java), Wonosadi forest (Yogyakarta), Cibodas botanical garden (West Java), Bogor botanical garden (Bogor), West Bali national park (Bali), Mekar Lestari nursery (Yogyakarta) and Borobudur orchids center (Central Java). Samples collected were the orchid leaves showing symptoms of ORSV infection.
Molecular detection: Total RNA was extracted from a 0.1 g symptomatic leaves of samples using total RNA isolation kit (SBS Genentech Co., Ltd., China). Total RNA was used as a template for complementary DNA (cDNA) construction by using the Titan One Tube RT-PCR Kit (Roche Applied science, USA). The RT-PCR was carried out by using a pair of Coat Protein (CP) gene primers of ORSV CP gene; ORSV forward primer CP-F1 (5'-ATGTCTTACACTATTACAGACCCG-3') and reverse primer ORSV CP-R1 (5'-GGAAGAGGTCCAAGTAAGTCC-3') (Lee and Chang, 2006). The PCR conditions for DNA amplification were 35 cycles at 95°C for 30 sec, 50°C for 45 sec, 72°C for 1 min and post-elongation at 72°C for 10 min. Bands of expected size 474 bp DNA product was observed on 2% agarose gel containing ethidium bromide 0.1% under UV transilluminator.
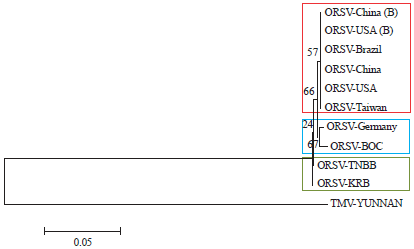
Phylogenetic analysis: Previously, the results with RT-PCR detection showed that ORSV infected three leaf samples of Phalaenopsis sp., then called ORSV BOC, ORSV KRB and ORSV TNBB isolates, respectively. The RT-PCR product samples were then subjected to nucleotide sequencing at 1st BASE company, Singapore. Nucleotide sequence results was then analyzed and compared using DNASTAR Lasergene software Version 3.0.25 DM and subjected to BLAST searches to identify related sequences available from the DDBJ database. The 11 obtained sequences of BLAST search were categorized as tobamovirus. Data were aligned with CLUSTAL W to assemble the ORSV CP gene and tobacco mosaic virus (TMV-YUNNAN) was used as the out group. Phylogenetic trees were obtained from the clustering of data by the Neighbour Joining (NJ) in MEGA version 4.0 software. The strength of the internal branches from the resulting tree was statistically tested by bootstrap analysis from 0:05 bootstrap replications.
RESULTS
Isolation of ORSV: Total orchid leaf samples were 88 samples that collected from 27 genus. Samples were commonly found from genus Phalaenopsis (5 locations) and Coelegyne (5 locations). The infected orchids showed variation of symtomps such as mosaic, necrotic, chlorotic, streak, mottle and ringspot on the upper part of the leaf and wilting leaf (Fig. 1a-h).
 | |
| Fig. 1(a-h): | Symptomps variation of samples collection infected by viruses (a) Necrosis, (b) Chlorotic, (c) Necrotic, (d) Streak, (e) Wilting leaf, (f) Necrotic dan necrosis, (g) Ringspot and (h) Mottle |
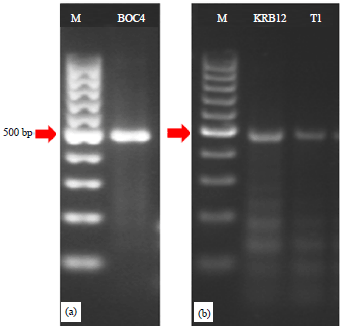 | |
| Fig. 2(a-b): | The RT-PCR detection of ORSV with specific primer of CP gene, (a) Lane 2; BOC4 (ORSV BOC isolate from nursery) and (b) Lane 2; KRB12 (ORSV KRB isolate from semi-narural forest), Lane 3; T1 (ORSV TNBB isolate from natural forest) and M: Marker molecular weight 100 bp DNA ladder (rainbow) |
Molecular detection: Detection with Reverse Transcription-Polymerase Chain Reaction (RT-PCR) showed that ORSV infected five leaf samples of Liparis sp. (W2), Dendrobium salacence (KRP20), Phalaenopsis amabilis (KRB12) and Phalaenopsis sp. (BOC4 and T1). The RT-PCR comfirmed the existence of ORSV coat protein gene that is 474 bp of its DNA (Fig. 2a-b).
Of total 88 samples of orchid leafs, only 5 samples were infected by ORSV based on RT-PCR detection, with a total incidence of 5.7%.
 | |
| Fig. 3: | Comparison of nucleotides sequence the the Coat Protein (CP) gene between isolates ORSV-BOC, ORSV-KRB, ORSV-TNBB and other ORSV isolate |
| Table 1: | Detection of ORSV in cultivated orchids in Java and Bali, Indonesia |
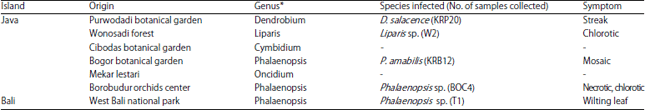 | |
| *Genus shown were representative of orchids found (total collecting orchids samples from 27 genus) | |
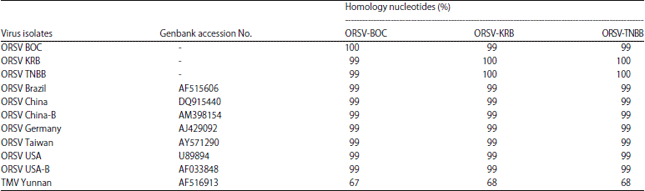
| Table 2: | Homology of nucleotides sequence identities of CP gene between ORSV-BOC, ORSV-KRB, ORSV-TNBB and other ORSV isolates |
 | |
Thus, it was concluded that Phalaenopsis is orchids genus which most abundantly infected by ORSV in Indonesia (Table 1).
Phylogenetic analysis: Phylogenetic analysis of the DNA compared with those of other ORSV showed highest homology corresponding CP gene of seven isolates ORSV on Genbank (99-100%), with the highest nucleotide sequence similarity to the 461 bases (Table 2). Result of phylogenetic tree for the CP gene analyzes ORSV BOC are displayed on the same branching with ORSV Germany, whereas ORSV KRB samples and TNBB on a separate branch of the possible second isolate is distinct strains with different pre-existing strains (Fig. 3).
DISCUSSION
The BOC4, KRB12 and T1, each were used as representative of the ORSV isolates from the orchids landscape (nursery), semi-natural forest (botanical garden) and natural forest (national park), respectively. The results proved that ORSV has been infecting natural as well as cultivated orchids in Java and Bali, Indonesia.
The ORSV capable of infecting orchids naturally in natural forests, that is spread only through contaminated soil or water. Meanthough, the ORSV infects orchids in semi-natural forest and nursery through the means as already mentioned, as well as through human aid intermediaries, such as mechanical inoculation and contamination of equipment used for separation plants for vegetative propagation and harvesting flowers (Zheng et al., 2010; Koh et al., 2014).
Information on the orchids infecting ORSV in Indonesia is still very limited. Previously, Inouye and Gara (1996) first reported the influx of ORSV in Indonesia by means of detection and identification of viruses that infected orchids in Indonesia. The sampling were carried out on 12 nursery in East Jakarta, Bogor, Sukabumi, Yogyakarta, Ujung Pandang and Bali. Furthermore, ORSV infection were reported in Mount Sindur, West Java (Isnawati, 2009; Syahierah, 2010), Bogor, Magelang and Malang (Lakani et al., 2010; Lakani, 2011), Bandung and Surabaya (Lakani, 2011). The host range of ORSV in Indonesia has been able to infect a variety of orchids including bulbophyllum, calanthe, cattleya, oncidium (Inouye and Gara, 1996; Isnawati, 2009), coelogyne, grammatophyllum (Syahierah, 2010), phalaenopsis (Inouye and Gara, 1996; Syahierah, 2010) and dendrobium (Syahierah, 2010; Lakani et al., 2010). This suggests that the infection is severe in Indonesia with proven ability of ORSV infection in various genera of orchids and patterns of infection that has spread almost the entire territory of the country.
In addition, based on the observation the symptoms found in conditions of severe infection is in the form of streaks and ringspot, which are typical symptoms of ORSV infection. This suggests that infection of ORSV to orchids in Indonesia has been on vulnerable stage. Therefore, ORSV infection should receive more attention as much as possible in order to reduce the severity of the disease and to prevent the spreading of this virus. For protection, it is also important to use sterilized and virus free equipment for the vegetative propagation and harvesting orchid flower. Furthermore, Indonesia requires a strict law enforcement to rule the mobility of imported orchids from abroad, thus prevent entry of disease-carrying materials into Indonesia.
The ORSV is the most economically important virus (Sherpa et al., 2006; Rao et al., 2015; Navalinskiene et al., 2005; Ajjikuttira and Wong, 2009) and this study provides evidence of three isolates ORSV from Java and Bali, Indonesia. Based on part of CP nucleotide, it was showed that ORSV isolates from Java and Bali were closely related to the other ORSV isolates from several Asian countries, Europe and America with the index similarity nucleotide sequences CP gene 99-100%. The results are consistent with the previously reported (Lakani et al., 2010). Phylogenetic analysis of the CP nucleotide confirmed the genetic relationship of ORSV from Java and Bali isolates with those corresponding isolates. The ORSV BOC isolate was shown on the same branching as ORSV isolate from Germany. It suggested that ORSV BOC isolate might be originally from one of those countries, which was spread via vegetative materials. Germany is an exporter of orchid seeds and plants to Indonesia in 1997-2001 (BPPP., 2002). The German state ranks 14th, while the biggest exporters country are Taiwan, followed by Thailand, the Netherlands, Japan and United States.
CONCLUSION
Isolates of ORSV KRB and TNBB on separate branches and not mixed with other ORSV isolates having undergone many mutations that led to speciation becoming a pure isolates from Indonesia. Speciation of ORSV KRB and TNBB isolates might be caused by the presence of mutation, which occurred in the CP gene bases. Mutations that occur are insertion, deletion and substitution. There are changes in amino acid compositions and lead to lower similarity of amino acid sequence compared with the other isolates. These might occur due to natural adaptation of virus to Indonesia environment.
ACKNOWLEDGMENT
This study was funded by Indonesia-Managing Higher Education for Relevance and Efficiency (IM-HERE) research grant for 2010, No.: UGM/BI/1628/I/05/04. The authors also would like to express gratitude to Rizko Hadi for kindly doing language review of this manuscript.
REFERENCES
- Choi, S.K., S.H. Choi, K.H. Ryu, C.W. Choi, J.K. Choi and W.M. Park, 2002. Identification and characterization of a ringspot isolate of Odontoglossum ringspot virus from Cymbidium var. 'Grace Kelly'. Plant Pathol. J., 18: 317-322.
Direct Link - Inouye, N. and I.W. Gara, 1996. Detection and identification of viruses of Orchids in Indonesia. Bull. Res. Inst. Bioresour., 4: 109-118.
Direct Link - Koh, K.W., H.C. Lu and M.T. Chan, 2014. Virus resistance in orchids. Plant Sci., 228: 26-38.
CrossRefDirect Link - Lakani, I., G. Suastika, N. Mattjik and T.A. Damayanti, 2010. Identification and molecular characterization of Odontoglosum Ringspot Virus (ORSV) from Bogor, Indonesia. Hayati J. Biosci., 17: 101-104.
Direct Link - Lee, S. and Y.C. Chang, 2006. Multiplex RT-PCR detection of two orchid viruses with an internal control of plant nad 5 mRNA. Plant Pathol. Bull., 15: 187-196.
Direct Link - McMillan, Jr., R.T. and W.A. Vendrame, 2005. Color break in orchid flowers. Proc. Florida State Hortic. Soc., 118: 287-288.
Direct Link - Navalinskiene, M., J. Raugalas and M. Samuitiene, 2005. Viral diseases of flower plants: 16. Identification of viruses affecting orchids (Cymbidium Sw.). Biologia, 2: 29-34.
Direct Link - Ong, J.W.L., R.D. Phillips, K.W. Dixon, M.G.K. Jones and S.J. Wylie, 2016. Characterization of the first two viruses described from wild populations of hammer orchids (Drakaea spp.) in Australia. Plant Pathol., 65: 163-172.
CrossRefDirect Link - Pant, B., 2013. Medicinal orchids and their uses: Tissue culture a potential alternative for conservation. Afr. J. Plant Sci., 7: 448-467.
CrossRefDirect Link - Rao, X., Y. Li, J. Sun, X. Li, M. Li and M. Xiang, 2015. Genetic diversities of Cymbidium mosaic virus and Odontoglossum ringspot virus isolates based on the coat protein genes from orchids in guangdong province, China. J. Phytopathol., 163: 324-329.
CrossRefDirect Link - Ryu, K.H. and W.M. Park, 1995. The complete nucleotide sequence and genome organization of odontoglossum ringspot tobamovirus RNA. Arch. Virol., 140: 1577-1587.
CrossRefDirect Link - Sherpa, A.R., T.K. Bag, V. Hallan and A.A. Zaidi, 2006. Detection of Odontoglossum ringspot virus in orchids from Sikkim, India. Aust. Plant Pathol., 35: 69-71.
CrossRefDirect Link - Zheng, Y.X., B.N. Shen, C.C. Chen and F.J. Jan, 2010. Odontoglossum ringspot virus causing flower crinkle in Phalaenopsis hybrids. Eur. J. Plant Pathol., 128: 1-5.
CrossRefDirect Link - Zettler, F.W., N.J. Ko, G.C. Wisler, S. Elliot Mark and S.M. Wong, 1990. Viruses of orchids and their control. Plant Dis., 74: 621-626.
CrossRefDirect Link