
M.L.M.C. Dissanayake
Department of Export Agriculture, Faculty of Agricultural Sciences, Sabaragamuwa University of Sri Lanka, Belihuloya, Sri Lanka
Shin-Ichi Ito
Department of Biological and Environmental Sciences,
Yoshihiko Akakabe
Department of Biological Chemistry, Faculty of Agriculture, Yamaguchi University, 1677-1 Yoshida, Yamaguchi, Yamaguchi, 753-8515, Japan
Asian Journal of Plant Pathology
Year: 2015 | Volume: 9 | Issue: 1 | Page No.: 16-26







ABSTRACT
Plant based botanical fungicides are one of the most important alternative for hazardous fungicides. Antifungal activity (AFA), identification and understanding the mechanism of action of predominant antifungal compounds present in medicinal plant (Acorus calamus L) were carried out in this study. The antifungal activity and both n-butanol extract and purified antifungal compound of A. calamus (L) was evaluated against five fungal strains (Fusarium oxysporum, Fusarium solani, Colletotrichum gloesporioides, Botrytis cinerea and Alternaria solani) by using agar overlay techniques and bioautography. Isolation and screen of antifungal compounds (AFC) was carried out by Thin Layer Chromatography (TLC) and bioautography. The AFC were elucidated and identified by using gas chromatography-mass spectrometry. The mode of action of purified AFC from A. calamus was studied by Evans blue staining and generation of Reactive Oxygen Species (ROS). Both n-butanol extracts and purified AFC exhibited different degrees of antifungal activity against all fungal pathogens tested. Both n-butanol extract from leaf and rhizome of A. calamus and purified AFC showed prominent one major spot on TLC after detection with p-anisaldehyde. In bioautographic technique, clear inhibition zones at same distance of TLC spot was observed against all fungal strains examined. The major purified antifungal compounds were identified as a β-asarone (94.3%) and α-asarone (3.4%). Fungal cells treated with different concentration of purified AFC showed a rapid production of ROS. The extent of dead cells stained with Evans blue dye seemed to correlate with the level of ROS production. These results suggested both n-butanol extracts and purified compounds from A. calamus show antifungal activity against plant pathogenic fungi and main compound attributed for antifungal activity is β-asarone. Furthermore, AFC caused damages to the cell membrane, leading to altering cell morphology and eventually cell death. Thus A. calamus extract has potential application as an alternative or integrated method to chemical control on plant diseases caused by five different plant pathogens examined.
PDF Abstract XML References Citation
Received: November 17, 2014;
Accepted: January 14, 2015;
Published: March 26, 2015
How to cite this article
M.L.M.C. Dissanayake, Shin-Ichi Ito and Yoshihiko Akakabe, 2015. TLC Bioautography Guided Detection and Biological Activity of Antifungal Compounds from Medicinal Plant Acorus calamus Linn. Asian Journal of Plant Pathology, 9: 16-26.
URL: https://scialert.net/abstract/?doi=ajppaj.2015.16.26
URL: https://scialert.net/abstract/?doi=ajppaj.2015.16.26
INTRODUCTION
The intensive and frequent applications of fungicides are often required to control fungal pathogens causing devastated diseases of crops. The great public awareness of environmental and health issues as well as development of fungicide resistance in plant pathogens against synthetic fungicide has stimulated an increasing demands to shift away from the reliance on conventional fungicides. Plant based botanical extracts either as pure compounds or as standardized extracts provide an enormous bio resource of potential use as antifungal compounds due to their antimicrobial activities (Sasidharan et al., 2011; Cos et al., 2006; Fabricant and Farnsworth, 2001).
The common names of Acorus calamus Linn (family Araceae) are “Sweet flag” or “Waan-Nam” and native to central Asia and Eastern Europe (Gilani et al., 2006). It is a perennial, semi aquatic and smelly plant found both temperate and sub temperate and well known medicinal plant in traditional medicine in several countries (Mehrotra et al., 2003; Caius, 1986; Shin-Chen, 1973; Duke and Ayensu, 1985). Acorus calamus plants have been used in herbal medicine to cure fever, for asthmas, bronchitis and as an all-round sedative (Leung, 1980). Extract of rhizome and root and essential oil from this plant have been reported to possess several important biological activity including antifungal (Lee, 2005; Lee et al., 2004) antibacterial (McGaw et al., 2002; Phongpaichit et al., 2005) allelopathic (Nawamaki and Kuroyanagi, 1996), anticellular and immunosuppressive (Mehrotra et al., 2003) properties. However very few study has been done to investigate its potential use as antifungal activity against plant pathogenic fungi (Suvarna et al., 2011; Singh et al., 2010; Mungkornasawakul et al., 2002). Previous study (Dissanayake and Jayasinghe, 2013; Dissanayake, 2015) shows methanol extract of sweet flag were highly effective against pathogenic fungi Rhizoctonia solani, Colletotrichum musae, Fusarium oxysporum and Fusarium proliferatum.
Despite the wide scale use of sophisticated HPLC and GC techniques, HPLC bioassays and hyphenated methods. TLC is still proving its worth as a simple, inexpensive and robust means for the chemical and biological screening of plant extracts, with subsequent activity-guided isolation of natural products (Hostettmann, 1991). In an effort to discover new antimicrobial compounds, many research studies have been screened plant extracts with different biological activities. In this regard, several bioassays were developed for screened antimicrobial properties of plant extracts (Hostettmann et al., 2001). Out of those techniques, bioautography is a highly efficacious assay for the detection of antimicrobial compounds because it allows localization of activity even in a complex matrix and therefore facilitates the target-directed isolation of the active constituents (Rahalison et al., 1991). Bioautography facilitate rapid and precise detection of new antimicrobial compounds from plants and other natural products. Especially this technique allows the localization of antimicrobial activity directly on a chromatographic plate where the organism is applied (Navarro et al., 1997). The method is fast, cheap and permits a better bioassay-directed fractionation of bioactive compounds (Hamburger and Cordell, 1987). Bioautography is particularly important to avoid the time-consuming isolation of inactive compounds.
Thus, the objectives of this study were to (1) Evaluate the antifungal activity of n-butanol extract of both rhizome and leaves of A. calamus (Sweet flag) against range of plant pathogenic fungi of significant importance in plant diseases, (2) Identify the antifungal compounds that contribute to the antifungal activity of both rhizome and leaves of A. calamus and (3) Understand the mechanism of action of the predominant antifungal compound present in A. calamus leaves and rhizomes.
MATERIALS AND METHODS
Sample collection and extraction: Both rhizome and leaves of A. calamus (Sweet flag) were collected from surrounding areas of Belihuloya, Sri Lanka. The dried plant materials were milled to a fine powder using grinder and stored in the dark at room temperature (25°C) in airtight containers until utilized at the Laboratory of Molecular Plant Pathology, Department of Biological and Environmental Sciences Faculty of Agriculture, Yamaguchi University, Japan. Dried samples were ground and defatted with hexane three times. The defatted sample was extracted with methanol three times. Combined methanol extracts were suspended into water and the suspension was then partitioned with n-butanol to solubilize antifungal compounds in the organic phase, leaving sugars, amino acids and salty compounds in the water phase. The n-butanol extract was vacuum dried and the resultant residue was used for further study.
Fungal isolates: Five fungal pathogens of significant important in plant diseases; Fusarium oxysporum, Fusarium solani, Colletotrichum gloesporioides, Botrytis cinerea and Alternaria solani were obtained from Laboratory previously mentioned fungi, those were cultured on Potato Dextrose Agar (PDA) (39 g of PDA, was suspended in 1 L of distilled water).
Assay of antifungal activity: Both rhizome and leaves of A. calamus extract were tested for their efficiency against the pathogen by using an agar dilution technique (Jorgensen et al., 1999). Different concentrations of the extracts; 25, 12.5, 6.25 and 3.12% were obtained by amending PDA. The amended medium was dispensed into sterile petri plates and allowed to solidify with streptomycin (100 μg mL-1). Four milliliter diameter mycelia discs of each fungus were inoculated on each amended agar plate. Inoculated plates were incubated at 25°C and radial growths of each fungal growth were recorded for 7 days. Each treatment was replicated three times with appropriate untreated controls. Then all the plates were incubated at 25°C in dark condition. The percentage of growth inhibition was calculated according to the following equation (Vincent, 1947):
where, C is the colony diameter of the mycelium on the control plate (mm) and T is the colony diameter of the mycelium on the treated plate (mm).
Thin layer chromatography of Acorus calamus: Chemical constituents of the A. calamus extracts were separated on aluminium-backed Thin Layer Chromatography (TLC) plates (silica gel 60 F 254, 20x20 cm, layer thickness 0.25 mm, particle size of 10-20 μm; Merck, Darmstadt, Germany). The TLC plates were developed under saturated conditions with system developed in our laboratory, i.e., chloroform/methanol/water (6:3:0.5) and separated chemical compounds were detected using acidified p-Anisaldehyde (5.3 mL p-Anisaldehyde: 100 mL ethanol: 1 mL sulphuric acid) as a spray. After spraying, the chromatograms were heated at 110°C in an incubator to allow for optimal colour development.
Thin-layer chromatography-direct bioautography: Ten microliter (0.1 mg mL-1) of A. calamus extract were loaded onto TLC plates in a narrow band and eluted using the chloroform/ methanol/water (6:3:0.5) solvent systems. The developed plates were dried under a stream of fast moving air for 3 days to remove traces of solvent on the plates.
All fungal strains were shake-incubated separately in potato dextrose broth (Becton Dickinson, Sparks, MD, USA) at 25°C for 7 days. The culture was filtered through three layers of gauze cloth to remove mycelia. Spores of the each fungus were obtained by centrifugation (3000xg, 10 min). Dehydrated Potato Dextrose Agar (PDA) was used to prepare an immersion of spores to spray on TLC plates. Fifteen milliliter of PDA solution was allowed to cool down. As soon as the temperature reached 48°C, spores were added to the PDA to give a concentration of approx. About 1x106 spores/mL for a 20/20 cm plate for each fungal strains F. oxysporum, F. solani, C. gloesporioides, B. cinerea and A. solani. The PDA media amended with spores of respective fungi was sprayed on TLC plates rapidly to avoid immediate solidification.
Isolation of antifungal compounds: The A. calamus extract were applied to Thin Layer Chromatography (TLC) plates (silica gel 60 F254, 20x20 cm, layer thickness 0.25 mm, particle size of 10-20 μm; Merck, Darmstadt, Germany), developed in chloroform/methanol/water (6:3:0.5) as the mobile phase and dried. After development, TLC plates were examined under ultraviolet light and spots marked on the plate were scraped off separately. Compound was extracted from the silica gel with 80% ethanol and the tube containing AFC was centrifuged at 12000xg for 10 min. The resulting supernatant was vacuum-dried to obtain the dried AFC having a white appearance. The dried AFC were examined for purity by TLC and subjected to antifungal activity tests as described above with three different concentrations (100, 300 and 500 μg mL-1).
Identification of bioactive compound: The spot on the TLC plate, which showed high antifungal activity, was subjected to structural analysis. The structure of the compound with antifungal activity was elucidated and confirmed using Nuclear Magnetic Resonance spectroscopy (NMR). The isolated compounds were identified using a Shimadzu QP-5050 GC-MS (Shimadzu, Kyoto, Japan) equipped with a capillary column of DB-WAX (0.25 mm i.d.x60 m; film thickness, 0.25 mm). The oven temperature was programmed to increase from 50-230°C at a rate of 2°C min-1. The injection port and ionizing source were kept at 240 and 230°C, respectively. Helium was the carrier gas, at a flow rate of 1.5 mL min-1 and ionization voltage was 70 eV.
Detection of reactive oxygen species in fungal cells: Fusarium oxysporum was shake-incubated in Potato Dextrose Broth (PDB) (Becton Dickinson, Sparks, MD, USA) at 25°C for 7 days. The culture was filtered through three layers of gauze cloth to remove mycelia. Conidial cells of the fungus were obtained by centrifugation (3000 g, 10 min) of the supernatant. Conidial cells suspended in PDB (1x106 cells mL-1) were shake-incubated at 25°C for 24 h. A 500 μL aliquot of the F. oxysporum cell suspension was mixed with different concentration of AFC (100, 300 and 500 ppm) dissolved in 80% (v/v) ethanol and the mixture was incubated for 3 h. Dead cells were detected by Evans blue staining under a light microscope. Generation of ROS in the fungal cells exposed to AFC was determined by monitoring the conversion of nonfluorescent dihydrorhodamine 123 (DHR123; Sigma-Aldrich) to fluorescent rhodamine 123 using a fluorescent microscope (BZ-9000, Keyence, Osaka, Japan).
Statistical analysis: The experiment was conducted using a completely randomized design. Standard errors of means of three replicates were computed using computer software Microsoft Excel. Analysis of variables (ANOVA) was used to analyze the recorded data. Statistical Analyzing Software (SAS) 9.0 was used to analyze the data. The mean separation was done by Duncan’s multiple range test at p<0.05.
RESULTS
Assay of antifungal activity: The n-butanol extracts from the both leaves and rhizomes of A. calamus showed growth inhibition activity against all fungal isolates examined, although the extents of growth inhibition were different among the fungal stains. The different concentrations of the purified AFC (100, 300, 500 μg mL-1) were evaluated by in vitro inhibition assay of mycelia growth on petri dishes. The purified AFC showed 100% mycelium growth inhibition for B. cinerea, C. gloesporioides and A. solani in the culture media amended with plant extract at 500 μg mL-1. Whereas purified AFC from A. calamus showed moderate inhibition of mycelia growth of F. solani (67%) and F. oxysporum (55%) (Fig. 1).
Thin layer chromatography: Thin Layer Chromatography (TLC) separation of n-butanol extracts from leaves showed an eight spots (1-8). Whereas n-butanol extracts from rhizome of A. calamus showed four spots (3, 6, 7 and 8) (Fig. 2a). However TLC of n-butanol extracts prepared from rhizome and leaves from A. calamus consistently showed one main discrete spot (Rf = 0.7) in gross chemical composition and activity. Chemical compounds relevant to spots (1-8) were recovered from the TLC plates under UV illumination and subjected to antifungal activity test. The antifungal activity test revealed that only compound recovered from spots 8 (Rf = 0.7) had the predominant antifungal activity against fungal stains examined. The TLC separation of purified AFC consistently showed one main discrete spot with same Rf value (Fig. 2b). During development of TLC methodology a number of solvent systems were trialed and a mixture of chloroform/methanol/water (6:3:0.5) was eventually found to produce significant degree of separation of active compounds. The anisaldehyde-acid spray reagent was chosen as it was seen to detect a wider range of compounds.
Thin-Layer Chromatography (Direct bioautography): Inhibitory activity following TLC separation of n-butanol extracts was found to be attributed by the presence of only one major constituents spots 8 (Rf = 0.7) and gave clear zones of inhibition of fungal growth corresponding to the positions producing large inhibitory zones at Rf 0.7 for each fungal strains F. oxysporum, F. solani, C. gloesporioides, B. cinerea and A. solani (Fig. 3).
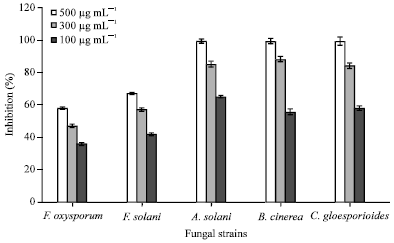 | |
| Fig. 1: | Mycelium growth inhibition of plant pathogenic fungi Fusarium oxysporum, Fusarium solani, Alternaria solani, Botrytis cinerea and Colletotrichum gloesporioides with different concentration of purified AFC |
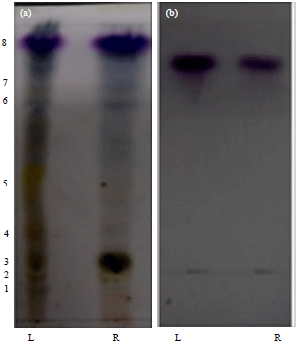 | |
| Fig. 2(a-b): | (a) TLC profile of n-butanol extracts from the both Leaves (L) and Rhizomes (R) of Acorus calamus and (b) TLC profile of purified antifungal compound |
 | |
| Fig. 3(a-f): | (a) TLC profile of purified antifungal compound and Bioautography of purified AFC from Acorus calamus for fungal strains of, (b) F. oxysporum, (c) F. solani, (d) C. gloesporioides, (e) B. cinerea and (f) A. solani. White areas indicate occurrence of zones of inhibition of fungal growth |
Identification of bioactive compound: The antifungal compound corresponding to inhibition zone of spots 2 were further purified by TLC. The structure of the compound with antifungal activity was elucidated and confirmed using Nuclear Magnetic Resonance spectroscopy (NMR).
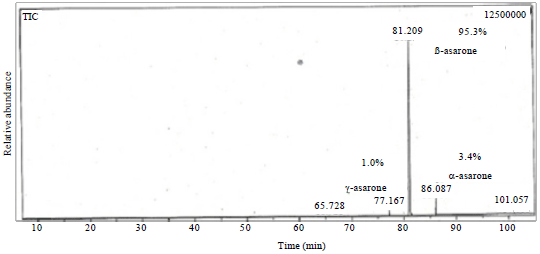 | |
| Fig. 4: | Gas chromatography-mass spectrometry data of purified AFC from A. calamus |
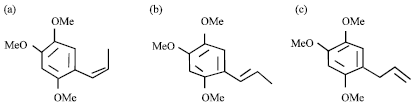 | |
| Fig. 5(a-c): | Structure of (a) β-asarone (cis-2, 4, 5-trimethoxy-1-propenylbenzene), (b) α-asarone (trans-2,4,5-trimethoxy-1-propenylbenzene) and (c) γ-asarone (1-allyl-2,4,5-trimethoxybenzene) as deduced from NMR spectroscopy |
According to Gas Chromatography-Mass Spectrometry analysis spot was identical to those of β-asarone (94.3%), α-asarone (3.4%) and γ-asarone (1%) (Fig. 4). The chemical structure of β-asarone (cis-2, 4, 5-trimethoxy-1-propenylbenzene), α-asarone (trans-2, 4, 5-trimethoxy-1-propenylbenzene) and γ-asarone (1-allyl-2, 4, 5-trimethoxybenzene) as deduced from NMR spectroscopy (Fig. 5).
Antifungal mode of action of β-asarone: β-asarone showed fungicidal activity in a dose-dependent manner (Fig. 6a) of the plant pathogenic fungi in PDB. Fusarium oxysporum cells treated with β-asarone were stained with DHR 123. Fungal cells treated with 500 ppm AFC, at which most fungal cells were killed by the compound, showed a rapid production of ROS (Fig. 6b). The extent of dead cells stained with Evans blue dye (Fig. 6a) seemed to correlate with the level of ROS production (Fig. 6b).
DISCUSSION
Although many studies (Ganjewala and Srivastava, 2011; McGaw et al., 2002; Rani et al., 2003; Phongpaichit et al., 2005) have investigated importance of biological activity, especially antimicrobial properties of A. calamus against human pathogens, very few studies (Suvarna et al., 2011; ; Mungkornasawakul et al., 2002) have been carried out to find its potential as antifungal agents against plant pathogens.
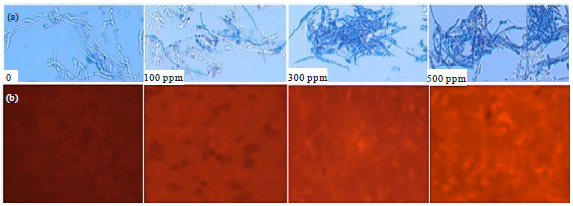 | |
| Fig. 6(a-b): | Fungicidal action of purified AFC from Acorus calamus at different concentrations (a) Evans blue staining of F. oxysporum exposed to different concentrations (0, 100, 300 and 500 ppm) of purified AFC and (b) Epifluorescence microscopy F. oxysporum cells exposed to different concentrations (0, 100, 300 and 500 ppm) of purified AFC for 60 min |
In this study, the antifungal activity of n-butanol extracts was tested from both leaves and rhizomes of A. calamus against phytopathogenic fungal strains of F. oxysporum, F. solani, C. gloesporioides, B. cinerea and A. solani. Both Fusarium fungal strains need higher concentration to inhibition of mycelium growth in in vitro. Similar results have been found in previous studies (Suvarna et al., 2011). Present study revealed n-butanol extracts from the both leaves and rhizomes of A. calamus has ability to impair the mycelia growth of all tested fungal stains confirming its fungicidal properties.
In this study, for first time we attempted to identify main active ingredient responsible for antifungal activity of A. calamus plant parts using TLC-Bioautography followed by gas chromatography-mass spectrometry analysis. According to present study, TLC bioautography-guided strategy confirmed that one major compound (spot 8) is significant contribution to the overall antifungal activity. According to Gas chromatography-mass spectrometry analysis the major antifungal compound was identified as asarone with their different isomeric forms β-asarone (94.3%) α-asarone (3.4%) and γ-asarone (1%) from A. calamus. β-asarone was seem to comprise a large proportion of extractable materials and was found to be principally responsible for antifungal activity in extracts of A. calamus. Many studies (Devi and Ganjewala, 2009; Venskutonis and Dagilyte, 2003; Rani et al., 2003; Perrett and Whitfield, 1995) reported that the phenylpropanoids β-asarone and α-asarone found in leaf, rhizomes and root are responsible for entire antimicrobial activity Acorus calamus. It has also been reported occurring in A. gramineus (Lee et al., 2004). However, no simple method is available for separation and isolation of asarones (alpha, beta and gamma) in single isomeric form due to their similar physical properties. So far, HPLC and GC-MS are two most frequently used techniques for the analysis of asarone. According to our findings showed TLC bioautography followed by gas chromatography-mass spectrometry analysis was highly applicable for identification of target-detected isolates of antifungal compound even up to isomeric forms. Results presented in this study about TLC fractions separation and contain of β-asarone and α-asarone in purified AFC is completely matches with those reported by Devi et al. (2013). TLC-bioautography method is very useful in isolating compounds with antimicrobial activity because the Rf of the active compound can be used in bioassay guided fractionation instead of requiring labour intensive determination of activity of fractions.
The proportion of each chemical compound in the essential oil varies among the varieties of A. calamus, corresponding to the ploidy level of the taxon (Mazza, 1985; Rost and Bos, 1999). β-asarone concentrations vary markedly among the oils from the three varieties. The tetraploid plant oil is high in β-asarone (90-96%) and also contains α-asarone. The triploid plants contain a small portion of β-asarone (5%) in their oil and the diploid plants lack β-asarone but contain high amounts of geranyl acetate. Therefore according to our findings, A. calamus existing in Sri Lanka could be tetraploid.
Fusarium oxysporum cell suspension treated with purified AFC showed rapid cell death with increases of concentration and correlate with the level of ROS production. In the present study, that ROS was also produced in F. oxysporum cells treated with purified AFC. Interestingly, ROS production levels coincided with the extent of cell death, suggesting involvement of ROS production in the fungicidal action of purified AFC. Loss of membrane integrity in the fungal cells exposed to purified AFC was suggested by the increased uptake of the fluorescent dye, propidium iodide. ROS production occurs in fungal cells treated with α-tomatine, a spirostanol saponin of tomato (Ito et al., 2007). In the present study it was found that ROS was also produced in F. oxysporum cells treated with purified AFC from A. calamus. Interestingly, ROS production levels coincided with the extent of cell death, suggesting involvement of ROS production in the fungicidal action of AFC from A. calamus. These results suggest that purified AFC causes damage to the cell membrane, leading to altering cell morphology and eventually cell death. Based on Scanning Electron Microscopy (SEM) study (Phongpaichit et al., 2005) also reported β-asarones altered the fungal hyphal morphology with collapsed form.
The n-butanol extracts of A. calamus as well as purified compounds show antifungal activity and main compound attributed for antifungal activity is β-asarone. AFC causes damage to the cell membrane, leading to altering cell morphology and eventually cell death. These indicate potential application of A. calamus extract as an alternative or integrated method to chemical control of plant pathogens examined. However, prior to their use as botanical fungicide, toxicity tests against humans and other animals need to be conducted.
REFERENCES
- Cos, P., A.J. Vlietinck, D.V. Berghe and L. Maes, 2006. Anti-infective potential of natural products: How to develop a stronger in vitro proof-of-concept. J. Ethnopharmacol., 19: 290-302.
CrossRefDirect Link - Devi, A.S., Sudhasini and S. Babu, 2013. Purification of beta Asarone from Acorus calamus L. Res. J. Pharmaceut. Biol. Chem. Sci., 4: 1279-1284.
Direct Link - Devi, S.A. and D. Ganjewala, 2009. Antimicrobial activity of Acorus calamus (L.) rhizome and leaf extract. Acta Biologica Szegediensis, 53: 45-49.
Direct Link - Dissanayake, M.L.M.C. and J.A.N. Jayasinghe, 2013. Antifungal activity of selected medicinal plant extracts against plant pathogenic fungi: Rhizoctonia solani, Colletotrichum musea and Fusarium oxysporum. Int. J. Sci. Inventions Today, 2: 421-431.
Direct Link - Dissanayake, M.L.M.C., 2015. Identification of causative pathogen of flower bud wilt disease in Dendrobium sp. and in-vitro growth inhibition by medicinal plant extract. Int. J. Plant Soil Sci., 4: 132-139.
CrossRefDirect Link - Fabricant, D.S. and N.R. Farnsworth, 2001. The value of plants used in traditional medicine for drug discovery. Environ. Health Perspect., 109: 69-75.
CrossRefPubMedDirect Link - Ganjewala, D. and A.K. Srivastava, 2011. An update on chemical composition and bioactivities of Acorus species. Asian J. Plant Sci., 10: 182-189.
CrossRefDirect Link - Gilani, A.U., A.J. Shah, M. Ahmad and F. Shaheen, 2006. Antispasmodic effect of Acorus calamus Linn. is mediated through calcium channel blockade. Phytother. Res., 20: 1080-1084.
CrossRefPubMedDirect Link - Hamburger, M.O. and G.A. Cordell, 1987. A direct bioautographic TLC assay for compounds possessing antibacterial activity. J. Nat. Prod., 50: 19-22.
CrossRef - Hostettmann, K., J.L. Wolfender and C. Terreaux, 2001. Modern screening techniques for plant extracts. Pharmaceut. Biol., 39: 18-32.
Direct Link - Ito, S.I., T. Ihara, H. Tamura, S. Tanaka and T. Ikeda et al., 2007. α-Tomatine, the major saponin in tomato, induces programmed cell death mediated by reactive oxygen species in the fungal pathogen Fusarium oxysporum. FEBS Lett., 581: 3217-3222.
CrossRefDirect Link - Jorgensen, J.H., J.D. Turnidge and J.A. Washington, 1999. Antibacterial Susceptibility Tests: Dilution and disk Diffusion Methods. In: Manual of Clinical Microbiology, Murray, P.R., M.A. Pfaller, F.C. Tenover, E.J. Baron and R.H. Yolken (Eds.). 7th Edn., ASM Press, Washington, DC., USA., ISBN-13: 9781555811266, pp: 1526-1543.
- Lee, H.S., 2005. Pesticidal constituents derived from Piperaceae fruits. Agric. Chem. Biotechnol., 48: 65-74.
Direct Link - Lee, J.Y., J.Y. Lee, B.S. Yun and B.K. Hwang, 2004. Antifungal activity of β-asarone from rhizomes of Acorus gramineus. J. Agric. Food Chem., 52: 776-780.
CrossRefPubMedDirect Link - Mazza, G., 1985. Gas-chromatographic and mass spectrometric studies of the constituents of the rhizome of Calamus: I. The volatile constituents of the essential oil. J. Chromatogr. A, 328: 179-194.
CrossRefDirect Link - McGaw, L.J., A.K. Jager and J. Van Staden, 2002. Isolation of β-asarone, an antibacterial and anthelmintic compound, from Acorus calamus in South Africa. Afr. J. Bot., 68: 31-35.
Direct Link - Mehrotra, S., K.P. Mishra, R. Maurya, R.C. Srimal, V.S. Yadav, R. Pandey and V.K. Singh, 2003. Anticellular and immunosuppressive properties of ethanolic extract of Acorus calamus rhizome. Int. Immunopharmacol., 3: 53-61.
CrossRefDirect Link - Mungkornasawakul, P., D. Supyen, C. Jatisatienr and A. Jatisatienr, 2002. Inhibitory effect of Acorus calamus L. extract on some plant pathogenic molds. Acta Horticulturae, 576: 341-345.
Direct Link - Nawamaki, K. and M. Kuroyanagi, 1996. Sesquiterpenoids from Acorus calamus as germination inhibitors. Phytochemistry, 43: 1175-1182.
CrossRefDirect Link - Navarro, V., G. Rojas, G. Delgado and X. Lozoya, 1997. Antimicrobial compounds detected in Bocconia arborea extracts by a direct bioautographic method. Arch. Med. Res., 29: 191-194.
Direct Link - Perrett, S. and P.J. Whitfield, 1995. Anthelmintic and pesticidal activity of Acorus gramineus (Araceae) is associated with phenylpropanoid asarones. Phytother. Res., 9: 405-409.
Direct Link - Phongpaichit, S., N. Pujenjob, V. Rukachaisirikul and M. Ongsakul, 2005. Antimicrobial activities of the crude methanol extract of Acorus calamus Linn. Songklanakarin J. Sci. Technol., 27: 517-523.
Direct Link - Rahalison, L., M. Hamburger, K. Hostettmann, M. Monod and E. Frenk, 1991. A bioautographic agar overlay method for the detection of antifungal compounds from higher plants. Phytochem. Anal., 2: 199-203.
CrossRefDirect Link - Rani, A.S., M. Satyakala, V.S. Devi and U.S. Murty, 2003. Evaluation of antibacterial activity of rhizome extract of Acorus calamus Linn. J. Scient. Induast. Res., 2: 623-625.
Direct Link - Rost, L.C.M. and R. Bos, 1999. Biosystematic investigations with Acorus L. - 3. Communication1. Constituents of essential oils. Plant Med., 27: 350-361.
CrossRef - Sasidharan, S., Y. Chen, D. Saravaran, K.M. Sundram and L.Y. Latha, 2011. Extraction, isolation and characterization of bioactive compounds from plant's extracts. Afr. J. Tradit. Complementary Altern. Med., 8: 1-10.
CrossRefPubMedDirect Link - Suvarna, S., Vinod, H. Rajendra, K. Anil and G.C. Reddy, 2011. A unique water soluble formulation of β-asarone from sweet flag (Acorus calamus L.) and its in vitro activity against some fungal plant pathogens. J. Med. Plants Res., 5: 5132-5137.
Direct Link - Venskutonis, P.R. and A. Dagilyte, 2003. Composition of essential oil of sweet flag (Acorus calamus L.) leaves at different growing phases. J. Essent. Oil Res., 15: 313-318.
CrossRefDirect Link - Vincent, J.M., 1947. Distortion of fungal hyphae in the presence of certain inhibitors. Nature, 159: 850-850.
CrossRefPubMedDirect Link