
Boumaaza Boualem
Laboratory of Plant Protection, Department of Agronomy, University of Abdelhamid Ibn Badis, BP 300, 27000, Mostaganem, Algeria
Benkhelifa Mohamed
Laboratory of Plant Protection, Department of Agronomy, University of Abdelhamid Ibn Badis, BP 300, 27000, Mostaganem, Algeria
Belkhoudja Moulay
Laboratory of Vegetal Ecophysiology, Faculty of Sciences, University of Es Senia, BP 1524, ElMnouer, Oran, Algeria
Asian Journal of Plant Pathology
Year: 2015 | Volume: 9 | Issue: 4 | Page No.: 158-166







ABSTRACT
Three isolates of Botrytis cinerea were isolated from leaves and stems of different tomato varieties taken from four areas in the North-west of Algeria where tomato is mostly grown in greenhouses and high tunnels. The purpose of this study was to determine the effect of two salts: NaCl and CaCl2 tolerance of Trichoderma harzianum and to evaluate the impact of salinity on its antagonistic capacities in order to use it as a biological agent controlling Botrytis cinerea, the causal agent of grey mold disease of tomato. In the absence of salt, the volatile and non-volatile secondary metabolites from T. harzianum showed 52, 23-79, 81% inhibition of B. cinerea. In the presence of salt, the inhibition percentages of the mycelial growth of B. cinerea by volatile metabolites were stimulated by the high concentrations of NaCl 94,70 and 90,85% for CaCl2 compared to the control. However, non-volatile compounds from Trichoderma, the percentage of growth inhibition varied between 65.17% and 82, 12% for NaCl and 61.19-85.01 in the presence of CaCl2.
PDF Abstract XML References Citation
Received: September 14, 2015;
Accepted: October 20, 2015;
Published: November 12, 2015
How to cite this article
Boumaaza Boualem, Benkhelifa Mohamed and Belkhoudja Moulay, 2015. Salinity Influence upon Activity of Trichoderma harzianum against Botrytis cinerea. Asian Journal of Plant Pathology, 9: 158-166.
URL: https://scialert.net/abstract/?doi=ajppaj.2015.158.166
URL: https://scialert.net/abstract/?doi=ajppaj.2015.158.166
INTRODUCTION
Grey mould, caused by Botrytis cinerea (Sclerotiniaceae family) is an important plant disease that affects a large number of plant species and is particularly important in greenhouse production of tomatoes in Mediterranean basin (O’Neill et al., 1997). In greenhouse tomato, the fungus infects flowers, fruits and leaves and can grow through the petiole into the stem (Shtienberg et al., 1998; Kalogiannis et al., 2006).
Soil salinity is one of the major environmental factors that lead to a deterioration of agricultural land and reduction in crop productivity worldwide Keshavarzi et al. (2011). This problem is one of the major stresses especially in arid and semi-arid regions (Munns, 2002) and can severely limit plant growth and productivity (Allakhverdiev et al., 2000; Koca et al., 2007).
In Algeria, a wide range of environmental stresses (such as high and low temperature, drought, alkalinity, salinity and pathogen infection) are potentially harmful to the plants. Soil salinity and irrigation water are two of the main serious problems hindering the development of most plant species (Levignron et al., 1995). Thus, the effect of these factors may result from structural and physiological changes in the plant, an increase incidence and severity of diseases caused by various species pathogen. (Bouchibi et al., 1990) showed that relatively low levels of salinity (25-50 mEq) could increase the severity of phytophthora root rot of tomato with high Na:Ca ratios (10:1), Phytophthora (Blaker and MacDonald, 1986; MacDonald, 1982; Sanogo, 2004; Swiecki and MacDonald, 1988), F. oxysporum f. sp. vasinfectum (Turco et al., 2002), F. oxysporum f. sp. radicis-lycopersici (Jones et al., 1993), Verticillium dahliae and Alternaria solani (Nachmias et al., 1993).
Chemical control has always been essential for the management of gray mold, caused by Botrytis cinerea. Continuous use of the same fungicide against the same pathogen results in the development of fungicide resistant strains of the pathogen (Kumar and Dubey, 2001; Mamgain et al., 2013). Additionally chemical fungicides not only develop fungicide resistant strains but also accumulate in food and ground water as residues. In addition, it results in environmental hazards and has harmful side effects on human beings and animals. However, due to the polluting and non-biodegradable nature of such pesticides.
Biological control offers the chance to improve crop production within the existing resources, besides avoiding the problem of pesticide resistance (Khan et al., 2014). However, has been considered as a more natural and environmentally acceptable alternative to existing chemical treatment methods (Baker and Paulitz, 1996; Eziashi et al., 2007).
There are a variety of fungal species and isolates that have been reported as biocontrol agents, although Trichoderma species clearly dominate, perhaps due to their ease of growth and wide host range (Whipps and Lumsden, 2001). Trichoderma as a potent fungal biocontrol agent against a wide range of plant pathogens has attracted considerable scientific attention (Hermosa et al., 2012; Joshi et al., 2010; Moran-Diez et al., 2012). The effectiveness of this antagonist species depends on the suppress plant diseases by direct antibiosis or mycoparasitism as well as indirect IR (Lorito et al., 2010; Shoresh et al., 2010). The various mechanisms include antibiosis, parasitism, inducing host-plant resistance, competition and secretion of chitinolytic enzymes, mycoparasitism and production of inhibitory compounds (Harman et al., 2004). For the most effective control of disease, it seems necessary to examine the impact of salinity on its antagonistic capacities, in order to use it as a biological agent controlling B. cinerea of tomato.
MATERIALS AND METHODS
Fungal isolates: Botrytis cinerea isolates were obtained from decayed tomato (Lycopersicon esculentum) in North-western Algeria. The leaf fragments were placed on filter paper moistened with sterile water in a Petri dish. B. cinerea was cultured on Potato Dextrose Agar (PDA) incubated at 25°C.
Antagonist: The isolate of Trichoderma harzianum, obtained from the laboratory of plant protection (University of Mostaganem, Algeria) was grown in PDA. The media were added with the same concentration of NaCl (50, 100, 150, 300 mEq).
Colony growth inhibition assay with Trichoderma harzianum in dual culture method: Interactions between antagonistic of T. harzianum against Botrytis cinerea were determined by the method described by Dennis and Webster (1971). A 4 mm diameter mycelial disc from the margin of the Trichoderma one week old culture and the pathogen Botrytis cinerea were placed on opposite of the plate at equal distance from the periphery and in control plates only Botrytis cinerea was placed. The plates were incubated at 25±1°C, observed after 4 days and the colony interactions were measured as percentage of inhibition of radial growth of Botrytis cinerea by following equation:
where, L is the percentage inhibition of radial mycelial growth, D1 is radial growth of the pathogen in the control, D2 is radial growth of the pathogen in the presence of T. viride (Edgington et al., 1971).
Evaluation of volatile metabolites: The effect of the volatile metabolites released by T. harzianum on the mycelial growth of the pathogens was evaluated by the inverted plate technique as described by Dennis and Webster (1971). The 5 mm mycelial discs of T. harzianum obtained from the margin of young cultures were placed centrally on the PDA glass dish and incubated in 25±1°C for 72 h. In the control plates, sterile PDA media discs 5 mm in diameter were placed on the plates as mentioned above. At the end of the incubation period, the top of each petri dish was replaced with the bottom of the petri dish inoculated with pathogen and sealed together with adhesive tape. A completely randomized experimental design was used with three replicates. Radial growth of the pathogens was recorded on the 5th day of incubation and L was calculated, as described above.
The influence of NaCl and CaCl2 on the diameter growth was determined by growing the isolates in a PDA medium at 4 NaCl and CaCl2 levels (50, 100, 150, 300 mEq), control medium was not amended with salts.
Statistical analysis: All statistical analyses were analyzed by the software of statistics (STATBOX 6.0.4 grimmersoft). The data was analyzed by two-way factorial. Comparison of means and interactions was performed by Duncan’s multiple range tests. Statistical significance was assessed at the level of p = 0.05 or p = 0.01.
RESULTS
Effect of non-volatile metabolites from T. harzianum on the radial growth of B. cinerea: In the absence of salt (Table 1), isolates of B. cinerea do not present the same profile of growth inhibition, the optimum percent for growth inhibition of this fungus was from 63.76-68.15 of F27 and TR46, respectively. In the dual culture experiment, T. harzianum had a marked significant inhibitory effect on the growth of isolates (Table 1). Growth inhibition decreased as follows: TR46 (68.15%)>B27 (66.8 %)>F27 (63.76%). Data in Table 1 indicate that, application of sodium salt caused a significant increase the growth inhibition at various concentrations tested compared with control except TR46 (p>0.05), growth inhibition was decreased for sodium concentrations of 50 and 100 ppm can be obtained from this isolate relative to the control.
Calcium chloride stimulates mycelial growth of T. harzianum. These observations indicate that the growth inhibition of the F27 and B27 isolates might increase (85.01, 70.17%), even at high salinity (150 ppm). However, higher concentrations of calcium chloride (300 ppm) caused F27 and B27 to reduce the percentage of inhibition growth by 81.63 and 65.84%, respectively. Calcium chloride was reducing growth inhibition of the TR46 in the dual culture for all concentrations. The interaction between salt and concentration was significant (p<0.001).
Effect of volatile metabolites from T. harzianum on the radial growth of B. cinere: In the absence of salt, TR46 and F27 was most resistant and revealed lowest percent inhibition of mycelial growth as 52,23% and 53,13, respectively in combination with T. harzianum.
| Table 1: | Growth inhibition of three isolates by Trichoderma harzianum in dual culture after incubation at 25°C for 3 days |
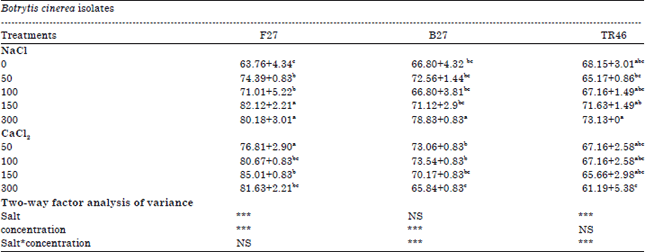 | |
| *,**,***Significant effects at 0.05, 0.01 and 0.001, respectively, NS: Not significant | |
| Table 2: | In vitro growth inhibition of three isolates of Botrytis cinerea by volatile compounds of T. harzianum in PDA medium after incubation at 25oC for 3 days |
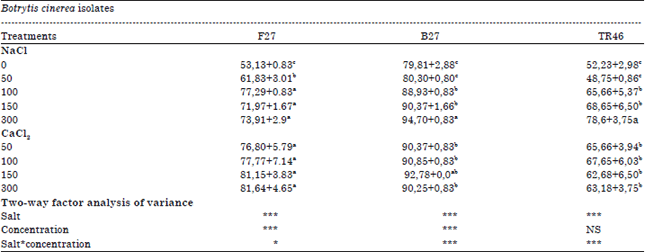 | |
| ****,**,***Significant effects at 0.05, 0.01, 0.001, respectively, NS: Not significant | |
After 3 days of incubation, it is observed that volatile compounds from Trichoderma harzianum exhibited maximum growth inhibition by adding 300 ppm of NaCl to the culture medium when compare to the others salts (Fig. 1). The highest growth inhibition, 94,70, 78,6 and 73,91% was obtained in the high concentrations from isolates B27, TR46 and F27, respectively. The interaction between salt and concentration was significant for all salts isolates (p<0.05) (Table 2). In the presence of low salt concentrations (50 and 100 ppm), the percentage of inhibition growth due to the interaction between fungi and T. harzianum was still very important with CaCl2 more than NaCl. However, the antifungal activity of volatile metabolites produced by T. harzianum showed 90.037% maximum inhibition of mycelia growth with B27 in presence of higher concentrations of calcium chloride (300 ppm). On the other hand the effect of CaCl2 on T. harzianum volatile metabolites action on the growth of B27 isolat presented in Table 2 showed that the volatile metabolic substances of antagonistic present the same profile of the percentage inhibition of mycelia growth in the different concentrations tested.
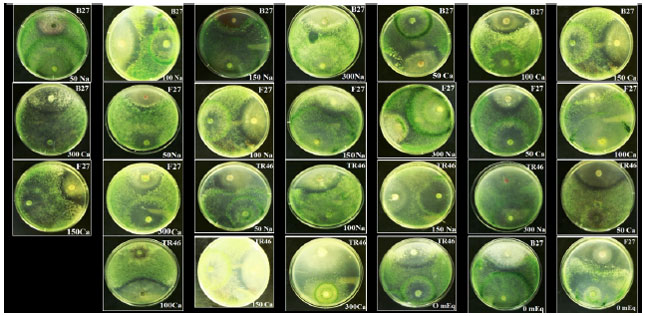 | |
| Fig. 1: | Antagonistic effect of the isolates on Botrytis cinerea in dual culture assay at 25°C on potato dextrose agar media |
DISCUSSION
In the present study, we tested effect of salinity on in vitro Trichoderma harzianum antagonism against B. cinerea for their production of volatile and non-volatile compounds.
In the absence of salt, The direct confrontation of T. harzianum against the different isolates of B. cinerea in vitro on PDA medium showed that T. harzianum inhibited the growth of pathogenic fungi at varying degrees.
The non-volatile secondary metabolites from T. harzianum were found more effective in suppressing the mycelial growth of all isolate when compared to volatile compounds except B27 isolate, growth inhibition was recorded by volatile compounds up to 79,81%.
The inhibition in radial growth of two interacting organisms in dual culture has been attributed to secretion of extracellular hydrolytic enzymes (Schirmbock et al., 1994) by the production of antibiotics Howell (1998) or as well as some cell walls degrading enzymes such as chitinases, glucanases that break down polysaccharides, chitins and þ-glucanase, there- by destroying cell wall integrity (Elad, 2000). Trichoderma species are known to produce a number of antibiotics such as Trichodernin, Trichodermol, Harzianum A and Harzianolide (Dennis and Webster, 1971; Kucuk and Kivanc, 2004).
Ajith and Lakshmidevi (2010) examined the potential of Trichoderma harzianum which suppress the mycelia growth of Colletotrichum capsici causing anthracnose on bell peppers. Doi and Mori (1994) reported volatile compounds produced from Trichoderma species were able to arrest and inhibit the hyphal growth of various plant pathogenic fungi. Amin et al. (2010) reported the effectiveness of volatiles produced by T. harzianum (Th-1) causing 53.63% inhibition of Colletotrichum capsici.
In the case of the evaluation the effect of NaCl and CaCl2 on the Trichoderma harzianum against B. cinerea, salinity is most favorable to the increase growth of Trichoderma harzianum, especially at high concentrations. T. harzianum was able to tolerate the presence of salt in the medium culture even at high concentrations.
The analysis shows that CaCl2 stimulate the development of the Trichoderma harzianum as compared to sodium chloride.
Our data indicate that volatile compounds from Trichoderma were found more effective in suppressing the mycelial growth of all fungal pathogens when compared to non-volatile metabolites.
In indirect confrontation, the effect of salinity on T. harzianum volatile compounds action is perceived in salt concentrations demonstrate that T. harzianum was able to colonize the space and to inhibit the mycelial growth of Botrytis cinerea. It was found that NaCl inhibited the growth of pathogen (94,70%) more than CaCl2 (92,78%).
In the direct confrontation, the growth of Botrytis cinerea was intensely inhibited by non-volatile metabolites was stimulated by salt and moderately inhibited the pathogen growth. The percentage of growth inhibition varied between 65.17 and 82, 12% for NaCl and 61.19-85.01 in the presence of CaCl2.
The growth and multiplication of T. harzianum were stimulated by the addition of salt to the growth media, it showed also some good antagonist abilities at the high concentrations of salinity demonstrated by confrontation techniques. The strain activity of T. harzianum was able to tolerate the presence of salt in the medium culture even at high concentrations. Benyahyia (1998) showed that increasing the salinity of the medium promotes the in vitro mycelia growth of Phytophthora citrophthora and P. parasitica agents of root rot of citrus with an optimum between -1.44 and -3.11 bars. The mycelium growth of T. harzianum developed in saline media between 2 and 6 g L‾1 of NaCl doses not differ from the control cultures without salt. Nevertheless, the capacity C declines significantly starting from 6 g L‾1 of NaCl but C value remain high (80%), showing an important capacity of the antagonist to cover the colony of the pathogen in presence of salt (Regragui and Lahlou, 2005).
Growth inhibition of the pathogens by the Trichoderma metabolites has been reported by several workers., such as Fusarium oxysporum, Rshizoctonia solani, Curvularia lunata, Bipolaris sorokiniana and Colletotrichum lagenarium, Colletotrichum acutatum, Colletotrichum gloeosporioides (Yang et al., 2006; Zivkovic et al., 2010). The antagonistic activity of Trichoderma depends on multiple synergistic mechanisms (Nallathambi et al., 2009; Howell, 2003). The various mechanisms include antibiosis, parasitism, inducing host-plant resistance, competition and secretion of chitinolytic enzymes, mycoparasitism and production of inhibitory compounds (Harman et al., 2004). Also it is found that there is large variety of volatile secondary metabolites produced by Trichoderma such as ethylene, hydrogen cyanide, aldehydes and ketones which play an important role in controlling the plant pathogens (Vey et al., 2001). Previous studies have demonstrated that before mycelia of fungi interact, Trichoderma sp. produces low quantities of extracellular exochitinases (Kullnig et al., 2000; Brunner et al., 2003). The diffusion of these enzymes dissolves cell fragments of host cells. These cell fragments in turn induce the production of further enzymes and trigger a cascade of physiological changes, stimulating rapid and directed growth of Trichoderma sp. (Zeilinger et al., 1999).
Although, Trichoderma sp. species is widely used to control many plant pathogens but not much work has been done to give the effect of salinity on the antagonist effect of Trichoderma sp. Knowledge of soil salinity and its potential effects on the antagonist against diseases is essential for disease management strategies. Therefore, it is important that biocontrol potential of Trichoderma harzianum under salinity conditions should be further evaluated in vivo against Botrytis cinerea causing grey mould disease.
REFERENCES
- Ajith, P.S. and N. Lakshmidevi, 2010. Effect of volatile and non-volatile compounds from Trichoderma spp. against Colletotrichum capsici incitant of anthracnose on bell peppers. Nat. Sci., 8: 265-269.
Direct Link - Allakhverdiev, S.I., A. Sakamoto, Y. Nishiyama, M. Inaba and N. Murata, 2000. Ionic and osmotic effects of NaCl-induced inactivation of photosystems I and II in Synechococcus sp. Plant Physiol., 123: 1047-1056.
Direct Link - Joshi, B.B., R.P. Bhatt and D. Bahukhandi, 2010. Antagonistic and plant growth activity of Trichoderma isolates of Western Himalayas. J. Environ. Biol., 31: 921-928.
Direct Link - Blaker, N.S. and J.D. MacDonald, 1986. The role of salinity in the development of phytophthora root rot of citrus. Phytopathology, 76: 970-975.
CrossRefDirect Link - Bouchibi, N., A.H.C. van Bruggen and J.D. MacDonald, 1990. Effect of ion concentration and sodium: Calcium ratio of a nutrient solution on Phytophthora root rot of tomato and zoospore motility and viability of Phytophthora parasitica. Phytopathology, 80: 1323-1329.
CrossRefDirect Link - Brunner, K., C.K. Peterbauer, R.L. Mach, M. Lorito, S. Zeilinger and C.P. Kubicek, 2003. The Nag1 N-acetylglucosaminidase of Trichoderma atroviride is essential for chitinase induction by chitin and of major relevance to biocontrol. Curr. Genet., 43: 289-295.
CrossRefDirect Link - MacDonald, J.D., 1982. Effect of salinity stress on the development of Phytophthora root rot of Chrysanthemum. Phyopathology, 72: 214-219.
CrossRefDirect Link - Dennis, C. and J. Webster, 1971. Antagonistic properties of species-groups of Trichoderma: II. Production of volatile antibiotics. Trans. Br. Mycol. Soc., 57: 41-48.
CrossRefDirect Link - Elad, Y., 2000. Biological control of foliar pathogens by means of Trichoderma harzianum and potential modes of action. Crop Protect., 19: 709-714.
CrossRefDirect Link - Eziashi, E.I., I.B. Omamor and E.E. Odigie, 2007. Antagonism of Trichoderma viride and effects of extracted water soluble compounds from Trichoderma species and benlate solution on Ceratocystis paradoxa. Afr. J. Biotechnol., 6: 388-392.
Direct Link - Amin, F., V.K. Razdan, F.A. Mohiddin, K.A. Bhat and P.A. Sheikh, 2010. Effect of volatile metabolites of Trichoderma species against seven fungal plant pathogens in-vitro. J. Phytol., 2: 34-37.
Direct Link - Harman, G.E., C.R. Howell, A. Viterbo, I. Chet and M. Lorito, 2004. Trichoderma species-opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol., 2: 43-56.
CrossRefPubMedDirect Link - Howell, C.R., 2003. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The history and evolution of current concepts. Plant Dis., 87: 4-10.
CrossRefDirect Link - Kalogiannis, S., S.E. Tjamos, A. Stergiou, P.P. Antoniou, B.N. Ziogas and E.C. Tjamos, 2006. Selection and evaluation of phyllosphere yeasts as biocontrol agents against grey mould of tomato. Eur. J. Plant Pathol., 116: 69-76.
CrossRefDirect Link - Khan, F., M. Mazid, T.A. Khan, H.K. Patel and R. Roychowdhury, 2014. Plant derived pesticides in control of lepidopteran insects: Dictum and directions. Res. J. Biol., 2: 1-10.
Direct Link - Koca, H., M. Bor, F. Ozdemir and I. Turkan, 2007. The effect of salt stress on lipid peroxidation, antioxidative enzymes and proline content of sesame cultivars. Environ. Exp. Bot., 60: 344-351.
CrossRefDirect Link - Kucuk, C. and M. Kivanc, 2004. In vitro antifungal activity of strains of Trichoderma harzianum. Turk. J. Biol., 28: 111-115.
Direct Link - Kullnig, C., R.L. Mach, M. Lorito and C.P. Kubicek, 2000. Enzyme diffusion from Trichoderma atroviride (=T. harzianum P1) to Rhizoctonia solani is a prerequisite for triggering of Trichoderma ech42 gene expression before mycoparasitic contact. Applied Environ. Microbiol., 66: 2232-2234.
CrossRefDirect Link - Kumar, D. and S.C. Dubey, 2001. Management of collar rot of pea by the integration of biological and chemical methods. Indian Phytopathol., 54: 62-66.
Direct Link - Levignron, A., F. Lopez, G. Vansuyt, P. Berthomieu, P. Foucroy and F. Casse-Delbart, 1995. Les plantes face au stress salin. Cahiers Agric., 4: 263-273.
Direct Link - Lorito, M., S.L. Woo, G.E. Harman and E. Monte, 2010. Translational research on Trichoderma: From omics to the field. Annu. Rev. Phytopathol., 48: 395-417.
CrossRefPubMedDirect Link - Mamgain, A., R. Roychowdhury and J. Tah, 2013. Alternaria pathogenicity and its strategic controls. Res. J. Biol., 1: 1-9.
Direct Link - Keshavarzi, M.H.B., M.S.O. Rafsanjani S.M. Moussavinik and A.P Lak, 2011. Effect of salt (NaCl) stress on germination and early seedling growth of Spinach (Spinacia oleracea L.). Ann. Biol. Res., 2: 490-497.
Direct Link - Moran-Diez, E., B. Rubio, S. Dominguez, R. Hermosa, E. Monte and C. Nicolas, 2012. Transcriptomic response of Arabidopsis thaliana after 24 h incubation with the biocontrol fungus Trichoderma harzianum. J. Plant Physiol., 169: 614-620.
CrossRefDirect Link - Munns, R., 2002. Comparative physiology of salt and water stress. Plant Cell Environ., 25: 239-250.
CrossRefDirect Link - Nachmias, A., Z. Kaufman, L. Livescu, L. Tsror, A. Meiri and P.D.S. Caligari, 1993. Effects of salinity and its interactions with disease incidence on potatoes grown in hot climates. Phytoparasitica, 21: 245-255.
CrossRefDirect Link - Nallathambi, P., C. Umamaheswari, B.B.L. Thakore and T.A. More, 2009. Post-harvest management of ber (Ziziphus mauritiana Lamk) fruit rot (Alternaria alternata Fr. Keissler) using Trichoderma species, fungicides and their combinations. Crop Protect., 28: 225-232.
CrossRefDirect Link - O'Neill, T.M.O., D. Shtienberg and Y. Elad, 1997. Effect of some host and microclimate factors on infection of tomato stems by Botrytis cinerea. Plant Dis., 81: 36-40.
CrossRefDirect Link - Regragui, A. and H. Lahlou, 2005. Effect of salinity on in vitro Trichoderma harzianum antagonism against Verticillium dahliae. Pak. J. Biol. Sci., 8: 872-876.
CrossRefDirect Link - Hermosa, R., A. Viterbo, I. Chet and E. Monte, 2012. Plant-beneficial effects of Trichoderma and of its genes. Microbiology, 58: 17-25.
CrossRefDirect Link - Sanogo, S., 2004. Response of chile pepper to Phytophthora capsici in relation to soil salinity. Plant Dis., 88: 205-209.
CrossRefDirect Link - Schirmbock, M., M. Lorito, Y.L. Wang, C.K. Hayes and I. Arisan-Atac et al., 1994. Parallel formation and synergism of hydrolytic enzymes and peptaibol antibiotics, molecular mechanisms involved in the antagonistic action of Trichoderma harzianum against phytopathogenic fungi. Applied Environ. Microbiol., 60: 4364-4370.
Direct Link - Shtienberg, D., Y. Elad, A. Niv, Y. Nitzani and B. Kirshner, 1998. Significance of leaf infection by Botrytis cinerea in stem rotting of tomatoes grown in non-heated greenhouses. Eur. J. Plant Pathol., 104: 753-763.
CrossRefDirect Link - Shoresh, M., G.E. Harman and F. Mastouri, 2010. Induced systemic resistance and plant responses to fungal biocontrol agents. Annu. Rev. Phytopathol., 48: 21-43.
CrossRefDirect Link - Živković, S., S. Stojanović, Ž. Ivanović, V. Gavrilović, T. Popović and J. Balaž 2010. Screening of antagonistic activity of microorganisms against Colletotrichum acutatum and Colletotrichum gloeosporioides. Arch. Biol. Sci., 62: 611-623.
CrossRefDirect Link - Swiecki, T.J. and J.D. MacDonald, 1988. Histology of chrysanthemum roots exposed to salinity stress and Phytophthora cryptogea. Can. J. Bot., 66: 280-288.
CrossRefDirect Link - Turco, E., D. Naldini and A. Ragazzi, 2002. Disease incidence and vessel anatomy in cotton plants infected with Fusarium oxysporum f. sp. vasinfectum under salinity stress. J. Plant Dis. Protect., 109: 15-24.
Direct Link - Yang, D.M., Y. Bi, X.R. Chen, Y.H. Ge and J. Zhao, 2006. Biological control of postharvest diseases with Bacillus subtilis (B1 strain) on muskmelons (Cucumis melo L. cv. Yindi). Acta Horticulturae, 712: 735-740.
CrossRefDirect Link - Zeilinger, S., C. Galhaup, K. Payer, S.L. Woo and R.L. Mach et al., 1999. Chitinase gene expression during mycoparasitic interaction of Trichoderma harzianum with its host. Fungal Genet. Biol., 26: 131-140.
CrossRefDirect Link - Edgington, L.V., K.L. Khew and G.L. Barron, 1971. Fungitoxic spectrum of benzimidazole compounds. Phytopathology, 61: 42-44.
CrossRefDirect Link