
Hoda M. Soliman
Department of Botany, Faculty of Science, Mansoura University, Egypt
Mohamed A. El-Metwally
Department of Mycological Research, Plant Pathology Research Institute, Agricultural Research Center, Egypt
Maged Taher Elkahky
Department of Plant Pathology, Faculty of Agriculture, Mansoura University, Egypt
Wael E. Badawi
Department of Botany, Faculty of Science, Mansoura University, Egypt
Asian Journal of Plant Pathology
Year: 2015 | Volume: 9 | Issue: 1 | Page No.: 1-15







ABSTRACT
Assessment the efficiency of fungal antagonists and chitosan as foliar application against cucumber grey mold was carried out in greenhouse condition. The evaluated antagonistic fungi were Trichoderma harzianum, Trichoderma hamatum, Trichoderma viride, Gliocladium roseum and Gliocladium vireins. Chitosan was tested at three concentrations 0.1, 0.2 and 0.3%. It was noticed that all treatments significantly reduced grey mold incidence in cucumber plants comparing with untreated plants. Among all tested antagonists, T. harzianum and T. viride better than others were in inhibiting disease incidence and improving plant defense against the pathogen. Chitosan treatments did not show antimicrobial activity against Botrytis cinerea isolates in vitro however, when it used as a foliar spray on greenhouse it showed a highly significant control level of grey mold disease. According to obtained results from the current study, it could be suggested that the application of antagonistic fungi and chitosan might be an easily applied, safely and cost effective alternative control method to chemical control of grey mold of cucumber.
PDF Abstract XML References Citation
Received: December 16, 2014;
Accepted: February 21, 2015;
Published: May 04, 2019
How to cite this article
Hoda M. Soliman, Mohamed A. El-Metwally, Maged Taher Elkahky and Wael E. Badawi, 2015. Alternatives to Chemical Control of Grey Mold Disease on Cucumber Caused
by Botrytis cinerea Pers. Asian Journal of Plant Pathology, 9: 1-15.
URL: https://scialert.net/abstract/?doi=ajppaj.2015.1.15
URL: https://scialert.net/abstract/?doi=ajppaj.2015.1.15
INTRODUCTION
Cucumber (Cucumis sativus L.) is an important vegetable and one of the most popular members of the Cucurbitaceae family (Lower and Edwards, 1986). Cucumber is a creeping vine that roots in the ground and grows up using supporting frames, wrapping around ribbing with thin, spiraling tendrils. Cucumber also, considers basic part of almost any human diets. It usually consumed as fresh product on salads. Controlling cucumber diseases with chemical pesticides put the human health in a direct risk since it is directly consumed without any process like cocking or any other preparing processes. Most cucumber production especially in Egypt coming through greenhouses system where intensive growth and all plant surrounding conditions consider favorable for almost all fungal diseases incidence particularly grey mold (sometimes known as botrytis blight). Grey mold (botrytis blight) caused by Botrytis cinerea Pers. which is a very common fungus in greenhouses. It has a global distribution and wide host range (Jarvis, 1977). Grey mold symptoms could be noticed on all parts of the infected plant since the pathogen could attack all old and fresh plant parts. Symptoms usually start with water soaked spot in any part followed by collapsing of this part or completely infected plant. Botrytis cinerea could cause seedlings damping off, stem or blossom blight or fruit rot but the characteristic symptom of botrytis infection of cucumber fruits in a greenhouse likely show as aggressive grey rot of the fruit (Ten Have et al., 2002). In the springtime, the fungus sclerotia which usually looked small, hard over-seasoning, dark-colored, structures start to germinate giving asexual spores (conidia) that helping disease to be spread in the area where the susceptible host growing. Fungal spores are continued to be produced during the growing season to re-infect new plants. Symptoms start to develop under warm temperature ranged from 18-25°C and dominant moist conditions. Copper or sulfur based organic fungicides generally used to help protecting plants from infections by apply these pesticides on a weekly base, when weather condition is continuously warm and wet or if Botrytis caused a problem in the past. Since the control of airborne pathogens depends mainly on the use of pesticides applications which increase the input of pesticides into the environment and cause development of pesticide-resistant strains of the pathogen, continues efforts are being made to look for a new safer, environmentally friend alternative protection strategies. Biological control agents are becoming increasingly interesting as alternatives to the use of chemical fungicides that are helping in decrease hazardous to the environment as well as being responsible in bringing about resistance to the disease (Elad et al., 1992). Plant resistance induction also is one of the new aspects that are developed during recent study to consider as one of possible alternative to chemical control. This study aims to evaluate safe and non-chemical alternative control method for grey mold of Cucumis sativus under greenhouse conditions. In general, application of biological control agents can destroy total or partial pathogen populations (Agrios, 1997) as Trichoderma spp. and Gliocladium spp. beside it could works as bio elicitors that enhance plant self-defense. Another solution might be the activation of the plant’s own defense system, known as Systemic Induced Resistance (ISR) (Sticher et al., 1997). This induced systemic resistance triggered biologically as reaction of application of some certain active oligoglucosides that called chemical elicitors (Benhamou et al., 1994) as chitosan. The aim of study, it could be suggested that the application of antagonistic fungi and chitosan might be an easily applied, safely and cost effective alternative control method to chemical control of grey mold of cucumber.
MATERIALS AND METHODS
Fungal pathogen isolation: Infected plant samples were collected from infected cucumber plants cultivated in different greenhouses in Kafr Badawi Village, Mansoura District in Dakahlia Governorate, Egypt. All collected samples were from plants showing the typical symptoms of grey mold. All samples were cut into small pieces then, washed in running tap water. After that, all samples were surface sterilized with 1% solution of sodium hypochlorite (NaOCl) for 1-3 min then rinsed directly with sterilized water for several times before drying them on sterilized filter paper. The dried pieces were then placed on the surface of pre poured plates of Potato dextrose agar media (PDA) supplemented with streptomycin sulphate (200 ppm). The dishes were incubated at 23±2°C for 5-7 days. Resulted isolates were purified using single spore technique according to Shabana (1987) then identified according to cultural characteristics; morphological and microscopic features (mycelial development and spore formation) described by Ellis (1971) and Sneh et al. (1992). All confirmed pathogenic Botrytis cinerea isolates were kept in PDA slants for further studies.
Pathogen spore suspension: Spore suspension of B. cinerea was mad by adding 5 mL of distilled water and two drops of Tween 80 to 7-10 days old B. cinerea cultures grown in PDA then scraping the surface of the culture with a sterilized spatula then filter the collected suspension through 2 layers of cheese cloth. The population of the suspension was adjusted to be 105 CFU mL-1 using Haemocytometer slide.
Biological Control Agents (BCAs): All antagonistic fungi used through the current study were obtained from Plant pathology Department, Faculty of Agriculture; Mansoura University. The certified isolates were Trichoderma harzianum, T. hamatum, T. viride, Gliocladium roseum and G. vireins. All used Isolates were cultured and maintained on Potato Dextrose Agar (PDA).
Pathogenicity test: It was carried out on 5 isolates of B. cinerea to choose the most aggressive isolate which would be used throughout this investigation. All B. cinerea, isolates were grown on PDA. Light regime of UV and normal fluorescent (12/12 h) was used to enhance sporulation (Last and Hamley, 1956). Spore suspension was prepared as mention above. The suspension used as a inoculum and it was mixed thoroughly with the surface layer of the pot soil were mixed with inoculum with rate 10 mL per pot then all inoculated pots were watered and left for one week to ensure even distribution of the inoculated fungus of B. cinerea. Three pots were used as replicates for each treatment as well as check treatment (uninfested soil). Hesham hybrid seeds (Seminis Company, 800 North Lindbergh Blvd-St. Louis, Missouri, USA) of Cucumis sativus which known as a susceptible to grey mold disease, were obtained from the El-Meer Company, Mansoura, Egypt. Three seeds were sown in each plastic pots containing 2-3 kg of soil mixture of clay: sand (3:1 v/v). Plants were Kept at greenhouse to grow and watered when necessary. To emphasis the infection, each pot was sprayed with 10 mL spore suspension (104 CFU mL-1 of B. cinerea/mL) 7 days after planting covered with polyethylene bags to maintain enough humidity around the plants. Control plants were sprayed with sterilized water. Disease incidence of pre-and post-emergence of damping off of cucumber plants were recorded after 14 days.
Laboratory experiments
Evaluation of the antagonistic fungi activity against Botrytis cinerea (dual culture test) in vitro: The antagonistic effect of above mentioned fungi was investigated using dual culture technique described by Raju and Murthy (2000). The interaction between the two fungi (pathogenic and antagonistic) was recorded either in the form of inhibition zone or overgrowth of fungi on B. cinerea (Desai et al., 2002). This experiment was conducted in 3 replicates per each fungus. Inhibition percentages of the pathogen were calculated just after overlapping of the two tested fungi according to the following equation:
where, A is the diameter of mycelial growth of pathogenic fungus in control and B is the diameter of mycelial growth of pathogenic fungus with antagonist.
Evaluation of antimicrobial activity of chitosan against Botrytis cinerea (dual culture test) in vitro: Chitosan was suspended in 0.1% acetic acid in distilled water according to Eikemo et al. (2003). The medium PDA was amended with chitosan just before solidification. The appropriate amount of chitosan was prepared by mixing it with 50 mL medium in each flask to give end concentrations of 0.1, 0.2 and 0.3% of chitosan. Each of 20 mm of PDA medium was poured in each Petri dish and then they were inoculated with 6 mm discs of fungal growth and incubated at 23±1°C. Three replicates were used per each treatment. Check Petri dishes having no chitosan were served as control treatment.
Greenhouse experiments
Foliar spray by biocontrol agents: Above-mentioned antagonistic fungi were tested as a foliar spray to cucumber plants in greenhouse. Suspension of the antagonists were prepared by inoculate conical flasks (500 mL) containing 150 mL sterilized potato dextrose broth with two of 6 mm diameter discs taken from the growing edge of 5 day old culture of each fungus and were incubated at 25±2°C for 7 days. After incubation, the fungal mats were collected in sterilized distilled water (10 g/100 mL water). The fungal suspension (spore and hyphae) was mixed and homogenized using sterile water and 0.03% Tween-20. The suspension population was determined using dilution plate method. After four weeks from planting under natural infection conditions of B. cinerea, the Trichoderma spp. and Gliocladium spp. of (4-5x107 CFU mL-1) and chitosan at concentrations 0.1, 0.2 and 0.3% (w/v) (15% degree of acetylation [DA], MW 1,000,000 kDa) were applied as foliar spray for 6 times on a weekly base. A complete randomized block design with three replicates was used. Each replicate contained ten plants. Samples of leaves and fruits were collected after 6, 8 and 10 weeks after planting for different analysis. Untreated plants (control) were sprayed with sterilized water at the same time.
Measurements
Disease incidence: As an indicator for the efficacy of the tested treatment on disease incidence was calculated for all tested treatments by obtaining the percentage of infected plants with grey mold in each treatment at the end of the experiment.
Disease severity: Percentage of each fruits rot disease severity was recorded as following equation:
whereas, D.S is disease severity %, n is number of infected fruits per scale, c is scale number and N is total examined fruits. Disease severity scale from 0 to 4 according to Cohen et al. (1991) was used, whereas, 0 = No fruits lesions, 1 = 1-25%, 2 = 26-50%, 3 = 51-75% and 4 = 76-100% infected area of cucumber fruits.
Determination of photosynthetic pigments: Third upper leaf was used to determine photosynthetic pigments after 50 days from planting according to the methods described by Mackinney (1941). Adding 10 mL methanol 90% to 0.05 g of fresh material for 24 h to extract the pigments then measure the absorbance of each sample by spectrophotometer at the wavelength 452.5, 650 and 665 nm for chlorophyll a, b and carotenoids, respectively.
Determination of total phenolic compounds: After 70 days from planting in fresh shoots were used to determine total phenol was using the Folin-Ciocalteu reagent (Singleton and Rossi Jr., 1965). Samples (2 g) were homogenized in 80% ethanol at room temperature. After that, samples were centrifuged at 10000 rpm for 15 min, under cooling and the supernatants were saved. The obtained residues were re-extracted twice in 80% ethanol and supernatants were collected, placed into evaporating dishes and evaporated to dryness at room temperature. Residues were dissolved in 5 mL of distilled water. One-hundred microlitres of this extract were diluted to 3 mL with distilled water and 0.5 mL of Folin-Ciocalteau reagent was added. After 3 min, 2 mL of 20% of sodium carbonate was added and the contents were mixed thoroughly. The developed color was photometrically measured at 650 nm wavelength after 60 min using catechol as a standard. The results were expressed as mg catechol/100 g of fresh weight material.
Assay of defense related enzymes activity
Enzyme extraction: Cucumber leaves were collected 72 h after each treatment and directly immersed in liquid nitrogen, lyophilized and then milled to a fine powder with a mortar and pestle. The powder (0.1 g) was extracted using 5 mL sodium phosphate buffer (0.05 M, pH 6.5) for 2 h at 4°C and then centrifuged at 20,000 g for 15 min at 4°C. After centrifugation, the supernatant was saved. The supernatant was designated as the crude leaf extract and its protein content was determined in comparison with a BSA (Bovine Serum Albumin) standard (Bradford, 1976).
Peroxidase activity (POD) activity: A 1 mL sample of assay mixture containing 0.05 M sodium acetate buffer pH 6.0, 0.2 mL of 0.1 M pyrogallol, 0.1 mL of 90 mM H2O2 and an aliquot of the crude leaf extract containing 10-40 μg protein were used. The reaction was allowed to precede for 6 min with Optical Density (OD) measurements taken every 60 sec, beginning 1 min after the crude leaf extract was added to the substrate (Lurie et al., 1997). Peroxidase enzyme activity was expressed as ΔOD300 nm min-1 mg-1 of protein.
Polyphenoloxidase activity (PPO) activity: Polyphenol oxidase activity was assayed as the increase in absorbance at 420 nm due to the formation of purpurogallin (Devi, 2002). The reaction mixture contained 2 mL of 0.02 M phosphate buffer (pH 7), 1 mL of 0.1 M pyrogallol and 1 mL of enzyme extract. The reaction mixture was incubated for 1 min at 25°C and the reaction was stopped by adding 1 mL of 2.5 N H2SO4. One enzyme unit is defined as unit per g fresh mass per minute.
Phenylalanine Ammonia Lyase activity (PAL): Phenylalanine ammonia lyase activity was determined according to the method described by Brueske (1980). Adding 0.5 mL borate buffer, 0.2 mL enzyme extract and 1.3 mL distilled water to the sample in a test tube, then the reaction was initiated by adding 1 mL (0.1 M) L-phenylalanine solution. After that, the reaction mixture incubated for 30-60 min at 32°C. Reaction was stopped by adding 0.5 mL of (1 M) trichloroacetic acid. A control treatment was done by adding phenylalanine after trichloroacetic acid. The absorbance was measured at 290 nm using spectrophotometer. Enzyme activity was expressed as unit per gram of fresh mass per min.
Statistical analysis: Obtained data were statistically analyzed through CoStat 3.4 software as the usual technique of analysis of variance (Gomez and Gomez, 1984). The means were compared using Least Significant Difference (LSD) at p = 0.05 as outlined by Duncan (1955).
RESULTS
Pathogenicity test: All 5 obtained isolates of B. cinerea were subjected to pathogenicity tests to determine the most aggressive one. The results indicated that all isolates have the potency to cause grey mold disease as causing typical symptoms of pre and post-emergence damping-off on cucumber seedlings.
| Table 1: | Percentage of pre and post-emergence damping-off on cucumber seedlings under greenhouse condition (artificial infection) |
 | |
| *Values followed by the same letter(s) in each column do not differ significantly (p≤0.05). Each value represents the mean of 3 replicates | |
| Table 2: | Effect of different treatments on percent inhibition of radial colony growth of B. cinerea |
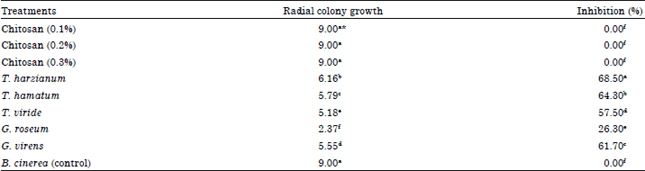 | |
| *Values followed by the same letter(s) in each column do not differ significantly (p≤0.05). Each value represents the mean of 3 replicates | |
The percentages of damping off disease incidence under artificial infection with different isolates of B. cinerea in greenhouse conditions were in the range of 22.53-55.41% and 32.21-64.43%, at pre and post-emergence stages, respectively as shown in Table 1. At pre-emergence stages, the 3rd isolate gave the highest percentage of damping off (55.41%) and the 5th isolate gave the lowest damping off incidence (22.53%) as shown in Table 1. While at post-emergence stages, the 3rd isolate was the highest in percentage of damping off incidence (64.43%) and the 4th isolate was the lowest percentage of damping off incidence (32.21%). So, the third isolate was virulent isolate and used for further studies.
Antagonistic activity of different biocontrol agents against B. cinera on solid medium (dual culture test): In single culture plates, the linear growth of both B. cinerea and fungal antagonists (T. harzianum, T. hamatum, T. viride, G. roseum and G. virens) increased by the time after inoculation. Results presented in (Table 2) suggested that all tested bio-agents had a potent antagonistic effect but it was observed that the linear growth of T. harzianum was the best in inhibition of pathogen growth. In dual culture plates, a great growth inhibition of B. cinerea was occurred after 6 days in comparison with control. Our result explains that significant success in biocontrol is achieved under in vitro conditions. Result showed that Trichoderma species and Gliocladium species could restrict growth of B. cinerea on Potato Dextrose Agar medium (PDA) in the dual culture (Table 2). Trichoderma harzianum was the most potent bioagents where it suppressed the colonization of Botrytis grey mold (BGM) fungus with 68.5% followed by T. hamatum with 64.3% and G. vireins with 61.7% after 6 days in comparison with control. The reduction of mycelial growth of the pathogen in dual culture was not nearly affected by chitosan concentrations as control.
| Table 3: | Effect of different treatments disease incidence and disease severity of cucumber plants infected with grey mould |
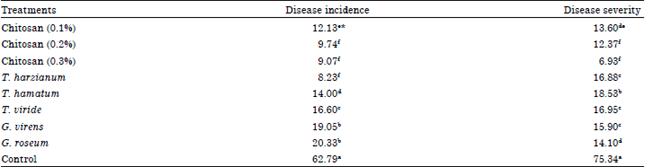 | |
| *Values of each column in each harvest followed by the same letter(s) are not significantly different according to Duncan’s multiple range test (p< = 0.05), each value represents the mean of 3 replicates | |
| Table 4: | Effect of different treatments on growth and yield parameters of cucumber plants infected with grey mould disease |
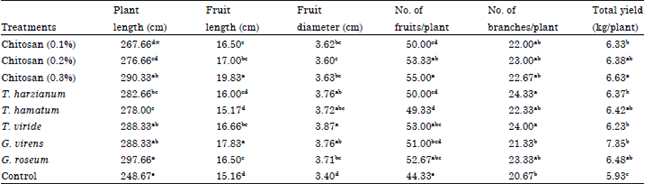 | |
| *Values of each column in each harvest followed by the same letter(s) are not significantly different according to Duncan, s multiple range test (p< = 0.05), each value represents the mean of 3 replicates | |
Present result explains that significant success in biocontrol was achieved under in vitro conditions. Results showed that Trichoderma species and Gliocladium species could restrict growth of B. cinerea on Potato Dextrose Agar medium (PDA) in the dual culture (Table 2). Trichoderma harzianum was the most potent bioagents where it suppressed the colonization of Botrytis Grey Mold (BGM) fungus with 68.5% followed by T. hamatum with 64.3% and G. vireins with 61.7% after 6 days in comparison with control. The reduction of mycelial growth of the pathogen in dual culture was not nearly affected by chitosan concentrations as control.
Greenhouse results
Disease incidence and disease severity results: Data presented in Table 3 showed that all tested treatments have a significant difference with un-treated control in both Disease Incidence (DI) and Disease Severity (DS). Application of T. harzianum gave the most significant reduction in disease incidence by only 8.23% followed by Chitosan 0.2 and 0.3% treatments that gave 9.07 and 9.74%, respectively where the untreated control gave 62.79% disease incidence. For disease severity, chitosan 0.3% treatment was the best in limitation of disease severity by only 6.93 comparing with control that disease severity was estimated to be 75.34%.
Growth and yield parameters: Data presented in Table 4 showed that the growth responses Plant length (cm), Fruit length (cm), Fruit diameter (cm), No. of fruits/plant, No. of branches/plant and total yield (kg/plant) of treated plants were significantly increased in Cucumis sativus plants infected with B. cinerea but the rate of increase was significantly higher in chitosan (0.2%) and chitosan (0.3%) than other bioagents.
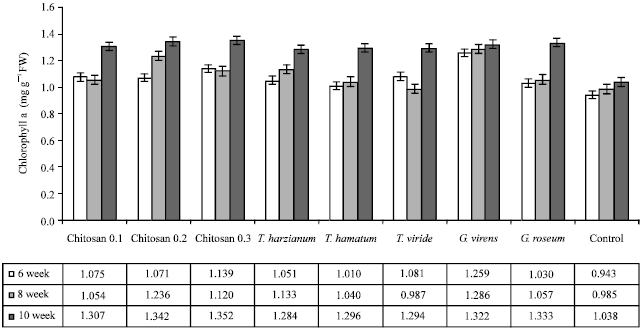 | |
| Fig. 1: | Effect of different treatments on chlorophyll a pigments in the leaves of cucumber plants infected with grey mold disease |
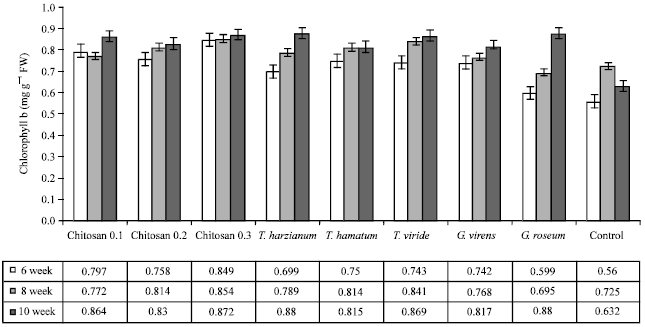 | |
| Fig. 2: | Effect of different treatments on chlorophyll b pigments in the leaves of cucumber plants infected with grey mold disease |
Photosynthetic pigments: The content of photosynthetic pigments (Chlorophyll a, b and carotenoids) in leaves of Cucumis sativus plants is illustrated in Fig. 1, 2 and 3, respectively.
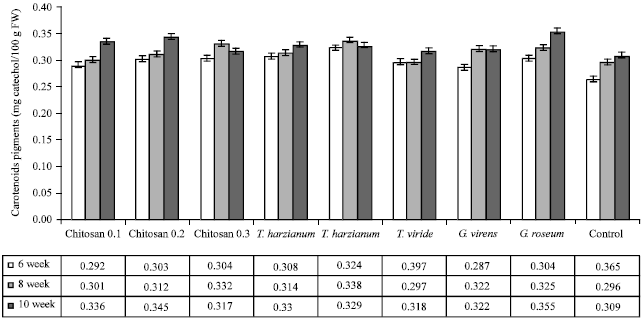 | |
| Fig. 3: | Effect of different treatments on carotenoids pigments in the leaves of cucumber plants infected with grey mold disease |
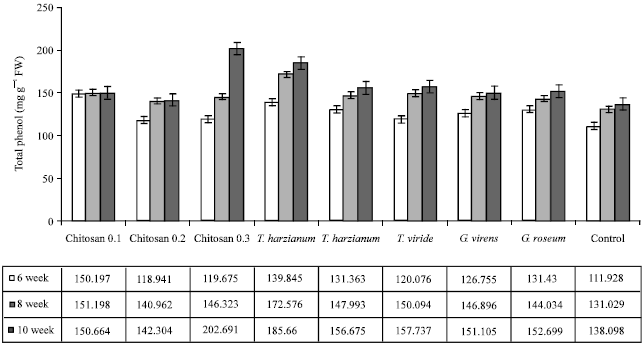 | |
| Fig. 4: | Effect of different treatments on total phenol in the leaves of cucumber plants infected with grey mold disease |
In general, the content of photosynthetic pigments was increased parallel to the increase in plant growth in all treatments but the content of photosynthetic pigments was greatly higher in T. harzianum treatment. Also it was noticeable that chitosan 0.3% showed a significant increase in Chlorophyll a, b and carotenoids.
Total phenol content: The effect of bioagents application on the total phenol contents of infected by B. cinerea is described in Fig. 4.
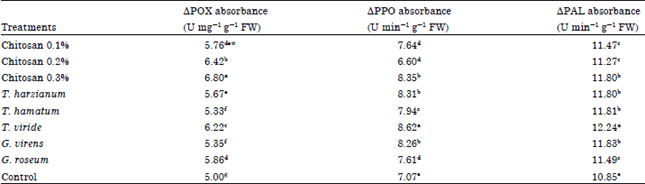
| Table 5: | Effect of different treatments on ΔPOX absorbance unit mg g-1 FW, ΔPPO absorbance unit min-1 g-1 FW and ΔPAL absorbance unit min-1 g-1 FW |
 | |
| *Values followed by the same letter(s) in each column do not differ significantly (p≤0.05). Each value represents the mean of 3 replicates | |
The foliar application of T. harzianum, chitosan 0.2% and chitosan 0.3% treatments significantly increased phenol contents of infected plants when compared with the untreated control plants. On the other hand, no significant differences were noticed between these treatments.
Assay of defense related enzymes activity: Data presented in Table 5 showed that defense related enzymes activity was significantly high in infected cucumber plants that were treated with antagonistic fungi especially T. harzianum. Then again, all chitosan treatment caused a significant increase on the activity of the screened enzymes mainly at higher concentration.
DISCUSSION
The prevalence of favorable environmental conditions such as temperature (3-28°C) and relatively high humidity (41-93%) prevailed during the period from December to February and apparently favored the pathogenesis of B. cinerea leading to substantial development of the disease. B. cinerea is a foliar pathogen under natural conditions (Elad, 1996; Davidson et al., 2004) but it could be survive in soil as a saprophytic fungus or on the remain of infected plant debris and can cause systemic infection through roots. In the past, chemical fungicides were commonly used in the control of plant fungal diseases (Nemec, 1980). Grey mold control using these chemicals produced successive, stronger and more extensive fluctuation in disease incidence that caused greater crop losses in sometimes. In addition, continuous using of these chemicals may not always be desirable due to their toxic effects on non-target organisms that could be beneficial, beside environmental pollution problems. Moreover, development of plant resistant pathogen isolates could cause disease outbreaks from time to time. Many of these chemical pesticides are also relatively expensive for the small farmers (Vasudevan et al., 2002). Based on the above, our study was planned to find some alternative to serve as substitute for the undesirable and unsafe chemical control. Taking in consideration, the effectiveness, the cost and environmental safety for these options for Botrytis cinerea control. The biocontrol agents have not harmful effects on humans, wild life or any other beneficial organisms, safe and effective biocontrol agents in both natural and controlled environments, in addition to no residues advantage in the food chain (Wiest et al., 2002). One of the essential characters of fungal biological control agents is to act, as mycoparasites against fungal plant pathogens due to their excreted hydrolytic enzymes (Chet, 1987; Hermosa et al., 2000). The seed-borne nature of B. cinerea and its ability to survive in soil makes it difficult challenge to control but the existing study has revealed that treatment of Trichoderma spp. and Gliocladium spp. effectively manage the disease.
Application of Trichoderma species, especially T. harzianum significantly controlled grey mold in tested cucumber plants under the nature infection. The antagonistic activity of these fungi could be owed to several mechanisms. Ramamoorthy et al. (2002) and Liu et al. (2004) suggested that producing some antimicrobial toxins could play an important role on the antagonism process. On the other hand, producing some plant growth promoting substances could be one of the mechanisms (Vessey, 2003). These growth promotors could enhance the overall growth condition beside its influence in inducing host resistance (Bigirimana et al., 1997; De Meyer et al., 1998). Benitez et al. (2004) stated that application of appropriate Trichoderma formulations can promote plant growth and crop precocity, increase legume production and reduce chemical treatments. T. harzianum also can antagonist the plant pathogens by competing with them on nutrients and space. It was found that most of Trichoderma spp. isolates were able to colonizing the soil and/or plant parts, occupying a physical space and avoiding the multiplication of the pathogens, in addition, modifying the environmental conditions, or promoting plant growth (Inbar et al., 1994) and induce plant defensive mechanisms (Monte and Liobell, 2003) and antibiosis (Keszler et al., 2000). Moreover, Cherif and Benhamou (1990) besides Harman and Bjorkman (1998) reported some direct mechanisms such as mycoparasitism and production of cell wall degrading enzymes against the pathogens. In the present study, T. harzianum found to be more effective than G. vireins. Khan and Gupta (1998) reported relatively greater control of M. phaseolina with T. harzianum than G. vireins. De Meyer et al. (1998) has also reported that T. harzianum to be more effective than G. vireins against plant pathogenic fungi. The chitinolytic system of T. harzianum comprises 6 distinct enzymes 2 of them are classified as acetyl glucosaminidases and the rest as endochitinases by Haran et al. (1995). Chitinase enzyme is concerned in the degradation of fungal cell wall and hence the lysis of target plant pathogens which leading to hyphal penetration by mycoparasites (Haran et al., 1996; Baek et al., 1999; Kumar and Gupta, 1999; Woo et al., 1999). In addition to the mycoparasitic function, it was noticed that extracellular chitinase may affect the biological activity of Trichoderma through other mechanisms such as inhibition of the pathogens growth and/or induction of host plant resistance. Accordingly, the increased activity of biocontrol could be partially due to increase of chitinase production. Howell (2003) and Harman (2006) reported the beneficial effects of G. vireins on plants comprise their ability to antagonize soil-borne pathogens by a combination of producing enzymatic lysis, secretion of antibiotics and competition for space and substrates. In the current study, also chitosan was tested as potential alternative to control plant pathogens. Using of such natural compounds may lead to decrease in consumption of harmful, chemical fungicides. Chitosan offers a great prospective as a biodegradable material that may have anti-microbial and elicitation activities. Benhamou (1996) and Oh et al. (1998) suggested that chitosan could have a dual effect on the host-pathogen interaction antifungal activity and activation of plant defense responses. During our tests, any antimicrobial activity of chitosan was not found against B. cinerea, this could be due to low tested concentration or the testing method. Lafontaine and Benhamou (1996) found that chitosan could affect the total yield of infected plants. They reported a total yield increase of tomato was highly correlated with the concentration of chitosan applied to soil inoculated with F. oxysporum f. sp. radicis-lycopersici. However, when fungal inoculation was not involved, fruit yield from chitosan-treated soil and untreated was similar. These finding leads to the fact that yield increasing was related to chitosan activity in disease reduction.
Although, Chitosan could have interference with the growth of some phytopathogenic fungi including B. cinerea (Du et al., 1997; El Ghaouth et al., 1994; Oh et al., 1998) but until now the mechanism by which it affects the growth of the pathogen is still unclear. Benhamou (1996) suggested that chitosan may interfere with the negatively charged residues of macromolecules exposed on the fungal cell surface. This is might lead to changes on the permeability of the fungal plasma membrane. Elicitation mechanism of chitosan was noticed as effect on several defense-related enzymes (Abeles and Forrence, 1979; Inui et al., 1997; Vander et al., 1998). These enzymes known to be participate in early defense mechanisms and to prevent pathogen infections (Smith, 1996). It has been reported that chitosan could stimulate other systems involved in resistance, such as phenylalanine ammonia lyase activities, lipoxygenase and lignin formation in wheat leaves (Bohland et al., 1997; Vander et al., 1998).
In our present study, it was observed that the growth parameters, carbohydrates, photosynthetic pigments, proline, phenol contents and yield parameters were significantly increased as compared to the equivalent untreated plants. On the other hand, it was noticed that application of these bioagents to the infected Cucumber plants significantly reduced both disease incidence and disease severity as compared to the untreated plants. It is evident from the present study that infection with the foliar pathogen B. cinerea significantly reduced the content of photosynthetic pigments in the leaves of Cucumber plants, this reduction may be due to the effect of pathogen on the chloroplasts or on chlorophyll content directly or on the enzymes related with photosynthesis. Significant increase in phenolic compounds in leaves was reported. These increase may be due to inhibition of catalase activity which in turn induces phenylalanine lyase gene expression and synthesis of phenolic compounds according to Vermerris and Nicholson (2006). It is well known that total phenols have long been considered as important defense-related compounds whose levels are naturally high in resistant varieties of many crops (Gogoi et al., 2001; Khaleifa et al., 2006).
REFERENCES
- Abeles, F.B. and L.E. Forrence, 1970. Temporal and hormonal control of β-1,3-glucanase in Phaseolus vulgaris L. Plant Physiol., 45: 395-400.
PubMedDirect Link - Agrios, G.N., 1997. Plant Pathology. 4th Edn., Academic Press, Cambridge, Massachusetts, ISBN-13: 9780120115648, Pages: 635.
Direct Link - Baek, J.M., C.R. Howell and C.M. Kenerley, 1999. The role of an extracellular chitinase from Trichoderma virens Gv29-8 in the biocontrol of Rhizoctonia solani. Curr. Genet., 35: 41-50.
CrossRefPubMedDirect Link - Baker, R., Y. Elad and I. Chet, 1984. The controlled experiment in the scientific method with special emphasis on biological control. Phytopathology, 74: 1019-1021.
Direct Link - Benhamou, N., 1996. Elicitor-induced plant defence pathways. Trends Plant Sci., 1: 233-240.
CrossRefDirect Link - Benhamou, N., P.J. Lafontaine and M. Nicole, 1994. Induction of systemic resistance to Fusarium crown and root rot in tomato plants by seed treatment with chitosan. Phytopathology, 84: 1432-1444.
CrossRefDirect Link - Benitez, T., A.M. Rincon, M.C. Limon and A.C. Codon, 2004. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol., 7: 249-260.
PubMedDirect Link - Bohland, C., T. Balkenhohl, G. Loers, I. Feussner and H.J. Grambow, 1997. Differential induction of lipoxygenase isoforms in wheat upon treatment with rust fungus elicitor, chitin oligosaccharides, chitosan and methyl jasmonate. Plant Physiol., 114: 679-685.
Direct Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Brueske, C.H., 1980. Phenylalanine ammonia lyase activity in tomato roots infected and resistant to the root-knot nematode, Meloidogyne incognita. Physiol. Plant Pathol., 16:: 409-414.
CrossRefDirect Link - Cherif, M. and N. Benhamou, 1990. Cytochemical aspects of chitin breakdown during the parasitic action of a Trichoderma sp. on Fusarium oxysporum f. sp. radicis-lycopersici. Phytopathology, 80: 1406-1414.
Direct Link - De Meyer, G., J. Bigirimana, Y. Elad and M. Hofte, 1998. Induced systemic resistance in Trichoderma harzianum T39 biocontrol of Botrytis cinerea. Eur. J. Plant Pathol., 104: 279-286.
CrossRef - Du, J.M., H. Gemma and S. Iwahori, 1997. Effects of chitosan coating on the storage of peach, Japanese pear and kiwifruit. J. Jpn. Soc. Hortic., 66: 15-22.
CrossRefDirect Link - Eikemo, H., A. Stensvand and A.M. Tronsmo, 2003. Induced resistance as a possible means to control diseases of strawberry caused by Phytophthora spp. Plant Dis., 87: 345-350.
CrossRefDirect Link - Elad, Y., N.E. Malathrakis and A.J. Dik, 1996. Biological control of Botrytis-incited diseases and powdery mildews in greenhouse crops. Crop Prot., 15: 229-240.
CrossRefDirect Link - Elad, Y., 1996. Mechanisms involved in the biological control of Botrytis cinerea incited diseases. Eur. J. Plant Pathol., 102: 719-732.
CrossRefDirect Link - Davidson, J.A., S. Pande, T.W. Bretag, K.D. Lindbeck and G.K. Kishore, 2004. Biology and Management of Botrytis spp. in Legume Crops. In: Botrytis: Biology, Pathology and Control, Elad, Y., B. Willium, P. Tudzynski and N. Delen (Eds.). Chapter 16, Kluwer Academic Publishers, The Netherlands, ISBN-13: 978-1402026249, pp: 295-318.
Direct Link - Elad, Y., H. Yunis and T. Katan, 1992. Multiple fungicide resistance to benzimidazoles, dicarboximides and diethofencarb in field isolates of Botrytis cinerea in Israel. Plant Pathol., 41: 41-46.
CrossRefDirect Link - Elad, Y., G. Zimand, Y. Zaqs, S. Zuriel and I. Chet, 1993. Use of Trichoderma harzianum in combination or alternation with fungicides to control cucumber grey mould (Botrytis cinerea) under commercial greenhouse conditions. Plant Pathol., 42: 324-332.
CrossRefDirect Link - El Ghaouth, A., J. Arul, C. Wilson and N. Benhamou, 1994. Ultrastructural and cytochemical aspects of the effect of chitosan on decay of bell pepper fruit. Physiol. Mol. Plant Pathol., 44: 417-432.
CrossRefDirect Link - Gogoi, R., D.V. Singh and K.D. Srivastava, 2001. Phenols as a biochemical basis of resistance in wheat against karnal bunt. Plant Pathol., 50: 470-476.
CrossRefDirect Link - Harman, G.E. and T. Bjorkman, 1998. Potential and Existing Uses of Trichoderma and Gliocladium for Plant Disease Control and Plant Growth Enhancement. In: Trichoderma and Gliocladium, Volume 2: Enzymes, Biological Control and Commercial Applications, Harman, G.E. and C.P. Kubicek (Eds.). Taylor and Francis, London, UK., ISBN-13: 9780203484456, pp: 229-265.
- Harman, G.E., 2006. Overview of mechanisms and uses of Trichoderma spp. Phytopathology, 96: 190-194.
CrossRefDirect Link - Hermosa, M.R., I. Grondona, E.A. Iturriaga, J.M. Diaz-Minguez, C. Castro, E. Monte and I. Garcia-Acha, 2000. Molecular characterization and identification of biocontrol isolates of Trichoderma spp. Applied Environ. Microbiol., 66: 1890-1898.
CrossRefDirect Link - Howell, C.R., 2003. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The history and evolution of current concepts. Plant Dis., 87: 4-10.
CrossRefDirect Link - Inbar, J., M. Abramsky, D. Cohen and I. Chet, 1994. Plant growth enhancement and disease control by Trichoderma harzianum in vegetable seedlings grown under commercial conditions. Eur. J. Plant Pathol., 100: 337-346.
CrossRefDirect Link - Inui, H., Y. Yamaguchi and S. Hirano, 1997. Elicitor actions of N-acetylchitooligosaccharides and laminarioligosaccharides for chitinase and L-Phenylalanine ammonia-lyase induction in rice suspension culture. Biosci. Biotechnol. Biochem., 61: 975-978.
CrossRefDirect Link - Keszler, A., E. Forgacs, L. Kotai, J.A. Vizcaino, E. Monte and I. Garcia-Acha, 2000. Separation and identification of volatile components in the fermentation broth of Trichoderma atroviride by solid-phase extraction and gas chromatography-mass spectrometry. Chromatogr. Sci., 38: 421-424.
CrossRefDirect Link - Khaleifa, M.M.A., C.R. Azzam and S.A. Azer, 2006. Biochemical markers associated with disease resistance to damping-off and root-rot diseases of peanut mutants and their productivity. Egypt. J. Phytopathol., 34: 53-74.
Direct Link - Khan, M.R. and J. Gupta, 1998. Antagonistic efficacy of Trichoderma species against Macrophomina phaseolina on eggplant. J. Plant Dis. Prot., 105: 387-393.
Direct Link - Kumar, A. and J.P. Gupta, 1999. Variations in enzyme activity of tebuconazole tolerant biotypes of Trichoderma viride. Indian Phytopathol., 52: 263-266.
Direct Link - Lafontaine, P.J. and N. Benhamou, 1996. Chitosan treatment: An emerging strategy for enhancing resistance of greenhouse tomato plants to infection by Fusarium oxysporum f.sp. radicis-lycopersici. Biocontrol Sci. Technol., 6: 111-124.
CrossRefDirect Link - Liu, X., J.H. Zhuang, Z.G. Gao, C.C. Yang and J. Chen, 2004. Construction of engineering Trichoderma strains and their characteristics against tomato gray mold. J. Zhejang Univ., 30: 419-420.
Direct Link - Mackinney, G., 1941. Absorption of light by chlorophyll solutions. J. Biol. Chem., 104: 315-322.
Direct Link - Monte, E. and A. Liobell, 2003. Trichoderma in organic agriculture. Proceedings of the 5th World Avocado Congress, October 19-24, 2003, Granada, Spain, pp: 725-733.
Direct Link - Nemec, S., 1980. Effects of 11 fungicides on endomycorrhizal development in sour orange. Can. J. Bot., 58: 522-526.
CrossRefDirect Link - Oh, S.K., D. Cho and S.H. Yu, 1998. Development of integrated pest management techniques using biomass for organic farming (I). Plant Pathol. J., 14: 278-285.
Direct Link - Raju, M.R.B. and K.V.M.K. Murthy, 2000. Efficacy of Trichoderma spp. in the management of collar rot of groundnut caused by Aspergillus niger Van Tieghem. Indian J. Plant Prot., 28: 197-199.
Direct Link - Ramamoorthy, V., T. Raguchander and R. Samiyappan, 2002. Enhancing resistance of tomato and hot pepper to Pythium diseases by seed treatment with fluorescent pseudomonads. Eur. J. Plant Pathol., 108: 429-441.
CrossRefDirect Link - Rojo, F.G., M.M. Reynoso, M. Ferez, S.N. Chulze and A.M. Torres, 2007. Biological control by Trichoderma species of Fusarium solani causing peanut brown root rot under field conditions. Crop Protect., 26: 549-555.
CrossRefDirect Link - Haran, S., H. Schickler, A. Oppenheim and I. Chet, 1995. New components of the chitinolytic system of Trichoderma harzianum. Mycol. Res., 99: 441-446.
CrossRefDirect Link - Singleton, V.L. and J.A. Rossi, 1965. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic., 16: 144-158.
CrossRefDirect Link - Smith, C.J., 1996. Tansley review No. 86 accumulation of phytoalexins: Defence mechanism and stimulus response system. New Phytol., 132: 1-45.
CrossRefDirect Link - Solfrizzo, M., C. Altomare, A. Visconti, A. Bottalico and G. Perrone, 1994. Detection of peptaibols and their hydrolysis products in cultures of Trichoderma species. Nat. Toxins, 2: 360-365.
Direct Link - Sticher, L., B. Mauch-Mani and J.P. Metraux, 1997. Systemic acquired resistance. Annu. Rev. Phytopathol., 35: 235-270.
CrossRefPubMedDirect Link - Ten Have, A., K.B. Tenberg, J.A.E. Benen, P. Tudzynski, J. Visse and J.A.L. van Kan, 2002. The Contribution of Cell Wall Degrading Enzymes to Pathogenesis of Fungal Plant Pathogens. In: Agricultural Applications (The Mycota, Volume 11), Kempken, F. (Ed.). Springer Science and Business Media, New York, USA., ISBN-13: 978-3540426288, pp: 341-358.
- Vander, P., K.M. Varum, A. Domard, N.E. El Gueddari and B.M. Moerschbacher, 1998. Comparison of the ability of partially N-acetylated chitosans and chitooligosaccharides to elicit resistance reactions in wheat leaves. Plant Physiol., 118: 1353-1359.
CrossRefDirect Link - Vessey, J.K., 2003. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil, 255: 571-586.
CrossRefDirect Link - Wiest, A., D. Grzegorski, B.W. Xu, C. Goulard and S. Rebuffat et al., 2002. Identification of peptaibols from Trichoderma virens and cloning of a peptaibol synthetase. J. Biol. Chem., 277: 20862-20868.
CrossRefDirect Link - Woo, S.L., B. Donzelli, F. Scala, R. Mach and G.E. Harman et al., 1999. Disruption of the ech42 (Endochitinase-Encoding) gene affects biocontrol activity in Trichoderma harzianum P1. Mol. Plant-Microbe Interact., 12: 419-429.
CrossRefDirect Link - Lurie, S., E. Fallik, A. Handros and R. Shapira, 1997. The possible involvement of peroxidase in resistance to Botrytis cinereain heat treated tomato fruit. Physiol. Mol. Plant Pathol., 50: 141-149.
CrossRef - Mukherjee, P.K. and M.P. Haware, 1993. Biological control of Botrytis gray mold of chickpea. International Chickpea Newslett., 28: 14-15.
Direct Link - Cohen, Y., U. Gisi and E. Mosinger, 1991. Systemic resistance of potato plants against Phytophthora infestans induced by unsaturated fatty acids. Physiol. Mol. Plant Pathol., 38: 255-263.
CrossRefDirect Link