
Y.Y. Molan
Department of Plant Protection, College of Food and Agriculture Science, King Saud University, P.O. Box 2460, Riyadh 11451, Saudi Arabia
Asian Journal of Plant Pathology
Year: 2009 | Volume: 3 | Issue: 1 | Page No.: 22-26







ABSTRACT
The presumptive mycoparasites Trichoderma sp., Pythium sp., Gliocladium sp. and Verticillium sp. were detected in 42 (51.2%), 2 (2.5%), 23 (28.4%) and 1 (1.2%) of a total 81 soils, respectively, when soil samples were placed on sectors of potato dextrose agar colonized by the appropriate host fungi. Most (81) of the soils in the study contained three or more mycoparasities, but the frequency of detection on replicate host sectors suggested that Trichoderma sp. and Gliocladium sp. were the more abundant species in all soils in which they occurred. The type of host fungus markedly influenced the efficiency of detection of the different mycoparasities: Fusarium sp. was most efficient for Trichoderma sp. and Gliocladium sp. and Rhizoctonia solani for Trichoderma sp. Only a single host was suitable for consistent detection of each of the mycoparasite species.
PDF Abstract XML References Citation
How to cite this article
Y.Y. Molan, 2009. Detection of Presumptive Mycoparasites in Soil Placed on Host-Colonized Agar Plates in Riyadh Region, Saudi Arabia. Asian Journal of Plant Pathology, 3: 22-26.
URL: https://scialert.net/abstract/?doi=ajppaj.2009.22.26
URL: https://scialert.net/abstract/?doi=ajppaj.2009.22.26
INTRODUCTION
Soil-borne pathogens produce serious to the yield of losses in different plant groups (Iftikhar et al., 2003). Micro-organisms that colonized in the rhizosphere were classified according to their effects on plants, some of these micro-organisms considered plant pathogens, whereas others trigger beneficial effects (Mantellin and Touraine, 2004). Fungi with antagonistic activity toward plant pathogens had an essential role in plant growth and health. A plant and site dependent specificity of the composition of antagonistic morphotypes and their genotypic diversity was found by Berg et al. (2005).
Mycoparasites and presumptive mycoparasites have biocontrol potential some are responsible for natural suppressiveness of soils to certain plant pathogens, examples being Pythium nunn in Pythium-suppressive soils (Lifshitz et al., 1984; Martin and Hancock, 1986) and Trichoderma hamatum in Rhizoctonia suppressive soils (Liu and Baker, 1980). Many studies showed that root colonization by specific fungi was found recently gave increases in plant growth, yield or a measurable control of known root pathogens (Cook, 1993; Cook and Baker, 1983; Linderman, 1986). Fusarium species were economically important as pathogens on many agricultural, horticultural and field crops grown in the world. Chet (1987) showed that several species of Trichoderma were used successfully against certain pathogenic fungi. Similarly, Harman (2000) showed that Trichoderma sp. was used as commercial bio-fungicides to control a range of economically important soil-borne fungal plant pathogens. The objective of this study was to isolate the mycoparasitic fungi from soils of the region, Saudi Arabia, that might be used as biological control agents as an alternative to chemical control methods.
MATERIALS AND METHODS
This study was conducted in 2004-2005 at the Plant Protection Department, College of Food and Agriculture Science, King Saud University, Riyadh, Saudi Arabia.
Isolation of the Host Fungi
Initial studies (not described) involved comparisons of many potential host fungi which were isolated from different host plants. Then 6 hosts were selected to isolate the mycoparasities populations in 81 soils. These hosts were Fusarium proliferatum, Fusarium sp., Botrytis cinerea, Sclerotinia sp., Rhizoctonia solani and Pythium sp. Identification of the different types of developed fungal isolates colonies were carried out and identified based on microscopic morphologies according to Ellis (1971), Kirk and Ansell (1992) and Samson and Pitt (2000).
Isolation of Mycoparasites from Soil
Eighty one soils were collected from arable fields (47), garden sites (19) and woodlands (15) in the Riyadh region of Saudi Arabia (Table 1, 2). Each sample was bulked from usually 2 or 3 subsamples which mixed thoroughly and stored for up to 1 week in a polyethylene bags at room temperature.
| Table 1: | Occurrence of different mycoparasites in 81 soils as determined by detection on agar previously colonized by appropriate host fungi in Al-Dereia, Al- Oiaina and Al Wasil wa Al Amaria regions, Saudi Arabia |
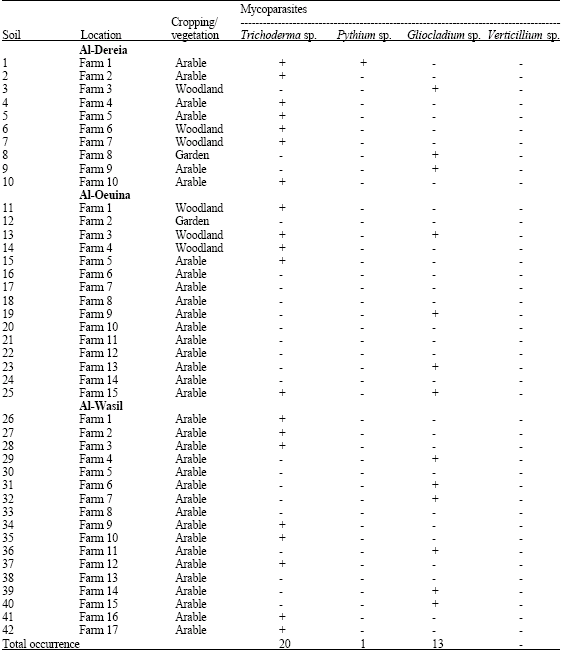 | |
| +: Present; -: Absent | |
| Table 2: | Occurrence of different mycoparasites in 81 soils as determined by detection on agar previously colonized by appropriate host fungi in Al- Kharj, Dayrab and Wadi Hanifa regions, Saudi Arabia |
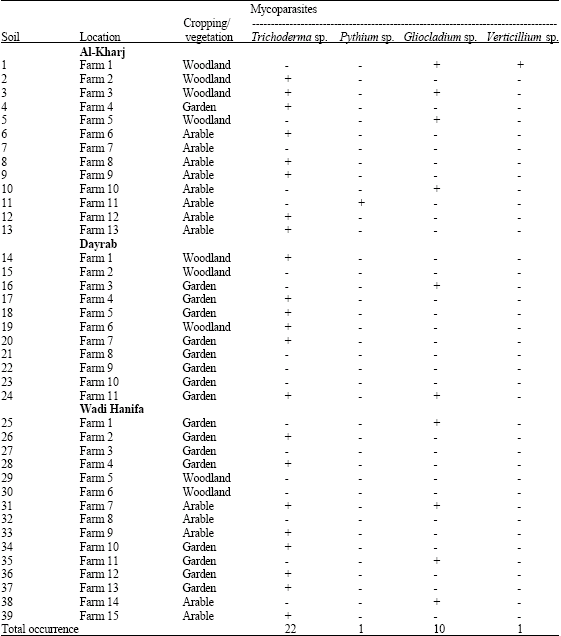 | |
| +: Present; -: Absent | |
Plates of Potato-Dextrose Agar (PDA, 15 mL per 9 cm diam. plate) were inoculated in the centrally with the different host fungi and incubated at 25°C for different time periods, until the colony margin just reached the edge of the plate. Then, each agar was cut into 6 equal sectors, which were placed separately in sterilized plastic petri dishes. A sample of soil (0.4 mL, ca. 0.4 g), was placed on the oldest part of each host fungal sector (Mullingan and Deacon, 1992). The sectors were incubated again at 25°C and examined after 7, 14 and 21 days of incubation. Mycoparasities were detected by the presence of sporulation or other fungal structures on the host colony. Identification were confirmed with pure cultures, obtained by subculturing of the mycoparasities from representative host sectors onto plates of fresh PDA. The host sectors were selected at randomized, so for any one soil they were usually from different colonies.
RESULTS AND DISCUSSION
The presumptive mycoparasities Trichoderma sp., Pythium sp., Gliocladium sp. and Verticillium sp. were detected in 42, 2, 23 and 1 of a total 81 soils, respectively of a total 81 soils when samples were placed on sectors of agar colonized by the appropriate host fungi (Table 1, 2). Trichoderma sp. and Gliocladium sp. were the more abundant species in all soils in which they occurred for the soils. Of the 6 hosts used, Fusarium sp. was more efficient for Trichoderma sp., Rhizoctonia solani for Trichoderma sp., Fusarium sp., for Gliocladium sp. Mullingan and Deacon (1992) reported that, all earlier studies researchers have used a single host to identify presumptive mycoparasites and thus probably underestimated the presence of mycoparasities in soils (Foley and Deacon, 1985). Also, mycoparasites could be detected and identified to at least generic level without the need for subculturing (Mullingan and Deacon, 1992). Fusarium sp., Botrytis cinerea and Rhizoctonia solani were gave different patterns of detection of mycoparasities (Table 3). Trichoderma sp. and Gliocladium sp., were detected on all areas and 2 hosts. Trichoderma sp. was seen mostly on all hosts except Pythium sp., Verticillium sp. was seen only on Rhizoctonia solani. Comparison of Table 1 and 2 shows that Pythium sp. was reported only in Al-Dereia and Al- Kharj regions for one time. Only a single host was suitable for consistent detection of one single mycoparasitie. These results cannot be ascribed to chance, because in several soils a mycoparasities that was not detected. Table 3 shows that the greatest likelihood of detecting all the mycoparasities in any one soil was by the use of Fusarium sp., for Trichoderma sp., Rhizoctonia solani for Trichoderma sp., Fusarium sp., for Gliocladium sp. The hosts in this study were selected for rapid, uniform growth on PDA and for the known abilities of mycoparasities to grow across them in culture (Laing and Deacon, 1991). Theses abilities were precisely matched by the results of the soil survey. The hosts for future surveys could be selected on this basis, or specific pathogens could be used to detected potential biocontrol agents (Van den Boogert et al., 1990). But the best host for detection of a mycoparasities may not be the best for its isolation into pure culture. Trichoderma was detected in all 42 soils, Gliocladium sp. in 23 soils, Pytuhium sp. in 2 soils and Verticillium sp. in one soil (Table 3). Moreover, all soils had more than one mycoparasities: 8 soils contained three types, 33 contained 3 types and 4 contained 1 type.
| Table 3: | Incidence of detection of different mycoparasites in soils samples were placed on agar colonized by different host fungi |
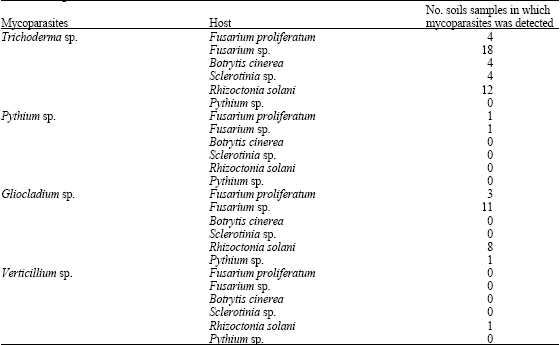 | |
As an estimate of the abundance of each mycoparasities in each soil, Table 1 and 2 shows, the cases in which a mycoparasite was detected on at least half of the replicate sectors of any 1 host. Overall, Trichoderma sp. was detected in 51% of the samples of soil placed on host colonies and Gliocladium sp. in 28% of all samples. In present study, only some of previously reported soil-borne mycoparasites were detected (Deacon and Henry, 1978; Foley and Deacon, 1985). Further study, with different hosts and a broader geographical or ecological rang of sites should resolve these issues.
REFERENCES
- Berg, G., C. Zachow, J. Lottmann, M. Gotz, R. Costa and K. Smalla, 2005. Impact of plant species and site on rhizosphere associated fungi antgonistic to Verticillium dahliae kleb. Applied Environ. Microbiol., 71: 4203-4213.
CrossRefDirect Link - Cook, R.J., 1993. Making greater use of introduced microorganisms for biological control of plant pathogens. Annu. Rev. Phytopathol., 31: 53-80.
CrossRef - Harman, G.E., 2000. Myths and Dogmas of Biocontrol Changes in Perceptions Derived from Research on Trichoderma harzinum T-22. Plant Dis., 84: 377-393.
CrossRefDirect Link - Iftikhar, S., A. Sultan, A. Munir, S. Iram and I. Ahmed, 2003. Fungi associated with rice, wheat cropping system in relation to zero and conventional tillage technologies. J. Biol. Sci., 3: 1076-1083.
Direct Link - Mantelin, S. and B. Touraine, 2004. Plant growth-promoting bacteria and nitrate availability: Impacts on root development and nitrate uptake. J. Exp. Bot., 55: 27-34.
CrossRefDirect Link