
Saruchi Sharma
Parasitology and Entomology Laboratory, Department of Biosciences, Himachal Pradesh University, Shimla, 171005, India
D.R. Thakur
Parasitology and Entomology Laboratory, Department of Biosciences, Himachal Pradesh University, Shimla, 171005, India
American Journal of Food Technology
Year: 2014 | Volume: 9 | Issue: 6 | Page No.: 318-324







ABSTRACT
Legume seeds are rich and varied source of secondary plant metabolites which are toxic or anti metabolic towards insect pests, possibly by virtue of these toxic substances they show resistance and susceptibility against stored grain pest species. Despite the potential nutritional and health promoting value the presence of antinutritional factors limit biological value and usage of legumes as food. Antinutritional factors interfere with digestion and also make the seeds unpalatable when consumed in raw form. Based on these possibilities the biochemical basis of bruchid resistance has been studied in different legume genotypes and the results recorded revealed that highly resistant soybean genotypes possesed high amount of fats, proteins and antinutritional factors (phenols and 4-5 times more trypsin inhibitors) than cowpea and chickpea (kabuli>desi) genotypes which contain high amount of carbohydrates and low amount of antinutritional factors and were susceptible towards Callosobruchus species.
PDF Abstract XML References Citation
Received: April 20, 2014;
Accepted: July 27, 2014;
Published: October 23, 2014
How to cite this article
Saruchi Sharma and D.R. Thakur, 2014. Biochemical Basis for Bruchid Resistance in Cowpea, Chickpea and Soybean
Genotypes. American Journal of Food Technology, 9: 318-324.
DOI: 10.3923/ajft.2014.318.324
URL: https://scialert.net/abstract/?doi=ajft.2014.318.324
DOI: 10.3923/ajft.2014.318.324
URL: https://scialert.net/abstract/?doi=ajft.2014.318.324
INTRODUCTION
A study on the relative susceptibility of cowpea, chickpea and soybean genotypes against bruchids (C. maculatus and C. chinensis) was undertaken and significant difference among the genotypes were observed with regard to their relative susceptibility to pests. The variation in seed parameters was primarily due to variation in percentage infestation level, adult emergence, reduction in seed weight and inherent capacity of each genotype to be attacked by bruchid. The results of parameters (Growth performance of C. maculatus and C. chinensis) studied pertaining to resistance varied significantly among different genotypes of three legumes used in present study, which indicated that the variations in different parameters may be due to genetic factors, possible presence of biochemical contents of seeds such as antibiotics, tannin contents, trypsin inhibitiors, phenol content etc. or presence of low nutritional components and high antinutritional components in seeds. Some stored legumes are not often attacked by bruchids and preference of pests to those commodities has been noted at two levels. First is host selection by the pest species which is sensory to a large extent and second, the ability to complete development on the selected host (Janzen, 1977).
Many studies have indicated that some chemical factors are responsible for pest resistance. Adjadi et al. (1985) studied the genetics of bruchid resistance in cowpea and observed that two recessive genes (rcm1, rcm1 and rcm2, rcm2) in homozygous condition were responsible resistance to bruchid pests. Despite the potential nutritional and health promoting value the presence of antinutritional factors limit biological value and usage of legumes as food. Antinutritional factors interfere with digestion and also make the seeds unpalatable when consumed in raw form. These factors can be divided into two categories as protein and non-protein factors. Non protein anti-nutritional factors include alkaloids, tannins, phytic acid, sapponins and phenols while Trypsin inhibitors, chymotrypsin inhibitors lectins and antifungal peptides are protein anti-nutritional factors (Duranti and Gius, 1997). Certain morphological and physiological characteristics inherited by plants from a core of defense mechanism against insects and other pest and pathogens that would otherwise attack them. These defenses are the result of long term natural selection. These morphological features either produce physical stimuli or inhibit insect activity. From the gene pool of a crop species, certain crosses produce phenotypes that vary from complete susceptibility to high levels of resistance against insects. Through biochemical estimation of legumes, it is easy to evaluate the components discourage or encourage the bruchids to infest the legume seeds. Therefore, vulnerable legume cultivars can be genetically modified and resistant strains can be engineered. Several chemical factors, protein anti-metabolites which include protease inhibitors, lecithins, apart from physical factors have been found to confer resistance to legumes against pest species are controlled by single gene or genes of the same family, hence can be used for developing transgenic species. The other secondary plant compounds such as polyphenols and tannins which play important role in conferring resistance to legumes have not progressed much because of the fact that they are multigenic factors and controlled by different genes. A study was thus necessary regarding the control factors so as to be incorporated in evolving pest control strategies.
The present study includes the biochemical analysis including quantitative estimation of nutritional and anti nutritional components in different cultivars of Vigna sinensis, Glycine max L. (Merill) and Cicer arietinum, which probably are responsible for susceptibility and resistance against C. maculatus and C. chinensis. Based on these possibilities the biochemical basis of bruchid resistance has been studied in these legume genotypes.
MATERIALS AND METHODS
For biochemical estimation, samples of thirteen genotypes of soyabean (G. max) viz., JS-335, JS-9305, JS-9560, Bragg, Palamsoya, P2-2, Pb-1, Himsoya, P9-2-2, Shivalik, Harasoya, PK-472, P13-4, were obtained from CSK Himachal Pradesh Krishi Vishvavidalaya, Palampur, samples of eight chickpea (C. arietinum) cultivars viz., HK-1, HK-2, HC-1, HC-5, C-235, GNG-663, GNG-1481 and GNG-1581 were obtained from CCS, Agriculture University, Hisar and samples of ten genotypes of cowpea (V. sinensis) viz., IC-106816, IC-106817, IC-106819, IC-106826, IC-106831, IC-106835, IC-106839, IC-326634, IC-326996 and IC-311584, from National Bereau of Plant Genetic Resources (NBPGR) Phagli, Shimla for the experimental studies. Then samples of different cultivars of each legumes were homogenized and analyzed for different nutritional components (carbohydrates, proteins and lipids) and anti-nutritional components (phenols, tannin and trypsin inhibitor).
Homogenization: Homogenates of different cultivars seed samples (both normal and infested) were prepared in cold PBS (Phosphate buffer saline, pH 7.2) in Remi motor homogenizer. First the samples were grinded in electric grinder and 1 g (w/w) amount of each seed genotype mixed with PBS and transferred to homogenizer at 3000-3500 rpm.
Centrifugation: Homogenate was then centrifuged in a centrifuge at 3000-3500 rpm for 10 min. Then the supernatant was used for biochemical studies.
Estimation of carbohydrates by Anthrone’s method (Hedge and Hofreiter, 1962), proteins by Lowry et al. (1951), lipids by AACC (1999), TIU by Kakade et al. (1969), phenols by FCR method, tannins by Follin-Dennis spectrophotometric method (Pearson, 1976) have been undertaken in the present study.
Statistical analysis: All experiments were performed in triplicates. Analysis at every time point from each experiment was carried out in triplicate. Means, standard errors and standard deviations were calculated from replicates within the experiments and analyzed using Microsoft Excel.
RESULTS AND DISCUSSION
All the genotypes of three legumes used in present study showed differential susceptibility against both the bruchid species. This relative resistance was due to the seed physical factors and biochemical factors such as nutritional and antinutritional factors. According to the recorded data it can be summarized that nutritional composition was not the cause of relative resistance or susceptibility to bruchid in legumes but antinutritional composition of legumes found responsible for relative resistance in them against bruchid species.
The nutritional components viz., carbohydrates, proteins and fat (mg g-1) varied significantly among all the genotypes of cowpea, chickpea and soybean used in the present study. The protein content in cowpea genotypes varied from 200.03±2.41 to 270.92±3.89 mg g-1. The content value of carbohydrates varied from 535.85±5.99 to 581.4±1.02 mg g-1. The amount mg g-1 fat varied significantly from 10.42 to 13.20 mg g-1 (Table 1). The composition of nutritive contents recorded in present study for different cowpea genotypes is in close agreement with the studies of Tindal (1992) who reported 22-26% proteins, 1-2% fats and oil and 60-65% carbohydrates in cowpea and Hedley (2001) also reported the protein content in the range of 22.90-23.65 and 1.2% fat contents in cowpea genotypes.
The protein contents values of chickpea genotypes varied significantly between 182.59±4.93 to 261.27±4.48 mg g-1 seeds.
| Table 1: | Biochemical composition of cowpea genotypes |
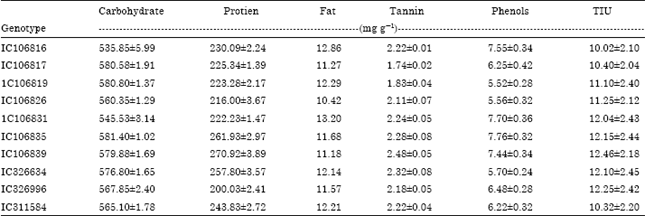 | |
| Reported values are the Mean±SD (n = 3), TIU: Trypsin inhibitor units | |
| Table 2: | Biochemical composition of chickpea genotypes |
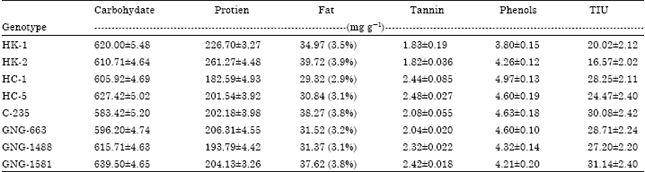 | |
| Reported values are the Mean±SD, (n = 3), TIU: Trypsin inhibitor units | |
| Table 3: | Biochemical composition of soybean genotypes |
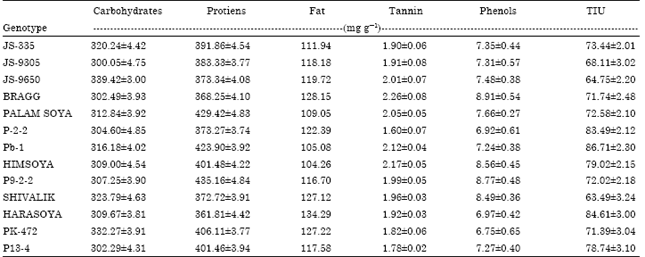 | |
| Reported values are the Mean±SD, (n = 3), TIU: Trypsin inhibitor units | |
Similar trend of protein concentration was also recorded by Sharma et al. (2013) where the values ranged between 18.0±1.52 to 23.0±0.57% in desi genotypes and 28±1.00 to 31±0.57% in kabuli genotypes. Similarly, Hedley (2001) reported 15.5-28.2% protein contents and 3.1-7% fat contents among chickpea genotypes. Carbohydrate content varied between 583.42±5.20 to 639.50±4.65 mg g-1 and total fat content values ranged from 29.32 to 39.72 mg g-1 (Table 2). The value of protein in soybean genotypes varied from 321.86±2.54 to 465.16±2.84 mg g-1. The crude carbohydrate of soybean genotypes ranged between 300.05±4.72 to 339.42±3.00 mg g-1 and fat content of seed for soybean genotypes ranged between 111.94 to 134.29 mg g-1 seed (Table 3). The nutritional contents of soybean genotyeps reported in this study are consistant with the findings of Azlan et al. (2011) who reported 37.37% carbohydrates and Wilson (2004) reported the proteins between 34-56.80% and oil content between 8.1-27.9% for soybean genotypes. Similarly, the values of antinutritional components for different genotypes recorded were different as for cowpea genotypes the value of tannin ranged between 1.74±0.02 to 2.48±0.05 mg g-1. Similar values of tannin contents were recorded by Afiukwa et al. (2012) in 104 genotypes of cowpea. phenols ranged between 5.52±.28 to 7.76±0.32 mg g-1 and TIU ranged between 10.12±2.10 to 12.46±2.18 mg g-1 (Table 1), the TIU units recorded for cowpea in the present study are closely related to 4.6 to13.3 mg g-1 TIU content in seeds of Vigna unguiculata reported by Kochhar et al. (1988), for chickpea the value of tannin recorded ranged between 1.82±0.36 to 2.48±0.027 mg g-1. Similar concentration of tannin contents ranged between 0.07-0.13% in kabuli and 0.18-0.22% in desi chickpea genotypes have been reported by Sharma et al. (2013). Phenol values recorded ranged between 3.80±0.15 to 4.97±0.13 mg g-1 and TIU values were 16.57±2.0 to 31.14±2.40 mg g-1 (Table 2). Results of present study found in agreement with Kansal et al. (2008) who reported trypsin inhibitor contents of 8.71 to 39.47 mg g-1 in ten desi chickpea cultivars and for soybean the tannin values between 1.60±0.07 to 2.26±0.08 mg g-1, phenol values between 6.75±0.65 to 8.91±0.54 mg g-1 and TIU values were ranged between 64.75±2.20 to 86.71±2.30 mg g-1 (Table 3). Similar findings of trypsin inhibitor activity were recorded by Guillamon et al. (2008) who reported the 43-83 TIU mg g-1 among soybean genotypes.
According to the recorded data it can be summarized that nutritional composition is not the cause of relative resistance or susceptibility to bruchid in legumes but antinutritional composition of legumes is responsible for relative resistance in them against bruchid species. All the genotypes of chickpea show relative behavior to both the bruchid species, but the genotypes HK-I and HK-2, HC-5 and GNG-1488 with less TIU units, phenols and Tannin had high adult emergence, growth index, weight loss and less developmental to both the species as compared to partial susceptible GNG-663, HC-I genotypes and resistant C-235 and GNG-1581 genotypes which showed less adult emergence, growth index, wt. loss and more developmental period because of the presence of high antinutritional factors (Tannin, Phenols and TIU) which affect their metabolic activity and inhibit their growth.
All the soybean genotypes showed relative resistance to both the species as compared to cowpea and chickpea which were recorded highly susceptible to both the species. Among soybean genotypes JS-335, JS-9305, JS-9560, PK-472, P9-2-2, him soya and P13-4 were partially susceptible to both the species, as had high adult emergence, growth index, wt. loss as compared to highly resistance genotypes (Hara Soya, Shivalik, Bragg and Pb-I).
The high resistance behaviour of soybean genotypes to both the species might be because of the presence of high amount of phenols and TIU, which is known to inhibit the action of trypsin enzyme. It does not necessarily interfere with the digestion of protein, but may retard the liberation of methionine amino acid from the protein (Richardson, 1977), thus methionine cannot be used in protein synthesis. Phenolic compounds are responsible to form complex with food nutrients such as minerals and proteins, thus rendering them less soluble or less susceptible to enzymatic degradation and less available for absorption (Towo et al., 2003). Despite the antinutritional activity of phenols they play a significant role by reducing oxidative damage because of their ability to act as antioxidents, presence of these anti-nutritional factor in high amount inhibit the growth of larvae by blocking and disturbing the metabolic pathways. In comparison to all the genotypes of three legumes, soybean genotypes with less amount of carbohydrates, high amount of proteins, fats and antinutrional components (phenols and TIU) were highly resistant to both the species of Callosobruchus than chickpea and cowpea genotypes. Although, soybeans are important source of proteins and oils but their acceptability and utilization by both the bruchid species has been limited due to the presence of high level of phenols and TIU as compared to cowpea and chickpea, which provided defensive mode to soybean against C. maculatus and C. chinensis. The present study is in close agreement with the findings of Moraes et al. (2000) and Ignacimuthu et al. (2000) who reported that presence of antinutritional factors in seeds protect them against different pests and Srinivasan and Durairaj (2007) reported 3-5 times higher TIU in the resistant accessions of V. umbellata than susceptible accessions. Trypsin inhibitors are wide spread antinutritioanl substances which blocks the trypsin activity and therefore, reduce digestion of proteins. The T1 contents varied signifincatly among desi and kabuli chick pea genotyeps. In present study desi chick pea genotypes were resistant to both the bruchid species probably due to the presence of high antinutritional factors like tannin, phenols and TIUs as compared to kabuli chickpeas. These antinutritional factors resist the survival of larvae in them by blocking the utilization of nutrients through astringency and enzymes inhibition. The findings of Khattab and Arntfield (2009) are in the close agreement with the present results, who reported that tannins are secondary metabolities which inhibit the digestive enzymes and therefore, lower the digestibility of important nutrients especially proteins and starch. Similarly, Diaz et al. (2010) reported that anti nutritional factor tannins are polymeric flavinoids that comprise a small part of the broad and diverse group of phenolic compounds produced by plants as secondary metabolities.
REFERENCES
- Adjadi, O., B.B. Singh and S.R. Singh, 1985. Inheritance of bruchid resistance in cowpea. Crop Sci., 25: 740-742.
CrossRefDirect Link - Afiukwa, C.A., O. Onwuchekwa, U.A. Ibiam, C.O. Edeogu and P.M. Aja, 2012. Characterization of cowpea cultivars for variations in seed contents of some Antinutritional Factors (ANFs). Cont. J. Food Sci. Technol., 6: 25-34.
Direct Link - Duranti, M. and C. Gius, 1997. Legume seeds: Protein content and nutritional value. Field Crops Res., 53: 31-45.
CrossRefDirect Link - Diaz, A.M., G.V. Caldas and M.W. Blair, 2010. Concentrations of condensed tannins and anthocyanins in common bean seed coats. Food Res. Int., 43: 595-601.
CrossRefDirect Link - Guillamon, E., M.M. Pedrosa, C. Burbano, C. Cuadrado, M.D.C. Sanchez and M. Muzquiz, 2008. The trypsin inhibitors present in seed of different grain legume species and cultivar. Food Chem., 107: 68-74.
CrossRefDirect Link - Ignacimuthu, S., S. Janarthanan and B. Balachandran, 2000. Chemical basis of resistance in pulses to Callosobruchus maculatus (F.) (Coleoptera: Bruchidae). J. Stored Prod. Res., 36: 89-99.
CrossRefDirect Link - Kakade, M.L., N. Simons and I.E. Liener, 1969. An evaluation of natural vs. synthetic substrates for measuring the antitryptic activity of soybean samples. Cereal Chem., 46: 518-526.
Direct Link - Kochhar, N., A.F. Walker and D.L. Pike, 1988. Effect of variety on protein content, amino acid composition and trypsin inhibitor activity of cowpeas. Food Chem., 29: 65-78.
CrossRef - Kansal, R., M. Kumar, K. Kuhar, R.N. Gupta, B. Subrahmanyam, K.R. Koundal and V.K. Gupta, 2008. Purification and characterization of trypsin inhibitor from Cicer arietinum L. and its efficacy against Helicoverpa armigera. Braz. J. Plant Physiol., 20: 313-322.
CrossRef - Khattab, R.Y. and S.D. Arntfield, 2009. Nutritional quality of legume seeds as affected by some physical treatments 2. Antinutritional factors. LWT-Food Sci. Technol., 42: 1113-1118.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Moraes, R.A., M.P. Sales, M.S.P. Pinto, L.B. Silva and A.E.A. Oliveira et al., 2000. Lima bean (Phaseolus lunatus) seed coat phaseolin is detrimental to the cowpea weevil (Callosobruchus maculates). Braz. J. Med. Biol. Res., 33: 191-198.
PubMed - Azlan, A., Z. Amom, M.R.M. Rusydi and C.W. Noraliza, 2011. Nutritional changes in germinated legumes and rice varieties. Int. Food Res. J., 18: 705-713.
Direct Link - Richardson, M., 1977. The proteinase inhibitors of plants and micro-organisms. Phytochemistry, 16: 159-169.
CrossRefDirect Link - Srinivasan, T. and C. Durairaj, 2007. Biochemical basis of resistance in rice bean, Vigna umbellata Thunb. (Ohwi and Ohashi) against Callosobruchus maculatus F. J. Entomol., 4: 371-378.
CrossRefDirect Link - Towo, E.E., U. Svanberg and G.D. Ndossi, 2003. Effect of grain pre-treatment on different extractable phenolic groups in cereals and legumes commonly consumed in Tanzania. J. Sci. Food Agric., 83: 980-986.
CrossRefDirect Link