
Ema Komalasari
Master Program of Crop Science, Faculty of Agriculture, Universitas Padjadjaran, 45363 Jatinangor, Indonesia
Fitri Widiantini
Faculty of Agriculture, Universitas Padjadjaran, 45363 Jatinangor, Indonesia
Santika Sari
Faculty of Agriculture, Universitas Padjadjaran, 45363 Jatinangor, Indonesia
Nono Carsono
Faculty of Agriculture, Universitas Padjadjaran, 45363 Jatinangor, Indonesia
LiveDNA: 62.31343
Asian Journal of Crop Science
Year: 2020 | Volume: 12 | Issue: 2 | Page No.: 124-129







ABSTRACT
Background and Objective: Rice tungro disease remains as one of the major threat to rice production in Indonesia and other countries. One of the proven methods in preventing the occurrence of tungro virus is through the use of resistant varieties. The development of resistance of glutinous rice (Oryza sativa glutinosa) to tungro virus is expected to be achieved through hybridization between glutinous rice with tungro resistance rice. The aim of this study was to estimate number of gene controlling tungro resistance and obtain rice progenies that resistant to the virus. Materials and Methods: Around 230 F2 progenies of Ketonggo×Utri Merah as well as Ketonggo× ARC12596 were evaluated. Forced-tube inoculation method was used to inoculate tungro disease and standard evaluation system for rice from IRRI was applied to assess the tungro infection symptoms. Scoring was performed at 2, 4 and 10 weeks after inoculation (wai). Results: The distinguished progenies with recovery abilities and stable have been obtained. The inheritance of tungro resistance was supposed to be controlled by recessive genes with different patterns, for Ketonggo×Utri Merah was susceptible 3: 1 resistant (complete dominance) and susceptible 9: 7 resistant (duplicate recessive epistasis) for Ketonggo×ARC12596. Conclusion: Recessive genes are supposed to control the tungro virus resistance but the inheritance pattern on both crosses is slightly different, indicating donor parents have dissimilar resistant genes.
PDF Abstract XML References Citation
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Ema Komalasari, Fitri Widiantini, Santika Sari and Nono Carsono, 2020. Segregation Ratios of Tungro Virus Resistance on F2 Progenies Derived from Utri Merah and ARC12596. Asian Journal of Crop Science, 12: 124-129.
DOI: 10.3923/ajcs.2020.124.129
URL: https://scialert.net/abstract/?doi=ajcs.2020.124.129
DOI: 10.3923/ajcs.2020.124.129
URL: https://scialert.net/abstract/?doi=ajcs.2020.124.129
INTRODUCTION
Rice tungro disease (RTD) is one of the factors that inhibit rice productivity in Indonesia and other countries. In 2011, the Directorate of Food Crop Protection, Ministry of Agriculture, Indonesia reported that 13,868 ha of rice fields in Indonesia have been attacked by tungro1. Data from the Ministry of Agriculture of Indonesia2 showed that tungro is still an important disease in rice with an extensive attack in 2015, 2016 and 2017 which were reaching an area of 4.803, 3.518 and 5.159 ha, respectively. The disease is caused by infection with 2 viruses, which are the Rice Tungro Bacilliform Virus (RTBV) and the Rice Tungro Spherical Virus (RTSV). Both viruses are effectively transmitted by semi-persistent green leafhopper (GLH) Nephotettix viruscens (Distant)3.
Control of tungro virus can be done by managing the crop environment. Jefferson and Chancellor 4 stated that the eradication of virus sources and reducing vector population density in the field can be reached through the disease management system. The use of resistant varieties to tungro is efficient controlling method to be applied by the farmer both in the technical irrigated rice fields and in the rainfed lowland rice fields5. In Indonesia, tungro-resistant varieties are used to suppress the spread of disease. Improving the resistance of varieties to tungro disease is done through plant breeding program using tungro resistant genetic sources5. However those rice lines have not been released yet and resistant rice lines produced are Oryza sativa rice, not Oryza sativa var. glutinousa, hence, the tungro-resistant glutinous rice has not been obtained so far. It is therefore that the aim of the research was to evaluate segregation ratios of tango virus resistance in F2 progenies at the 2 crosses and obtain tungro resistant genotypes from 2 sources of resistant varieties, i.e., Utri Merah and ARC12596. Thus it is expected that we obtain the tungro-resistant glutinous rice genotypes that would suppress the spread of tungro.
MATERIALS AND METHODS
The experiment was conducted from February-May, 2018 in the Indonesian Center for Rice Research (ICRR) greenhouse and experimental station of the Faculty of Agriculture, Padjadjaran University. F2 progenies were planted as many as 230 genotypes for both crosses: Ketonggo×Utri Merah and Ketonggo×ARC12596. Five check varieties (3 parents, one resistant variety and one susceptible variety) were grown also. Artificial hybridization was carried out in 2014 at the Indonesia Tungro Disease Research Station, Lanrang, south Sulawesi. Tungro isolates were obtained from Subang, West Java in 2017, which maintained in a greenhouse. Inoculation was done according to Cabauatan et al.6. Observation of visual symptoms was conducted 2 and 4 weeks after inoculation (wai hereafter) and in the field at 10 wai. Observations were made based on the standard evaluation system (SES) for 1-9 scale rice7. Test of segregation ratios was performed by Chi-square.
RESULTS
Evaluation of symptoms: The F2 progenies obtained from the 2 crosses were evaluated for tungro resistance at the age of 7 days after germination. The progenies of F2 derived from the Ketonggo×Utri Merah were classified as susceptible due to the appearance of their symptoms i.e., discoloration leaves from yellow to orange and stunted (Fig. 1a,b). The similar appearance was also found on the progenies of Ketonggo× ARC12596 (Fig. 1c,d). Meanwhile, another symptom of the tungro infection was also found, such as rolled out of leave, like a spiral. They were categorized as susceptible (Fig. 2a-c).
Progenies from both crosses showed various symptoms at different time of the observation (Fig. 3). In the second and 4th wai as many as 181 F2 progenies of the Ketonggo×Utri Merah did not show any symptoms of tungro virus infection but at the 10 wai, there were only 49 progenies that did not show any symptoms of tungro infection. From Ketonggo× ARC12596, 126 progenies did not show symptoms in the second wai but the number of the progenies without symptoms decreased in the 4th and 10 wai i.e., 115 and 31 progenies, respectively (Fig. 3).
In particular genotypes from both crosses, symptoms were seen at 2 and 4 wai but those symptoms disappeared at the end of the vegetative phase. Eight progenies from Ketonggo×Utri Merah and 12 progenies from Ketonggo× ARC12596 which show resistance reaction at the 10 wai were fruitfully obtained (Table 1). These genotypes had recovery abilities and did not show symptoms of discoloration of leaves until the end of the vegetative phase.
Segregation pattern analysis: The Chi square test was performed to estimate the number of genes controlling the tungro resistance. The grouping was made based on the scores of tungro symptoms as seen in the field. The Mendelian ratio and this modification were classified into groups of classes (2, 3 and 4 classes, Table 2 and 3).
 |
| Fig. 1(a-d): | Comparison of tungro virus resistance score in both crosses (a, b) Ketonggo×Utri Merah and (c, d) Ketonggo×ARC12596 |
 |
| Fig. 2(a-c): | Symptoms of tungro infected plants, (a) Rice genotypes expressing typical tungro symptoms, (b) Twisted leaf symptoms and (c) Leaf discoloration from yellow to orange |
| Table 1: | Score recovery genotypes from Ketonggo×Utri Merah and Ketonggo×ARC12596 at 2, 4 and 10 wai |
| Ketonggo×Utri Merah | Ketonggo×ARC12596 | ||||||
| Genotypes | 2 wai | 4 wai | 10 wai | Genotypes | 2 wai | 4 wai | 10 wai |
| C.1.6.3 | 7 | 7 | 1 | D.1.6.5 | 9 | 9 | 1 |
| C.2.3.2 | 7 | 7 | 3 | D.1.7.9 | 7 | 7 | 3 |
| C.3.1.4 | 9 | 7 | 3 | D.2.2.10 | 7 | 7 | 3 |
| C.3.7.5 | 9 | 9 | 3 | D.2.3.10 | 7 | 7 | 1 |
| C.4.1.10 | 1 | 9 | 3 | D.2.6.1 | 1 | 7 | 3 |
| C.4.3.6 | 1 | 7 | 3 | D.3.4.3 | 7 | 7 | 3 |
| C.4.4.6 | 9 | 1 | 3 | D.3.4.4 | 1 | 7 | 1 |
| C.4.4.9 | 9 | 9 | 1 | D.3.6.6 | 7 | 7 | 1 |
D.3.7.8 | 1 | 7 | 1 | ||||
D.4.1.2 | 7 | 7 | 1 | ||||
D.4.2.7 | 7 | 7 | 3 | ||||
D.4.5.10 | 7 | 1 | 1 | ||||
| wai: Week after inoculation | |||||||
| Table 2: | Inheritance of resistance to tungro disease in F2 Ketonggo×Utri Merah | ||
| Mendelian ratio and modification | Observed | χ2 | p-value |
| Two grouping | |||
| 3:01 | 159:71 | 3.92ns | 0.025-0.050 |
| 9:07 | 159:71 | 14.99* | <0.001 |
| 13:03 | 159:71 | 21.39* | <0.001 |
| 15:01 | 159:71 | 233.74* | <0.001 |
| Three grouping | |||
| 1:02:01 | 123:36:71 | 1045.71* | <0.001 |
| 9:03:04 | 123:36:71 | 4.66 ns | 0.050-0.100 |
| 9:06:01 | 123:36:71 | 252.64* | <0.001 |
| 12:03:01 | 123:36:71 | 238.43* | <0.001 |
| Four grouping | |||
| 9:3:3:1 | 49:58:47:76 | 269.66* | <0.001 |
| 6:3:3:4 | 49:58:47:76 | 7.48* | <0.001 |
| ns: Not significant at α = 1%, *Significant at α = 1% | |||
| Table 3: | Inheritance of resistance to tungro disease in the F2 Ketonggo×ARC12596 Mendelian ratio | ||
| Mendelian ratio and modification | Observed | χ2 | p-value |
| Two grouping | |||
| 3:01 | 146:84 | 15.68* | <0.001 |
| 9:07 | 146:84 | 4.59 ns | 0.025-0.050 |
| 13:03 | 146:84 | 46.52* | <0.001 |
| 15:01 | 146:84 | 354.56* | <0.001 |
| Three grouping | |||
| 1:02:01 | 84:89:57 | 220.60* | <0.001 |
| 9:03:04 | 84:89:57 | 64.72* | <0.001 |
| 9:06:01 | 84:89:57 | 142.39* | <0.001 |
| 12:03:01 | 84:89:57 | 220.60* | <0.001 |
| Four grouping | |||
| 9:3:3:1 | 31:142:49:8 | 52.43* | <0.001 |
| 6:3:3:4 | 31:142:49:8 | 270.11* | <0.001 |
| ns: Not significant at α = 1%, *Significant at α = 1% | |||
Chi-square test showed 2 segregation ratios that fit the Mendelian ratio i.e., 3:1 for Ketonggo×Utri Merah (Table 2) and susceptible 9:7 resistant (duplicate recessive epistasis) for Ketonggo×ARC12596 (Table 3).
DISCUSSION
Estimating number of genes controlling the resistance to tungro virus from 2 resistant parents is greatly challenging and the effort is also important in order to determine suitable selection method that will be applied for next successive generation. The gene controlling the trait in rice is not known yet so far. Therefore we are interested in exploring the resistant genotypes i.e., Utri Merah and ARC12596 as donor parents through phenotypic assessment of tungro virus infection.
Chancellor et al.8 stated that the initial symptom of virus infection in susceptible rice found that rice plants are very stunted, low number of tillers and changes in leaf color, from yellow to orange.
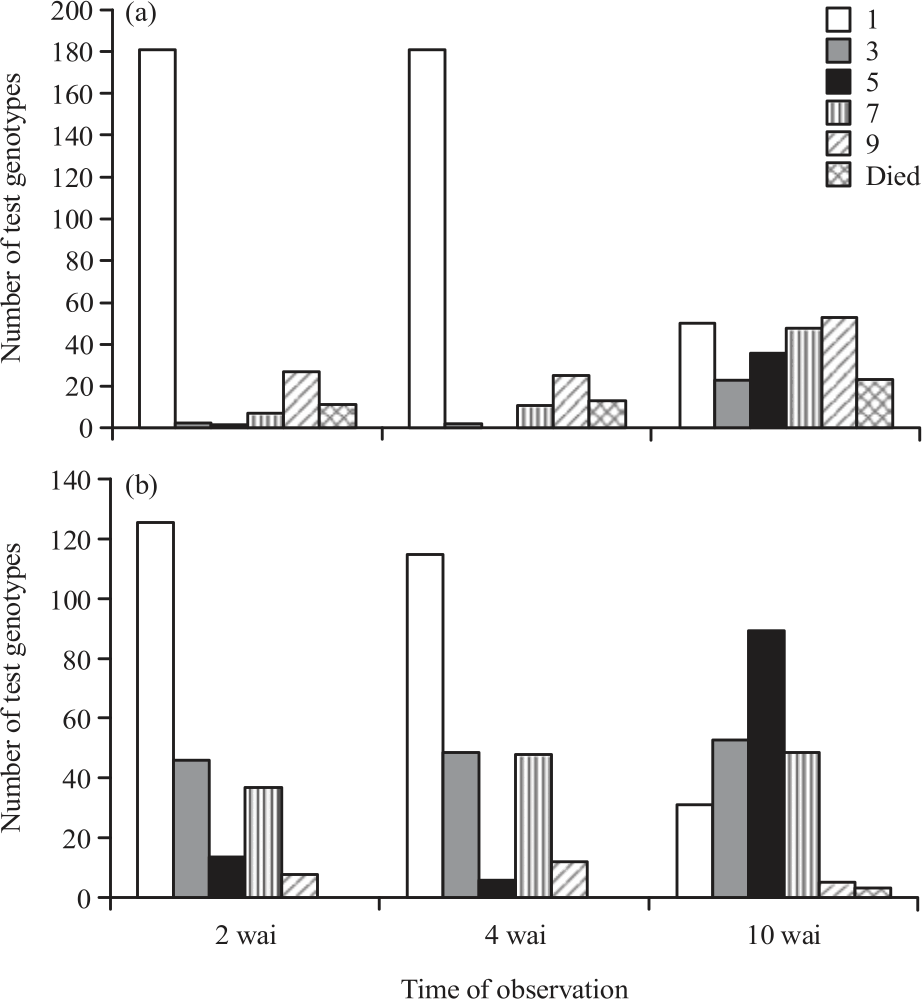 |
| Fig. 3(a-b): | Changes in the number of test genotype shown tungro symptoms at different time of observation, (a) Ketonggo×Utri Merah and (b) Ketonggo×ARC12596 |
In addition, Hibino et al.3, rice plant infected RTBV dan RTSV showed stunted and yellowing leaf symptoms. Tungro symptoms in rice plants, according to Azzam and Chancellor4, were very dependent on the age of the plant, the rice variety and the strain of the virus. The leaves of rice plants infected with the tungro virus Ling9 found to have phenotypic appearance such as, slim and rolled out and spiral.
Tungro infection leads to chlorophyll reduction in a plant, therefore leaf colour changes to yellow. Jabeen et al.10 reported that reduction in chlorophyll A and B content was high in susceptible plant, followed by moderate and the resistant cultivars that showed less reduction. However the colour changes can also be caused by the inhibited water transportation process, nutrition and photosynthesis product. Cruz et al.11 found that RTBV occurred both in the xylem and phloem, whereas RTSV directly to the xylem where the virus multiplies. Phloem and xylem damage affect the inhibition of plant growth may cause rice plant stunting and tiller number decrease.
The study demonstrated the presence of consistent resistant genotypes at 2, 4 and 10 wai. The results of this research showed the genotypes that having the similarity of resistant reaction from 2-10 wai were stable resistant genotypes. From the crossing of Ketonggo×Utri Merah, 47 stable resistant genotypes were obtained, while the Ketonggo×ARC12596 was 32 genotypes.
Genotypes that had recovery ability are potentially be used to control the spreading of tungro disease. Khatun et al.12 argued that the most effective method for controlling tungro disease was by using resistant varieties or varieties that have recovery ability when it compared to the method of destroying vectors using insecticide. However, Khatun et al.12 pointed out that susceptible varieties which did not have recovery ability would be much suffering from the rice tungro infection and had a potential of 100% yield losses.
Chi-square test showed 2 segregation ratios that fit the Mendelian ratio i.e., 3:1 (Ketonggo×Utri Merah). Segregation pattern with a ratio of 3:1 has a smaller p-value 0.025. (Table 2). This implies that resistance to tungro in Utri Merah (acc. #16680) is controlled by one pair gene with complete dominance (3 susceptible: 1 resistant). This finding is different with Azzam et al.13, who found segregation ratio 9:7.
A segregation ratio of 9 susceptible: 7 resistant with p-value 0.025-0.05 (Table 3) fitted for Ketonggo×ARC12596. This ratio indicates that the resistance gene in ARC12596 is controlled by 2 pairs of recessive genes or duplicate recessive epistasis. It implies a recessive allele at one locus and a recessive at other interact to produce expressed phenotype. Griffiths et al.14 defined complementary epistasis as a gene interaction where the gene functions would be needed by other genes in metabolism. Mendelian ratio from both crosses, showed that each resistant parent has a different genetic composition. Based on our data, it is assumed that genetic composition of rice donor plant plays significant role in the response to the virus infection.
CONCLUSION
Tungro virus resistance is controlled by simple genic, one and 2 pairs of recessive genes are supposed to control the trait.
Two recessive genes are supposed to be controlling the tungro virus resistance but the inheritance pattern on both crosses is slightly different, indicating donor parents have dissimilar resistant genes. The resistant genotypes are highly valuable for developing resistant glutinous rice to tungro virus.
The resistant genotypes are highly valuable for developing resistant glutinous rice to tungro virus.
SIGNIFICANCE STATEMENT
The study discovered F2 progenies of glutinous rice lines that resistant to tungro virus with different symptoms and inheritance patterns thus will open the possibility to develop more resistant rice lines and to release new resistant variety for Oryza sativa glutinousa. This study will help researchers, most particularly plant breeders to uncover the critical area of inheritance patterns of tungro resistance that many researchers were not able to explore. Thus a new approach on breeding Oryza sativa glutinousa with tungro resistance may be arrived at.
ACKNOWLEDGMENT
We wish to express our sincere gratitude to Dr. Ahmad Muliadi, Dr. Rahmini and Dr. Suprihanto, for support and the staffs at the plant protection research group ICRR.
REFERENCES
- Budiyanto, E., M. Nurhidayat, Suparni and S. Haryati, 2011. Plant Protection to Suppress Yield Loss of Rice. In: National Seminar on Tungro Disease: National Seminar on Tungro Disease. Technology Innovation in Tungro Disease Control and Main Rice Pests Towards Sustainable Self-Sufficiency, Hermanto, A. Muis and S. Pakki (Eds.)., Indonesia Center for Food Crops Research and Development, Makassar, ISBN:978-979-11549-53-1, pp: 1-9.
- Hibino, H., M. Roechan and S. Sudarisman, 1978. Association of two types of virus particles with penyakit habang (tungro disease) of rice in Indonesia. Phytopathology, 68: 1412-1416.
Direct Link - Jefferson, O.A. and T. Chancellor, 2002. The biology, epidemiology and management of rice tungro disease in Asia. Plant Dis. Int. J. Applied Plant Pathol., 86: 88-100.
Direct Link - Hasanuddin, A., 2008. Improve the resistance of rice varieties to tungro disease. Iptek Tanaman Pangan, 3: 215-228.
Direct Link - Cabauatan, P.Q., R.C. Cabunagan and H. Koganezawa, 1995. Biological variants of rice tungro viruses in the Philippines. Phytopathology, 85: 77-81.
Direct Link - IRRI., 2013. Standard Evaluation System for Rice. 5th Edn., International Rice Research Institute, Philippine, Pages: 55.
Direct Link - Chancellor, T.C.B., J. Holt, S. Villareal, E.R. Tiongco and J. Venn, 2006. Spread of plant virus disease to new plantings: A case study of rice tungro disease. Adv. Virus Res., 66: 1-29.
CrossRefPubMedDirect Link - Jabeen, A., T.V. Kiran, D. Subrahmanyam, D. Lakshmi, G. Bhagyanarayana and D. Krishnaveni, 2017. Variations in chlorophyll and carotenoid contents in Tungro infected rice plants. J. Res. Dev., Vol. 5, No. 1.
CrossRefDirect Link - Cruz, F.C.S., H. Koganezawa and H. Hibino, 1993. Comparative cytology of rice tungro viruses in selected rice cultivars. J. Phytopathol., 138: 274-282.
CrossRefDirect Link - Khatun, M.T., M.A. Latif, M.M. Rahman, M. Hossain and T.H. Ansari et al., 2017. Recovering ability of upland and rainfed lowland rice varieties against rice tungro disease. Bangladesh Rice J., 21: 91-100.
CrossRefDirect Link - Azzam, O., T. Imbe, R. Ikeda, P.D. Nath and E. Coloquio, 2001. Inheritance of resistance to rice tungro spherical virus in a near-isogenic line derived from Utri Merah and in rice cultivar TKM6. Euphytica, 122: 91-97.
CrossRefDirect Link