
Joshua Kiilu Muli
Department of Botany, Jomo Kenyatta University of Agriculture and Technology, Nairobi, Kenya
Cecilia Mweu
Institute for Biotechnology Research, Jomo Kenyatta University of Agriculture and Technology, Nairobi, Kenya
Nancy Budambula
Department of Biological sciences, University of Embu, Embu, Kenya
Sylvester Elikana Anami
Institute for Biotechnology Research, Jomo Kenyatta University of Agriculture and Technology, Nairobi, Kenya
LiveDNA: 254.19816
Asian Journal of Crop Science
Year: 2017 | Volume: 9 | Issue: 3 | Page No.: 71-81







ABSTRACT
Background and Objective: Tropical maize is the most cultivated crop in sub-Saharan Africa and is a staple food to over 220 million people. This study, evaluated the competence of four tropical maize (Zea mays L.) inbred lines to callus induction and regeneration through somatic embryogenesis when co-cultivated on yeast extract peptone medium (YEP). Materials and Methods: Transient GUS assay was used to evaluate the competence of the genotypes to Agrobacterium-mediated transformation using YEP as co-cultivation media or when YEP was supplemented with 2,4-D (YEP+2,4-D), cysteine (YEP+CYS), proline (YEP+PRO) or in combination (YEP+ALL). Data on all parameters were analyzed using multivariate ANOVA and SAS. Results: Co-cultivation media based on YEP alone did not impact callus induction and immature embryos exhibited preconscious germination. When YEP was supplemented with 2,4-D at concentrations 1.5 and 3 mg L–1 2,4-D, the formation of embryogenic calli was induced and regeneration initiated. Immature embryos had high transient GUS expression when co-cultivated with Agrobacterium on YEP, YEP+PRO and YEP+ALL media than when co-cultivated on MS media, suggesting that YEP promotes Agrobacterium-mediated integration of transgenes in tropical maize. Agrobacterium tumefaciens at a concentration of 0.07 (OD660) gave the highest transient GUS expression (20.90%) while concentrations of 0.8 and 0.2 resulted in low transient GUS expression (9.17 and 12.22%, respectively). Conclusion: The integration of YEP media in the Agrobacterium-mediated transformation protocols is likely to contribute in the development of a more efficient Agrobacterium-mediated transformation system for tropical maize genotypes.
PDF Abstract XML References Citation
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Joshua Kiilu Muli, Cecilia Mweu, Nancy Budambula and Sylvester Elikana Anami, 2017. Yeast Extract Peptone Based Co-cultivation Media Promotes Transient GUS Expression in Tropical Maize Genotypes. Asian Journal of Crop Science, 9: 71-81.
DOI: 10.3923/ajcs.2017.71.81
URL: https://scialert.net/abstract/?doi=ajcs.2017.71.81
DOI: 10.3923/ajcs.2017.71.81
URL: https://scialert.net/abstract/?doi=ajcs.2017.71.81
INTRODUCTION
Tropical maize plays a great role in the diet of more than 400 million people in the world and is the most preferred crop in Southern and Eastern Africa. It remains the most cultivated crop in Africa and contributes significantly to food and feed security1. The constraints to maize production include both biotic and abiotic factors. It is, therefore, necessary to develop varieties which are properly adapted to the stresses in question in order to increase maize yield in the region. Conventional breeding has for a long time played a role towards achieving this, however, better improved and precise techniques have emerged from biotechnology since the discovery of protoplasm transformation2, particle bombardment3 and Agrobacterium-mediated transformation4. All these techniques however depend on stable and high efficient uptake of genes and the ability to regenerate the transformed explants to whole plants which can express the cloned genes to achieve the desired results5. Although Agrobacterium-mediated transformation promises stable integration with a low copy number of the transgene as well as highly fertile transgenic plants, the reported transformation frequencies which range from 1.4-8% are not good enough6–8.
Recalcitrance of tropical maize genotypes to Agrobacterium-mediated transformation can be attributed to many factors acting either individually or in combination during the transformation process. For instance, it has been demonstrated that maize immature embryos produce substances that inhibit the growth of Agrobacterium around it thus affecting the chemotactic attraction of Agrobacterium9. Chu media, Linsmaier and Skoog (LS) media and Murashige and Skoog (MS) media have been optimized in order to enhance regeneration of plants and T-DNA transfer and integration into plant cells10,11. In sugarcane, the type of media and period of co-cultivation affects the frequency and efficiency of transformation12. Ombori et al.13 also showed that the type and media components affect transformation efficiency in immature maize embryos. Competency of tropical maize immature embryos to Agrobacterium-mediated transformation using GUS as a reporter gene and MS media has been reported before14. In this study, the competence of immature maize embryos to Agrobacterium-mediated transformation based on transient β-glucuronidase (GUS) expression using yeast extract peptone (YEP) as the co-cultivation media was sought, as well as assessing the impact of this media on the regeneration efficiency of immature maize embryos.
MATERIALS AND METHODS
This study was carried out between October, 2015 and November, 2016.
Plant materials and Agrobacterium strain: Embryos for all experiments were obtained from seeds of inbred lines CML 144, E04, A04 and CML 216 provided by the Kenyatta University Plant Transformation Laboratory from where the research was also done. The seeds were planted in plots in the research fields of the Jomo Kenyatta University of Agriculture and Technology. Ears were harvested at 14 days after pollination (DAP) and used either immediately or stored at 4°C for a maximum of 3 days as described by Frame et al.10. Whole ears were surface sterilised in 70% commercial grade ethanol for 3 min and 5% sodium hypochlorite for 20 min after which they were rinsed 3 times in sterile distilled water10. Agrobacterium tumefaciens strain EHA 101 with the standard binary vector pTF 102 was used to infect the embryos of the different maize lines. The T-DNA region of this construct consists of a spectinomycin-resistance marker gene for bacteria selection and a CaMV 35S promoter to drive the GUS reporter gene10. The bacteria strain containing the vector was maintained on YEP media containing 100 mg L–1 spectinomycin and 100 mg L–1 kanamycin. Bacteria cultures for weekly experiments were drawn from the mother plate refreshed from long term stored glycerol stocks. Before every experiment, bacteria cells were grown overnight in liquid YEP media and adjusted to an optical density (OD660) 0.8. This was designated the 100% Agrobacterium concentration which was serial diluted to 75, 50 and 25%, corresponding to 0.2, 0.03 and 0.007 OD’s, respectively.
Culture media and modifications: The infection media (IM) used was described by Frame et al.10 with a slight modification on the amount of Acetosyringone (AS) to 300 mM. Co-cultivation of infected embryos was performed on two types of media (Table 1). The first co-cultivation media (CCM) consisted of 2.5 g L–1 yeast extract, 5 g L–1 peptone, 2.5 g L–1 NaCl, 220 mg L–1 casein hydrolysate, 20 g L–1 sucrose, 10 g L–1 glucose, 8 g L–1 phytoagar and pH adjusted to 5.6. After autoclaving, 16 mg L–1 AgNO3, 300 mM AS, 100 mg L–1 kanamycin and 100 mg L–1 spectinomycin were added. The effects of adding 2, 4-D, proline and cysteine on transient GUS expression in this media were also analyzed. The second co-cultivation media consisted of MS macro and micro salts and vitamins, 1.5 mg L–1 2, 4-D, 20 g L–1 sucrose, 10 g L–1 glucose, 0.7 g L–1 proline, 220 mg L–1 casein hydrolysate 8 g L–1 phytoagar and pH adjusted to 5.6. After autoclaving, 16 mg L–1 AgNO3, 300 mg L–1 cysteine, 300 mM AS, 100 mg L–1 kanamycin and 100 mg L–1 spectinomycin were added.
| Table 1: | Media composition for transformation and regeneration |
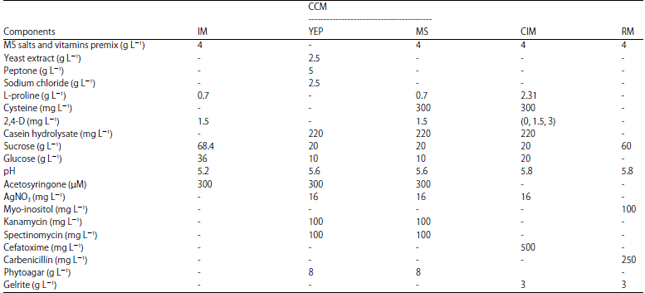 | |
IM: Infection medium, CCM: Co-cultivation media, CIM: Callus Induction medium, RM: Regeneration medium, YEP: Yeast extract peptone modified co-cultivation medium, MS: Murashige and Skoog based co-cultivation medium | |
This media was used as control in this study. After co-cultivation, callus induction was done on callus induction media (CIM) consisting of MS macro and micro salts and vitamins, three varying levels of (0, 1.5 and 3 mg L–1) 2, 4-D, 20 g L–1 sucrose, 20 g L–1 glucose, 2.31 g L–1 proline, 220 mg L–1 casein hydrolysate 3 g L–1 gelrite and pH adjusted to 5.8. After autoclaving, 16 mg L–1 AgNO3, 300 mg L–1 cysteine and 500 mg L–1 cefatoxime were added. Shooting and rooting of embryogenic calli were done in regeneration media (RM) consisting of MS macro and micro salts and vitamins, 100 mg L–1 myo-inositol, 60 g L–1 sucrose, 3 g L–1 gelrite and pH adjusted 5.8. After autoclaving, 250 mg L–1 carbenicillin was added. All media regimes used in this research were slightly modified from Frame et al.10 and Gorji et al.15.
Infection and co-cultivation: Immature zygotic embryos were excised from sterilized whole ears as described by Frame et al.16 into 2 mL Eppendorf tubes containing the infection media. The Agrobacterium strain used in this study was EHA 101 which harboured the GUS gene. The concentrations of Agrobacterium for infection were first optimized ranging between 0.8, 0.2, 0.07 and 0.03 at OD660 using immature embryos of genotype CML 216 in order to determine the optimal concentrations for GUS transient gene expression. Embryos from every inbred line infected with the different Agrobacterium concentration were co-cultivated alone in 3 replicates such that every inbred line had 3 plates for every Agrobacterium concentration. These plates were labelled and incubated in the dark at 18°C for 3 days after which transient GUS assays were done. This study also sought to determine whether the addition of growth regulator 2, 4-D, cysteine and proline in co-cultivation media would enhance the transient GUS expression. Immature maize embryos were co-cultivated in YEP media supplemented with each of these substances and transient expression of the GUS transgene monitored through blue staining.
Induction of embryogenic calli: For induction of embryogenic calli, immature embryos were allowed to rest for 3 days on co-cultivation media 1 and 2 in order to study the possible effects of YEP media on the regeneration frequency of tropical maize compared to MS media. To study the effects of 2,4-Dichlorophenoxyacetic acid (2,4-D) on development of embryogenic calli, embryos from co-cultivation media were plated into callus induction media with different concentrations of 2,4-D (0.0. 1.5 and 3 mg L–1). All embryos were then incubated in the dark at 25°C for 14 days before being sub-cultured into fresh media.
Shooting and rooting: Only embryogenic calli from 1.5 and 3 mg L–1 2,4-D levels were transferred to the shoot and root induction media since most embryos from 0 mg L–1 exhibited preconscious germination and any calli formed were non-embryogenic. These calli were then incubated at a 16/8 light and darkness regime to induce shoots and roots concurrently. After 2 weeks, the number of shoots that formed were counted and recorded. Three week old plantlets were transferred from culture bottles to peat moss in 10 cm diameter plastic pots and covered with a plastic paper for 48 h for gradual acclimatization and hardening. After hardening, the plantlets were transferred into 20 L buckets and developed to maturity.
Histochemical X-Gluc assays: Histochemical analysis of the GUS gene was done as described by Ishida et al.11 and Jefferson et al.17 with some adjustments as followed. After 3 days of co-cultivation, embryos were transferred into 2 mL Eppendorf tubes containing 300 μL tissue fixative, 500 μL 200 mM NaPO4 and 300 μL X-gluc solution and incubated for 24 h at 37°C. After hydrolysis of the X-gluc substrate by the β-glucuronidase enzyme, and reacting with oxygen, only embryos positive for the GUS gene turned blue and were counted under a microscope.
Statistical analysis: Data on all parameters under study were stored in Microsoft excel spread sheets version 2013 and analyzed using multivariate ANOVA at 95% confidence interval with SAS statistical computer software version 9.2. Mean separation was done using Tukey’s pairwise comparison test at 5% probability level. Transient transformation frequencies were calculated as the number of embryos that showed GUS expression over the total number of embryos infected and expressed as a percentage. Callus formation frequency was calculated as the number of embryos forming embryogenic calli over the total number of embryos plated on callus induction media and expressed as a percentage. Regeneration frequency was calculated as the average number of shoots forming per callus/genotype and expressed as a percentage.
RESULTS
Callus induction on YEP and MS media: Immature zygotic embryos from all maize genotypes on YEP callus induction media even when supplemented with either 2,4-D, proline and cysteine failed to form calli. Therefore, to mimic a transformation process which uses YEP media for co-cultivation, immature embryos were first allowed to rest on YEP and MS media for 3 days before proceeding to callus induction media (Table 1). Immature maize embryos that rested for 72 h on YEP media and then placed on MS callus induction media formed both friable embryogenic and non-embryogenic calli which were cream in colour (Fig. 1a, b) unlike calli formed from MS media which were compact and hard, white in colour and most of them being embryogenic (Fig. 1c, d).
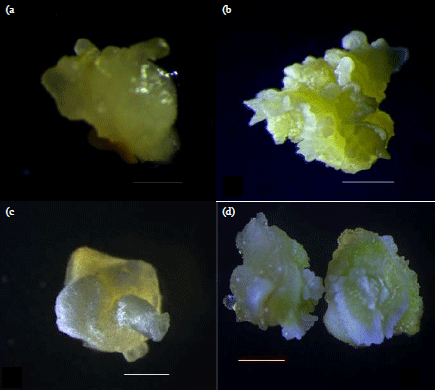 | |
| Fig. 1(a-d): | Embryogenic calli forming from E04 immature embryos pre-cultured in YEP and MS media (a) 14 day old immature embryo pre-cultured on YEP media supplemented with 3 mg L–1 2,4-D, (b) 21 day old immature embryo pre-cultured on YEP media supplemented with 1.5 mg L–1 2,4-D, (c) 7 day old immature embryo pre-cultured on MS media supplemented with 1.5 mg L–1 2,4-D and (d) 21 day old immature embryo pre-cultured on MS media supplemented with 3 mg L–1 2,4-D |
Effect of growth regulator 2,4-D on callus induction on YEP Media: At 1.5 and 3.0 mg L–1 2,4-D, immature embryos were responsive and formed calli. The optimal concentration of the growth regulator was 1.5 mg L–1 with 95.81% callus induction frequency. There was a significantly higher callus formation frequency at 1.5 and 2.0 mg L–1 hormone concentrations than at 0 mg L–1 (p<0.0001), with most calli forming at 1.5 mg L–1 concentration and none forming at 0 mg L–1 hormone concentration (Table 2). Of all the immature embryos pre-rested on MS media at 1.5 mg L–1 2,4-D concentration, inbred line E04 had the highest frequency of callus formation while inbred A04 had the least frequency of callus induction. Inbred line CML 144 had the highest callus formation frequency of on YEP media followed. When the growth regulator concentration was increased to 3 mg L–1 2,4-D immature embryos from CML 144 that were pre-rested on MS media had the highest callus induction frequency and the least callusing frequency observed on A04. However, when immature embryos previously rested on YEP media were subjected to callus formation at 3 mg L–1 2,4-D concentration, the highest callus formation frequency was observed in inbred line CML 216 and the least in CML 144 (Table 2). ANOVA revealed that there was a significantly higher callus formation frequency (p = 0.0288) in embryos previously pre-rested on MS media than those pre-rested YEP media. There was however no significant difference in callus formation based on the different genotypes (p = 0.5459).
Regeneration on MS media: Embryogenic calli were placed on regeneration media (Table 1) in the light and after 10 days, they turned green (Fig. 2a). Shoots appeared 1 week post embryogenic calli turning green. It was observed that the shoots which were formed from calli obtained from embryos rested on both YEP and MS media had no differences in growth and morphology.
| Table 2: | Percentage callus induction frequencies of tropical maize genotypes pre-rested on MS and YEP media at different 2,4-D concentrations |
 | |
Values with the same letter are not significantly different by Tukey’s pair-wise comparison (p<0.05). Means±SE | |
 | |
| Fig. 2(a-e): | Somatic embryogenesis from immature embryos pre-cultured on YEP media and regeneration of E04 tropical genotype, (a) Green somatic embryos growing in light, (b) Shoot and root structures forming, (c) Plantlets forming in regeneration media, (d) Plantlets in acclimatization pots and (e) Fully grown regenerated plant |
| Table 3: | Regeneration frequencies of different inbred maize lines |
 | |
Values with the same letter are not significantly different by Tukey’s pair-wise comparison (p<0.05). Means±SE | |
Data analysis revealed that there was a significantly higher regeneration frequency (p = 0.0001) in inbred maize lines CML 216 and CML 144 than that observed in inbred maize lines E04 and A04, with CML 216 forming most shoots and A04 forming the least shoots (Table 3). Based on the media previously used for pre-resting embryos, there was no significant difference on the regenerability of the calli (p = 0.4783). From 10 regenerated plants of each genotype which were hardened and acclimatized, only two plants from genotype E04, one from each media type exhibited soma clonal variations.
Competence of tropical immature maize embryos to Agrobacterium-mediated transformation
Optimization of Agrobacterium concentration: Optimization of the infection process with CML 216 revealed that Agrobacterium densities of OD660 = 0.07 resulted in the highest transient GUS expression of 36.67% after co-cultivating on YEP media for 3 days compared to 23.33% when co-cultivated on MS media (Fig. 3). Similar pattern were recorded at Agrobacterium concentrations of 0.02 with immature embryos co-cultivated on YEP media resulting in high transient GUS expression of 20% while those co-cultivated on MS media was 10%, concentrations of 0.8 and 0.03 on YEP and MS media recorded transient GUS expression of 16.67 and 6.67%, respectively. In the other maize lines (CML 144, E04 and A04), the transient GUS expression was also high at Agrobacterium concentrations of 0.07 on YEP media than on MS media (Table 4). Thus, the optimal concentration of Agrobacterium was 0.07. The lowest transient GUS expressions were observed at high Agrobacterium concentrations of 0.8 or 0.2 in all genotypes except for genotype E04 co-cultivated on YEP media whose lowest transient GUS expression was 5.56% at OD660 = 0.03. Analysis of variance also revealed that there was a significantly higher transient GUS expression at OD660 = 0.07 than all other Agrobacterium concentrations (p = 0.0002).
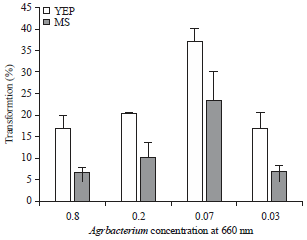 | |
| Fig. 3: | Transformation frequencies of CML 216 inbred line based on transient GUS expression using yeast extract peptone and Murashige and Skoog media |
| Vertical bars represent standard errors | |
To test whether the observed transient GUS expression was not a mere expression of the gene in Agrobacterium,20 T0 plants resulting from immature embryos either co-cultivated in YEP or MS media for the expression of the bar selectable marker gene were screened via PCR. Surprisingly, Five plants regenerated from immature embryos co-cultivated in YEP media were positive while all the 20 plants regenerated from immature embryos co-cultivated in MS media were negative (Fig. 4).
Supplementing YEP media results in differences in transformability: Blue staining was observed in all the transformed embryos (Fig. 5). The addition of 1.5 mg L–1 2,4-D growth regulator to YEP co-cultivation media resulted in lower transient GUS expression than co-cultivating on YEP media without the growth regulator. Except for genotype CML 144 which showed a slight increase in GUS expression from 7.78% on YEP media to 7.79% when co-cultivated on YEP+2,4-D, all other genotypes registered lower transient GUS expression values when co-cultivation was done on YEP+2,4-D compared to transient GUS expression observed when co-cultivation was done on YEP media (Table 5). However, co-cultivating immature maize embryos on YEP+2,4-D media resulted in higher transient GUS expression than co-cultivating on MS media. Analysis of variance however revealed that there was no significant difference in transient GUS expression in all the genotypes when co-cultivation was done on YEP+2,4-D media (p = 0.3566).
The addition of cysteine to YEP co-cultivation media was also observed to impact transient GUS expression negatively. Lower transient GUS expression was observed after co-cultivation was done on YEP+CYS media compared GUS expression observed when co-cultivation was done on YEP media in all genotypes (Table 5).
 | |
| Fig. 4: | Gel electrophoresis of PCR using bar gene specific primers on T0 plants; lines 1-5; putatively transgenic plants co-cultivated on YEP media. MS; plants co-cultivated on MS media negative for the transgene. Negative control was untransformed maize DNA and positive control was pTF 102 plasmid DNA |
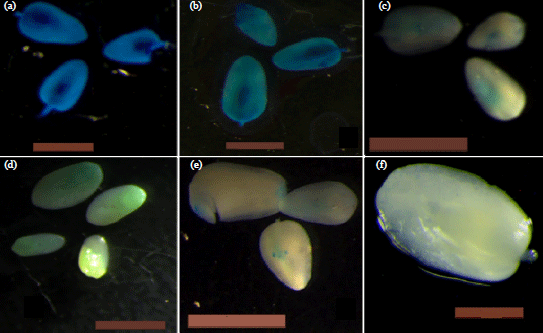 | |
| Fig. 5(a-f): | Immature maize embryos showing different GUS expression levels after co-cultivation in YEP media and MS media (a) CML 144 co-cultivated in YEP media at 0.07 Agrobacterium concentration, (b) A04 embryos co-cultivated in YEP+PRO media at 0.07 Agrobacterium concentration, (c) E04 embryos co-cultivated in YEP+PRO media at 0.2 Agrobacterium concentration, (d) A04 embryos co-cultivated in MS media at 0.8 Agrobacterium concentration, (e) CML 216 embryos co-cultivated in MS media at 0.03 Agrobacterium concentration and (f) Control (YEP without Agrobacterium) negative for GUS staining |
| Table 4: | Transient GUS expression in different maize genotypes after co-cultivating on YEP and MS media |
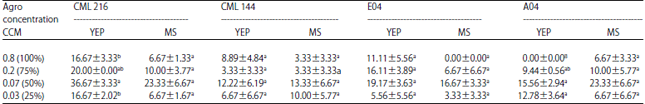 | |
Values with the same letter are not significantly different by Tukey’s pair-wise comparison (p<0.05) Means±SE | |
Analysis of variance also revealed that there was no significant difference in transient GUS expression when immature maize embryos of the different genotypes were co-cultivated on YEP+CYS media (p = 0.5079).
| Table 5: | Effects of different media supplements on transient GUS expression on maize genotypes |
 | |
Values with the same letter are not significantly different by Tukey’s pair-wise comparison (p<0.05) Means±SD | |
The supplementation of YEP co-cultivation media with proline seemed to impact transient transformation positively in three maize inbred lines. All the transient GUS expression values observed from the four genotypes after co-cultivation on YEP+PRO media were higher than those observed when co-cultivation was done on MS media (Table 5). There was a significantly higher transient GUS expression in the four genotypes when co-cultivation was done on YEP+PRO media (p = 0.0254), with the highest transient GUS expression observed in genotype CML 216 (19.17%) while the least transient GUS expression was observed in genotype A04 (7.87%).
Tropical maize inbred lines transformability and regenerability is genotype dependent: The formation of embryogenic calli, the ability of these embryogenic calli to regenerate and the competence of immature embryos to Agrobacterium-mediated transformation were all observed to be dependent on the genotype. When immature embryos were placed on CIM to induce the formation of callus, irrespective of the co-cultivation media previously used to rest immature embryos and the concentration of 2,4-D, statistical analysis revealed that there was a significantly higher callus formation frequency (p = 0.0206) in genotypes E04, CML 144 and CML 216 than that observed in A04. However, when these embryogenic calli were transferred to regeneration media, CML 216 calli were first to form shoots in addition to having most shoots in culture (Table 3). ANOVA also revealed that there was a significantly higher (p = 0.0001) transient GUS expression in CML 216 than all other genotypes. Additionally, the addition of different substances into the co-cultivation media led to un-uniform responses in the maize genotypes under study (Table 5). These observations highlight possible genotype dependence in these maize inbred lines, most of which are considered highly recalcitrant to Agrobacterium-mediated transformation.
DISCUSSION
This study investigated the effect of YEP media on transient transformation of maize immature embryos, the effects of the different co-cultivation media components on transient transformation as well as the regenerability of maize immature embryos after resting of YEP co-cultivation media. In any Agrobacterium-mediated transformation system, developing optical conditions for the transfer of a gene of interest within the T-DNA is a very critical step. Therefore, the choice of co-cultivation media for transformation is very important. Higher transient GUS expression was observed in immature embryos co-cultivated in YEP media when compared to MS media, suggesting that YEP media is superior to MS media in improving the competence of maize immature embryos in Agrobacterium-mediated transformation. This could be because YEP media is an optimal media for the growth of Agrobacterium when compared to MS media that was designed to be optimal for the growth of the explants and has been shown not to properly support the growth of bacteria18. The ability of YEP media to support the growth of Agrobacterium probably ensures a constant availability of actively dividing Agrobacterium cells in the media which were responsible for the observed increased transient transformation. Increase in transient transformation when using YEP+PRO could be attributed to the effect of proline in the media and its interaction with Agrobacterium. Proline is thought to increase the delivery of T-DNA by stimulating the growth of A. tumefaciens as well as supporting the development of plant cells. These results are consistent with the findings of Cao et al.19, who documented that the addition of proline in MS based co-cultivation media increased transient GUS expression and eventually the transformation efficiency in maize. It was also observed by Carvalho et al.20 that the addition of proline and asparagine to co-cultivation media increases transient GUS expression in sorghum. Findings from this study however contrast those of Frame et al.7 and Vega et al.21 who observed that the use of cysteine in MS based and N6 based co-cultivation media increased the frequency of transformation. It is speculated that co-cultivation media supplements like cysteine and 2,4-D do not directly affect the ability of Agrobacterium to infect explants and transfer the Agrobacterium T-DNA. However, sometimes, these substances may increase the frequency of transformed cells by directly affecting the explant or the interaction between the explant and Agrobacterium during co-cultivation. The observation that these substances increase transformation efficiency as observed by other authors could therefore be due to their interaction with the media used. The media in these cases favours the explant and these substances act as nutritional supplements to the explant but not to Agrobacterium.
In this study, Agrobacterium concentration of OD660 = 0.07 resulted in the best transient transformation when using YEP media for co-cultivation. A number of reports have documented differences in transient GUS expressions using different Agrobacterium cell densities/concentrations in different plants. All of these studies report6,12,22–24 optimum Agrobacterium densities of OD600 = 0.4-0.8 for co-cultivation although they involved the use of MS and N6 media for co-cultivation. In this study however, the optimum Agrobacterium cell concentration was OD660 = 0.07, way lower than what has been documented in other studies. The effectiveness of low Agrobacterium cell densities in this study could be because YEP media highly supports the growth of Agrobacterium unlike MS and N6 media hence starting with a low number of infection cells results in a faster growth rate. It has been shown by Gurlitz et al.25 that each plant cell binds a certain finite number of Agrobacterium cells beyond which the plant cell will lose viability and the transformation efficiency declines. Therefore, when using YEP co-cultivation media which is known to highly support the growth of Agrobacterium, the optimum number/ratio of Agrobacterium cells to explant cell is reached faster. When starting with an initial high infection number of bacterial cells (OD660 = 0.8), most of the explant cells lose viability resulting in a much reduced transient GUS expression by the 3rd day of co-cultivation due to a high ratio of Agrobacterium cells binding to the explant. An appropriate initial bacterial infection (OD660 = 0.07) and subsequent YEP co-cultivation ensured optimum Agrobacterium to explant ratio and therefore contributed to the high transient GUS expression observed.
Genotype-dependent response was observed with respect to transient transformation, callusing and regenerability. Generally, E04 was the best genotype with respect to callus formation (73.03%) while CML 216 was the best genotype for transformation and regeneration. Genotype-dependent response is not only limited to maize but has also been reported with other cereals and many other crops in general as documented by Karami26 and Karami et al.27. The susceptibility to Agrobacterium and the response to in vitro tissue culture conditions have indeed been shown to be highly genotype dependent not only in some of these inbred lines but also in other maize genotypes8,14,19,27. All the four genotypes in the study responded differently to Agrobacterium infection after co-cultivating them in different media regimes. Genotype CML 216 showed similarities in transient GUS expression when co-cultivated in (YEP, YEP+PRO) and (YEP+ALL, MS). A188 and Hi-II are temperate maize inbred lines which have been used for tissue culture and are efficiently transformed by Agrobacterium4,7,28 and still exhibit genotype dependent response. This is also the case with the tropical genotypes which have been used in other studies.8,29,30 More recently, Cao et al.19 working on 14 maize inbred lines reported genotype dependence in GUS expression based on their differences in response to Agrobacterium infection. All these genotypes, tropical or temperate display poor agronomic characteristics and have been mostly used in basic studies to test trait genes but none has been used on a commercial scale.
In this study, it was observed that the optimum 2,4-D concentration for callus formation was 1.5 mg L–1. In other studies, the auxin 2,4-D has commonly been used to induce embryogenic calli from maize in ranges of 2-3 mg L–1.31–33 Abebe et al.31 and Odour et al.33 reported optimum 2,4-D concentrations of 2 mg L–1 to form embryogenic calli from tropical maize immature embryos while Jia et al.34 used 3 mg L–1 to form embryogenic calli from mature embryos of tropical maize genotypes. Consistent with these findings, this study recorded best formation of embryogenic calli at 2,4-D concentrations of 1.5 mg L–1 after resting the embryos in YEP and MS co-cultivation media. This, therefore, suggests that YEP media does not significantly affect the callus forming ability and subsequent regenerability of tropical maize. Although unlike embryos rested in MS media for 3 days, some embryos rested on YEP co-cultivation media did not form any calli, suggesting that YEP media might have a slightly effect on the cell signalling pathway and to some extend disrupts the regeneration process of immature maize embryos.
CONCLUSION
In this study, a better co-cultivation media based on yeast extract peptone is reported. All the tropical maize genotypes involved in the study reported a higher transient transformation on YEP as well as YEP+PRO and YEP+ALL media compared to co-cultivation on MS media. Additionally, the use of low Agrobacterium cell densities (OD660 = 0.07) during the co-cultivation step was observed to increase transient transformation. Lastly 2,4-D concentrations of 1.5 mg L–1 was observed to be optimum for callus formation in maize inbred lines and is not affected by the co-cultivation media used. YEP based co-cultivation media as well as the use of low Agrobacterium concentrations at infection and co-cultivation could therefore be a new and better co-cultivating strategy with recalcitrant maize genotypes hence is highly recommended for tropical inbred lines.
SIGNIFICANCE STATEMENTS
This study discovers a frontier transformation procedure with a better transformation rate based on both transient and stable transformation. The study therefore highlights a novel hypothesis on genetic transformation of maize and provides a knowledge insight to readers and a logical platform for further studies in plant genetic transformation.
REFERENCES
- Rhodes, C.A., D.A. Pierce, I.J. Mettler, D. Mascarenhas and J.J. Detmer, 1988. Genetically transformed maize plants from protoplasts. Sci., 240: 204-207.
CrossRefDirect Link - Gordon-Kamm, W.J., T.M. Spencer, M.L. Mangano, T.R. Adams and R.J. Daines et al., 1990. Transformation of maize cells and regeneration of fertile transgenic plants. Plant Cell, 2: 603-618.
CrossRefPubMedDirect Link - Ishida, Y., H. Saito, S. Ohta, Y. Hiei, T. Komari and T. Kumashiro, 1996. High efficiency transformation of maize (Zea mays L.) mediated by Agrobacterium tumefaciens. Nat. Biotechnol., 15: 745-750.
CrossRefPubMedDirect Link - Que, Q., S. Elumalai, X. Li, H. Zhong and S. Nalapalli et al., 2014. Maize transformation technology development for commercial event generation. Front. Plant Sci., Vol. 5.
CrossRefDirect Link - Huang, X. and Z. Wei, 2005. Successful Agrobacterium-mediated genetic transformation of maize elite inbred lines. Plant Cell Tissue Organ Culture, 83: 187-200.
CrossRefDirect Link - Frame, B.R., J.M. McMurray, T.M. Fonger, M.L. Main and K.W. Taylor et al., 2006. Improved Agrobacterium-mediated transformation of three maize inbred lines using MS salts. Plant Cell Rep., 25: 1024-1034.
CrossRefDirect Link - Ombori, O., J.V.O. Muoma and J. Machuka, 2013. Agrobacterium-mediated genetic transformation of selected tropical inbred and hybrid maize (Zea mays L.) lines. Plant Cell Tissue Organ Cult., 113: 11-23.
CrossRefDirect Link - Sande, F.O., F.M. Njoka, A.J. Mgutu and S.E. Anami, 2014. Screening for water deficit tolerance, relative growth analysis and Agrobacterium-infectivity among tropical maize (Zea mays L.) inbred lines in Nairobi, Kenya. Int. J. Life Sci. Res., 2: 218-224.
Direct Link - Frame, B.R., H. Shou, R. Chikwamba, Z. Zhang and C. Xiang et al., 2002. Agrobacterium-mediated transformation of maize embryos using a standard binary vector system. Plant Physiol., 129: 13-22.
Direct Link - Ishida, Y., Y. Hiei and T. Komari, 2007. Agrobacterium-mediated transformation of maize. Nat. Protocols, 2: 1614-1621.
CrossRefDirect Link - Joyce, P., M. Kuwahata, N. Turner and P. Lakshmanan, 2010. Selection system and co-cultivation medium are important determinants of Agrobacterium-mediated transformation of sugarcane. Plant Cell Rep., 29: 173-183.
CrossRefDirect Link - Ombori, O., J.O. Muoma and J. Machuka, 2013. Effect of the medium used in the growth of Agrobacterium strain on subsequent gus expression of the infected immature zygotic embryos of the tropical maize line. Maize Genet. Cooperation Newslett., Vol. 87.
Direct Link - Anami, S.E., A.J. Mgutu, C. Taracha, G. Coussens and M. Karimi et al., 2010. Somatic embryogenesis and plant regeneration of tropical maize genotypes. Plant Cell Tissue Organ. Cult., 102: 285-295.
CrossRefDirect Link - Gorji, A.H., M. Zolnoori, A. Jamasbi and Z. Zolnoori, 2011. In vitro plant generation of tropical maize genotypes. Proceedings of the International Conference on Environmental, Biomedical and Biotechnology, Volume 16, August 19-21, 2011, Shanghai, China, pp: 52-59.
Direct Link - Jefferson, R.A., T.A. Kavanagh and M.W. Bevan, 1987. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J., 6: 3901-3907.
Direct Link - Leifert, C. and W.M. Waites, 1992. Bacterial growth in plant tissue culture media. J. Applied Bacteriol., 72: 460-466.
CrossRefDirect Link - Carvalho, C.H.S., U.B., N. Gunaratna, J. Anderson, H.H. Kononowicz, T.K. Hodges and J.D. Axtell, 2004. Agrobacterium-mediated transformation of sorghum: Factors that affect transformation efficiency. Genet. Mol. Biol., 27: 259-269.
CrossRefDirect Link - Vega, J.M., W. Yu, A.R. Kennon, X. Chen and Z.J. Zhang, 2008. Improvement of Agrobacterium-mediated transformation in Hi-II maize (Zea mays) using standard binary vectors. Plant Cell Rep., 27: 297-305.
CrossRefPubMedDirect Link - Amoah, B.K., H. Wu, C. Sparks and H.D. Jones, 2001. Factors influencing Agrobacterium-mediated transient expression of uid A in wheat inflorescence tissue. J. Exp. Bot., 52: 1135-1142.
CrossRefDirect Link - Kim, M.J., K. Baek and C.M. Park, 2009. Optimization of conditions for transient Agrobacterium-mediated gene expression assays in Arabidopsis. Plant Cell Rep., 28: 1159-1167.
CrossRefDirect Link - Yenchon, S. and S. Te-Chato, 2012. Effect of bacteria density, inoculation and co-cultivation period on Agrobacterium-mediated transformation of oil palm embryogenic callus. J. Agric. Technol., 8: 1485-1496.
Direct Link - Gurlitz, R.H., P.W. Lamb and A.G. Matthysse, 1987. Involvement of carrot cell surface proteins in attachment of Agrobacterium tumefaciens. Plant Physiol., 83: 564-568.
CrossRefDirect Link - Karami, O., 2008. Factors affecting Agrobacterium-mediated transformation of plants. Transgenic Plant J., 2: 127-137.
Direct Link - Karami, O., M. Esna-Ashari, G.K. Kurdistani and B. Aghavaisi, 2009. Agrobacterium-mediated genetic transformation of plants: The role of host. Biol. Planta., 53: 201-212.
CrossRefDirect Link - Zhao, Z.Y., W. Gu, T. Cai, L. Tagliani and D. Hondred et al., 2002. High throughput genetic transformation mediated by Agrobacterium tumefaciens in maize. Mol. Breed., 8: 323-333.
CrossRefDirect Link - Omer, R.A., J.M. Matheka, A.M. Ali and J. Machuka, 2013. Transformation of tropical maize with the NPK1 gene for drought tolerance. Int. J. Genet. Eng., 3: 7-14.
Direct Link - Abebe, D.Z., W. Teffera and J.S. Machuka, 2008. Regeneration of tropical maize lines (Zea mays L.) from mature zygotic embryo through callus initiation. Afr. J. Biotechnol., 7: 2181-2186.
Direct Link - Binott, J.J., J.M. Songa, J. Ininda, E.M. Njagi and J. Machuka, 2008. Plant regeneration from immature embryos of Kenyan maize inbred lines and their respective single cross hybrids through somatic embryogenesis. Afr. J. Biotechnol., 7: 981-987.
Direct Link - Oduor, R.O., E.N.M. Njagi, S. Ndung'u and J.S. Machuka, 2006. In vitro regeneration of dryland Kenyan maize genotypes through somatic embryogenesis. Int. J. Bot., 2: 146-151.
CrossRefDirect Link - Jia, X., J. Zhang, H. Wang and W. Kong, 2008. Efficient maize (Zea mays L.) regeneration derived from mature embryos in vitro. Maydica, 53: 239-248.
Direct Link