
Dedi Ruswandi
Faculty of Agriculture, Padjadjaran University, 40600 Bandung, Indonesia
LiveDNA: 62.18773
Budi Waluyo
Faculty of Agriculture, Brawijaya University, Malang, Indonesia
Andi Takdir Makkulawu
Indonesia Center for Cereals Research, Maros, Indonesia
Elia Azizah
Faculty of Agriculture, Universitas Singaperbangsa, Karawang, Indonesia
Yuyun Yuwariah
Faculty of Agriculture, Padjadjaran University, 40600 Bandung, Indonesia
Neni Rostini
Faculty of Agriculture, Padjadjaran University, 40600 Bandung, Indonesia
Asian Journal of Crop Science
Year: 2017 | Volume: 9 | Issue: 4 | Page No.: 141-148







ABSTRACT
Background and Objective: The assessment of the genetic diversity of new inbred lines has proven significant to the improvement of maize varieties. The objective of the study was to evaluate the diversity and relatedness of new Indonesian elite maize lines. Materials and Methods: Genetic diversity of 48 Indonesian elite maize lines was studied using six SSR markers. Some parameters were estimated including genetic relationship and genetic diversity. Genetic relationship was assessed using Jaccard’s similarity coefficient and was used to draw a neighbourhood-joining tree (NJT) by circle cluster using MEGA-pc software. Principal coordinate analysis (PCoA) was used for the estimation of genetic diversity. Results: All SSR loci detected in all genotypes were polymorphic. The population in this study is diverse as estimated by its polymorphic index coefficient (PIC). Population of 20 Indonesian maize lines was divided into 3 sub-groups, whereas 28 of mutant population was clustered into 7 sub-groups. Conclusion: SSR marker is a meaningful marker for genetic diversity study of Indonesian maize core collection. The forty-eight Indonesian maize genotypes were clustered into three major groups. These findings can be used in breeding methods for the development of populations.
PDF Abstract XML References Citation
Received: August 24, 2017;
Accepted: October 02, 2017;
Published: October 17, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Dedi Ruswandi, Budi Waluyo, Andi Takdir Makkulawu, Elia Azizah, Yuyun Yuwariah and Neni Rostini, 2017. Simple Sequence Repeats Analysis of New Indonesian Maize Inbred. Asian Journal of Crop Science, 9: 141-148.
DOI: 10.3923/ajcs.2017.141.148
URL: https://scialert.net/abstract/?doi=ajcs.2017.141.148
DOI: 10.3923/ajcs.2017.141.148
URL: https://scialert.net/abstract/?doi=ajcs.2017.141.148
INTRODUCTION
Simple sequence repeats (SSRs) is an effective molecular marker to select superior genotypes in early growth with desirable agronomic traits. SSR markers application in crop improvement includes DNA fingerprinting and genetic diversity analysis, assigning inbred lines to heterotic groups, high throughput genotyping and phenotyping, mapping both association and QTL and marker assisted selection for doubled haploid technology1. Plant breeders, therefore, applied SSRs in their programs for the reasons of low cost, consistency of product for every marker, a high amenability to multiplex PCR and simplicity and effectiveness1,2.
Diversity and similarity among inbred are important for breeder in selecting parental lines for developing new superior hybrids by reducing huge number of crosses combination. Some researchers reported the application of SSRs in barley and wheat2, maize3,4, cotton5, groundnut6, peanut7, soybean8, oat9, carrot10 and melon11. Genetic diversity studies on maize using SSR markers have been reported by many researchers in China4,12, India13, Eastern and Southern Africa14 and Mexico15. Masuka et al.14 assessed genetic diversity of parental from CIMMYT ESA for hybrid breeding using SSR markers to avoid the narrowing down of the genetic base. Abakemal et al.16 applied SSR to determine genetic diversity among and within quality protein maize (qpm) and non-qpm line in the hope of conversion of normal maize into quality protein maize. The use of SSR markers to evaluate the genetic diversity of mutant and parental lines was suggested by Ruswandi et al.17 since their ability to cluster those genotypes into different major group.
Maize development program in West Java was initiated from 2001 by crossing CIMMYT lines with local land races and by means gamma ray irradiation. The program produced 20 new inbred lines derived from crossing and 28 new potential mutant lines17. Some mutant lines were informed as high yield parental lines for hybrids18, a shading tolerant line for cultivation under maize/ Albizia agroforestry system19 and a high nutritious line for industry20. This study evaluated the diversity and relatedness of 48 Indonesian elite maize lines using SSR markers.
MATERIALS AND METHODS
The study was carried out from November, 2016 up to June, 2017 at the Plant Breeding and Seed Technology Laboratory, Padjadjaran University, Indonesia. DNA of 48 Indonesian maize genotypes consisting of 20 DR lines and 28 mutant DR were evaluated using SSR markers. The protocol of Asian Maize Biotechnology Network (AMBIONET), Agricultural Biotechnology Center (CIMMYT ABC), Philippines, was used including: DNA extraction, SSR amplification, separation and visualization21.
Selected primers for important traits, such as: SSR markers for resistance to downy mildew pathogens, tolerance to drought and tolerance to shading was used to amplify their DNA. These SSR markers included Bngl 137, Umc 1149, Phi 112, Phi 113, Phi 36, Bngl 589, Phi 126 and Bngl 125. The polymerase chain reactions (PCR) with Mastercycler ep gradient (eppendorf) were done using KAPA2G Fast Ready Mix (Boston, Massachusetts, USA) which contained 10 mL of reaction mix each of forward and reverse primer and 20 ng of genomic DNA. The profile of DNA amplification were: One cycle of initial denaturation (30 cycles, 93°C, 1 min), annealing (56°C/58°C/60°C, 2 min) and extension (72°C, 2 min). It was, then, followed by 1 cycle of final extension (72°C, 5 min). The SSR products were run on polyacrylamide gel in 1× TBE (70-75 W, 45 min). The gel was stained in Promega silver stain solution.
Replicated DNAs were scored based on the presence (1) or absence (0) of amplified products. Estimation of major allele, allele number, gene diversity, polymorphic index coefficient (PIC) used the program PowerMarker22. Genetic distances (GD) between pairs of lines were calculated as GD = 1-GS, where GS was Jaccard similarity coefficients. GS and GD were assessed using NTSYS-pc software23. The GD were estimated to construct a neighbourhood joining tree (NJT) by circle cluster using MEGA-pc software version 624. Principal coordinate analysis (PCoA) was also estimated for assessing of genetic diversity.
RESULTS AND DISCUSSION
Major allele, allele number, gene diversity and PIC of line, mutant and line-mutant population assessed by SSR markers are presented in Table 1. All SSR loci detected were polymorphic as shown by its allele/locus and its major allele frequency. The average of allele/locus was 8 for line, 14 for mutant and 16.5 for line-mutant. The mean of major allele frequency was 0.34 for line, 0.28 for mutant and 0.21 for line- mutant. Allele number in this study was higher than that estimated by Makumbi et al.25, Krishna et al.26, Babu et al.27 and Pandey et al.28,29, but lower than values previously assessed by Rocandio-Rodriguez et al.15. Abakemal et al.16 explained that the variation in average of allele number from extensive studies could have originated from the source of germplasm, sample size and repeat length of the SSRs used. Choukan and Warburton30 justified that dinucleotide SSR exhibits high numbers of alleles than other types of SSR loci.
| Table 1: | Allele frequency and number, gene diversity and PIC of line, mutant and line- mutant population |
 | |
 | |
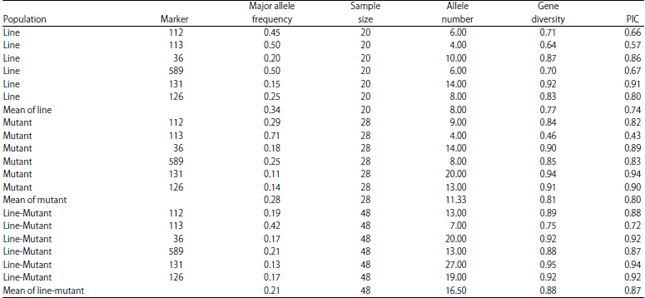
| Fig. 1: | Histogram of genetic distance from 20 Indonesian maize lines based on shared-allele distance |
Coefficient of co-phenetic correlation between genetic distance and allele frequency based on shared allele distance of lines at 0.93 | |
The high numbers of lines to evaluate increase chance of finding rare alleles as compared to small samples. They also stated that gene diversity corresponds to the projected heterozygousity for diploid data.
The line, mutant and mutant population in this study are diverse as estimated by its gene diversity and polymorphic index coefficient (PIC). The average of gene diversity was 0.77 for line, 0.81 for mutant and 0.88 for line- mutant. The mean of PIC was 0.74 for line, 0.80 for mutant and 0.87 for line-mutant. PIC indicated the inequitable power of any locus by considering the frequency and the number of alleles per locus in the population designating gene diversity of population21.
 | |
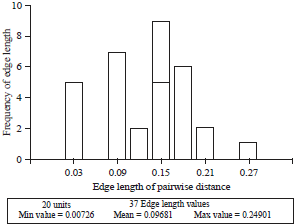
| Fig. 2: | Histogram of genetic distance from 28 Indonesian maize mutant based on shared-allele distance |
Coefficient of co-phenetic correlation between genetic distance and allele frequency based on shared allele distance of mutant at 0.90 | |
PIC value of maize population of this study was higher than that estimated by Pandey et al.29 and Abakemal et al.16; but lower than values previously assessed by Rocandio-Rodriguez et al.15. Choukan and Warburton30 and Xia et al.31 reported that high PIC values were as a result of wide diversity and number of genotypes used and SSR repeat type. Based on the above results, it is concluded that of this study mutant population is more diverse than their original line population. Therefore, mutation breeding can be used to improve genetic base population in Indonesia.
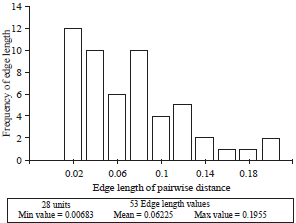
Genetic distance of line population, mutant population and line-mutant population are shown in Fig. 1-3, respectively. Genetic distance of line population based on their combined allele matrix differ from 0.01 up to 0.25 with an average of 0.10 (Fig. 1). This shows that there are high similarities within lines in the population. The allele frequency and genetic distance is closely related. The grouping of the lines shows that it is genetically fit, as is indicated by the cophenetic correlation coefficient of 0.93. Genetic distance of mutant population differed between 0.00-0.19 with an average of 0.06 (Fig. 2). Coefficient of cophenetic correlation of the mutant, however, is 0.90 showing similarities among the mutants. Genetic distance of line- mutant population varied between 0.00-0.25 with an average of 0.07 (Fig. 3). The coefficient of cophenetic correlation between genetic distance and allele frequency based on their shared allele matrix of 0.93 showed similarities among the mutants.
 | |
| Fig. 3: | Histogram of genetic distance from 48 Indonesian maize genotypes based on shared-allele distance |
Coefficient of co-phenetic correlation between genetic distance and allele frequency based on shared allele distance of lines and mutant at 0.93 | |
The population of 20 Indonesian maize lines was divided into 3 sub-groups (Fig. 4). They were ten lines derived from the combination of downy mildew resistant (DMR) and QPM lines (DR #1-10), five lines originated from the selection of DMR lines (DR # 11-15) and five lines formed by phenotypic selection of QPM lines (DR #16-20). This grouping is similar to the NJ analysis in which each group was clustered following DMR or QPM traits. The mutant population was clustered into 7 sub-groups (Fig. 5).
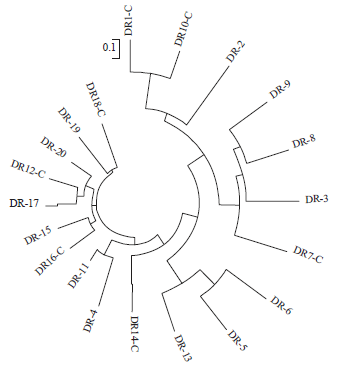 | |
| Fig. 4: | Neighbour-joining tree based on shared-allele distance matrix on 20 lines population |
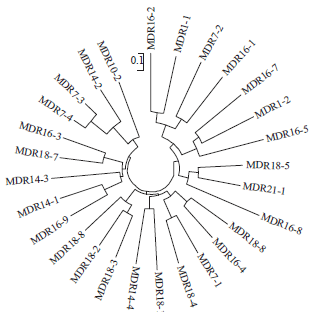 | |
| Fig. 5: | Neighbour-joining tree based on shared-allele distance matrix on 28 mutant population |
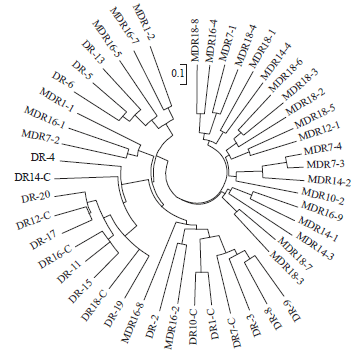 | |
| Fig. 6: | Neighbour-joining tree based on shared-allele distance matrix on 48 lines, both non-mutant and mutants Indonesian maize |
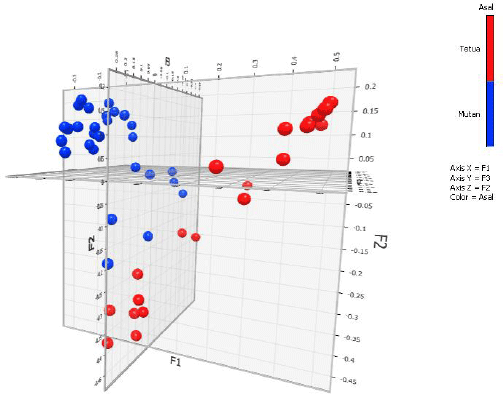 | |
| Fig. 7: | Population grouping of inbred lines (ancestors) and mutant based on principal coordinate analysis (PCoA) |
The position of the mutant in the sub-group was randomly distributed. Mutation, therefore, randomly and unpredictably changes the gene and the position of the locus. From the whole analysis of line-mutant population, there were three grouping based on their positions, i.e: line, mutant and line-mutant (Fig. 6). This shows that there is genetic differentiation based on SSR markers. The different groupings in the line and mutant populations indicated new genetic constitution in the mutant. On the contrary, the mutant- line grouping showed that mutation does not extremely change the whole genetic constitution. This means that mutation randomly changes genetic constitution of the line and express new traits.
The principal coordinate analysis (PCoA) was completed to check the diversity structure of new Indonesian maize population (Fig. 7). The result showed that the population of line and mutant can clearly be differentiated into genetic groupings based on their breeding methods. There are 3 genetic groups of population, namely lines population, mutant population and mixed mutant-lines population. This means that line and mutant population genetically differed. But, within the line and within mutant populations close relations with many similarities were shown. The PCoA have been widely used to confirm the structure and to acquire knowledge about genetic diversity of many crops, such as oat9, rice32, maize33, lentils34, chickpea35, durum wheat36 and garcinia37. This information, in turn, can be used for utilization of the genetic resources in genomic and breeding programs. Boczkowska and Tarczyk9 had confirmed the diversity of polish oat landraces into three different landraces group using PCoA. Based on similar analysis using PCoA, Choudhury et al.32 clustered North-Eastern region of India rice into three groups based on their population structure. Using the same analysis of PCoA, Semagn et al.33 studied genetic diversity among open-pollinated maize varieties in Eastern and Southern Africa and grouped them into 2-4 clusters depending on maturity groups, breeding programs, mega-environments and specific agronomic traits. Khazaei et al.34 categorized lentil accessions into three major groups, namely: South Asia, Mediterranian and Northern temperate, which reflected world’s agro-ecological zones based on PCoA. This study revealed that the clusters reflected the origins, pedigrees and breeding histories of the germplasm. Based on the results of our study, it is suggested that maize breeding programs, particularly hybrid programs have to deliberately use substantial genetic diversity within Indonesian maize inbred, as clustered mutant and non-mutant lines, in order to avoid narrow genetic diversity.
CONCLUSION
SSR markers were valuable for revealing genotypic diversity with PIC as indicated by the range from 0.57-0.94. The forty-eight Indonesian maize genotypes were clustered into three major groups are related with the breeding method to apply in the development of the populations. Thus, SSR markers are a meaningful marker for genetic diversity study of Indonesian maize breeding materials.
SIGNIFICANCE STATEMENTS
This study discovers the diversity of Indonesian maize mini core collection that can be beneficial for maize breeding program in Indonesia. This study will help researchers to uncover the critical areas of plant genetic resources of tropical maize that many researchers were not able to explore. Thus a new theory on utilization of new resources of gene into maize breeding may be arrived at.
ACKNOWLEDGMENTS
The authors were highly grateful for the Ministry of Agriculture, Republic Indonesia and Ministry of Research, Technology and Higher Education, Republic Indonesia for granting the research fund through National Competitive Grant KKP3N 2015 and PUPT 2017.
REFERENCES
- Prasanna, B.M., K. Pixley, M.L. Warburton and C.X. Xie, 2010. Molecular marker-assisted breeding options for maize improvement in Asia. Mol. Breed., 26: 339-356.
CrossRefDirect Link - Hayden, M.J., T.M. Nguyen, A. Waterman, G.L. McMichael and K.J. Chalmers, 2008. Application of multiplex-ready PCR for fluorescence-based SSR genotyping in barley and wheat. Mol. Breed., 21: 271-281.
CrossRefDirect Link - Bedoya, C.A., S. Dreisigacker, S. Hearne, J. Franco and C. Mir et al., 2017. Genetic diversity and population structure of native maize populations in Latin America and the Caribbean. PLoS ONE, Vol. 12.
CrossRefDirect Link - Zou, C.Y., L.J. Li, K.C. Yang, G.T. Pan and T.Z. Rong, 2010. Effects of mass selection on maize synthetic populations. Acta Agron. Sin., 36: 76-84.
CrossRefDirect Link - Hinze, L.L., P.J. Horn, N. Kothari, J.K. Dever, J. Frelichowski, K.D. Chapman and R.G. Percy, 2015. Nondestructive measurements of cottonseed nutritional trait diversity in the U.S. National Cotton Germplasm Collection. Crop Sci., 55: 770-782.
CrossRefDirect Link - Shasidhar, Y., M.K. Vishwakarma, M.K. Pandey, P. Janila and M.T. Variath et al., 2017. Molecular mapping of oil content and fatty acids using dense genetic maps in groundnut (Arachis hypogaea L.). Frontiers Plant Sci., Vol. 8.
CrossRef - Pandey, M.K., H.D. Upadhyaya, A. Rathore, V. Vadez and M.S. Sheshshayee et al., 2014. Genomewide association studies for 50 agronomic traits in peanut using the 'reference set' comprising 300 genotypes from 48 countries of the semi-arid tropics of the world. PLoS ONE, Vol. 9.
CrossRefDirect Link - Dong, D., X. Fu, F. Yuan, P. Chen and S. Zhu et al., 2014. Genetic diversity and population structure of vegetable soybean (Glycine max (L.) Merr.) in China as revealed by SSR markers. Genet. Resour. Crop Evol., 61: 173-183.
CrossRefDirect Link - Boczkowska, M. and E. Tarczyk, 2013. Genetic diversity among Polish landraces of common oat (Avena sativa L.). Genet. Resour. Crop Evol., 60: 2157-2169.
CrossRefDirect Link - Baranski, R., A. Maksylewicz-Kaul, T. Nothnagel, P.F. Cavagnaro, P.W. Simon and D. Grzebelus, 2012. Genetic diversity of carrot (Daucus carota L.) cultivars revealed by analysis of SSR loci. Genet. Resour. Crop Evol., 59: 163-170.
CrossRefDirect Link - Raghami, M., A.I. Lopez-Sese, M.R. Hasandokht, Z. Zamani, M.R.F. Moghadam and A. Kashi, 2014. Genetic diversity among melon accessions from Iran and their relationships with melon germplasm of diverse origins using microsatellite markers. Plant Syst. Evol., 300: 139-151.
CrossRefDirect Link - Liu, L., Y.D. Zhang, H.Y. Li, Y.Q. Bi and L.J. Yu et al., 2016. QTL mapping for gray leaf spot resistance in a tropical maize population. Plant Dis., 100: 304-312.
CrossRefDirect Link - Gupta, H.S., F. Hossain and V. Muthusamy, 2015. Biofortification of maize: An Indian perspective. Indian J. Genet. Plant Breed., 75: 1-22.
CrossRefDirect Link - Masuka, B.P., A. van Biljon, J.E. Cairns, B. Das and M. Labuschagne et al., 2017. Genetic diversity among selected elite CIMMYT maize hybrids in East and Southern Africa. Crop Sci., 57: 2395-2404.
CrossRefDirect Link - Rocandio-Rodriguez, M., A. Santacruz-Varela, L. Cordova-Tellez, H. Lopez-Sanchez, F. Castillo-Gonzalez, R. Lobato-Ortiz and J.J. Garcia-Zavala, 2014. Detection of genetic diversity of seven maize races from the high central valleys of Mexico using microsatellites. Maydica, 59: 144-151.
Direct Link - Abakemal, D., S. Hussein, J. Derera and K. Semagn, 2015. Genetic purity and patterns of relationships among tropical highland adapted quality protein and normal maize inbred lines using microsatellite markers. Euphytica, 204: 49-61.
CrossRefDirect Link - Ruswandi, D., Agustian, E.P. Anggia, A.O. Canama, H. Marta, S. Ruswandi and E. Suryadi, 2014. Mutation breeding of maize for anticipating global climate change in Indonesia. Asian J. Agric. Res., 8: 234-247.
CrossRefDirect Link - Ruswandi, D., J. Supriatna, B. Waluyo, A.T. Makkulawu, E. Suryadi, Z.U. Chindy and S. Ruswandi, 2015. GGE biplot analysis for combining ability of grain yield and early maturity in maize mutant in Indonesia. Asian J. Crop Sci., 7: 160-173.
CrossRefDirect Link - Syafii, M., I. Cartika and D. Ruswandi, 2015. Multivariate analysis of genetic diversity among some maize genotypes under maize-albizia cropping system in Indonesia. Asian J. Crop Sci., 7: 244-255.
CrossRefDirect Link - Marta, H., E. Suryadi and D. Ruswandi, 2017. Chemical composition and genetics of indonesian maize hybrids. Am. J. Food Technol., 12: 116-123.
CrossRefDirect Link - Liu, K. and S.V. Muse, 2005. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics, 21: 2128-2129.
CrossRefDirect Link - Tamura, K., G. Stecher, D. Peterson, A. Filipski and S. Kumar, 2013. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol., 30: 2725-2729.
CrossRefPubMedDirect Link - Makumbi, D., F.J. Betran, M. Banziger and J.M. Ribaut, 2011. Combining ability, heterosis and genetic diversity in tropical maize (Zea mays L.) under stress and non-stress conditions. Euphytica, 180: 143-162.
CrossRefDirect Link - Krishna, M.S.R., S.S. Reddy and V.C.B. Naik, 2012. Assessment of genetic diversity in Quality Protein Maize (QPM) lines using Simple Sequence Repeat (SSR) markers. Afr. J. Biotechnol., 11: 16427-16433.
Direct Link - Babu, B.K., P. Pooja, J.C. Bhatt and P.K. Agrawal, 2012. Characterization of Indian and exotic quality protein maize (QPM) and normal maize (Zea mays L.) inbreds using simple sequence repeat (SSR) markers. Afr. J. Biotechnol., 11: 9691-9700.
Direct Link - Pandey, N., F. Hossain, K. Kumar, A.K. Vishwakarma and V. Muthusamy et al., 2015. Microsatellite marker-based genetic diversity among Quality Protein Maize (QPM) inbreds differing for kernel iron and zinc. Mol. Plant Breed., 6: 1-10.
CrossRefDirect Link - Pandey, N., F. Hossain, K. Kumar, A.K. Vishwakarma and V. Muthusamy et al., 2016. Molecular characterization of endosperm and amino acids modifications among quality protein maize inbreds. Plant Breed., 135: 47-54.
CrossRefDirect Link - Choukan, R. and M.I. Warburton, 2005. Use of SSR data to determine relationships among early maturing Iranian maize inbred lines. Maydica, 50: 163-170.
Direct Link - Xia, X.C., J.C. Reif, D.A. Hoisington, A.E. Melchinger, M. Frisch and M.L. Warburton, 2004. Genetic diversity among CIMMYT maize inbred lines investigated with SSR markers: I. lowland tropical maize (Plant genetic resources). Crop Sci., 44: 2230-2237.
Direct Link - Choudhury, D.R., N. Singh, A.K. Singh, S. Kumar and K. Srinivasan et al., 2014. Analysis of genetic diversity and population structure of rice germplasm from North-Eastern region of India and development of a core germplasm set. PLoS ONE, Vol. 9.
CrossRefDirect Link - Semagn, K., C. Magorokosho, V. Ogugo, D. Makumbi and M.L. Warburton, 2014. Genetic relationships and structure among open-pollinated maize varieties adapted to Eastern and Southern Africa using microsatellite markers. Mol. Breed., 34: 1423-1435.
CrossRefDirect Link - Khazaei, H., C.T. Caron, M. Fedoruk, M. Diapari and A. Vandenberg et al., 2016. Genetic diversity of cultivated lentil (Lens culinaris Medik.) and its relation to the world's agro-ecological zones. Front. Plant Sci., Vol. 7.
CrossRefDirect Link - Torutaeva, E., A. Asanaliev, M.L. Prieto-Linde, A. Zborowska, R. Ortiz, T. Bryngelsson and L. Garkava-Gustavsson, 2014. Evaluation of microsatellite-based genetic diversity, protein and mineral content in chickpea accessions grown in Kyrgyzstan. Hereditas, 151: 81-90.
CrossRefDirect Link - Baloch, F.S., A. Alsaleh, M.Q. Shahid, V. Ciftci and L.E.S. de Miera et al., 2017. A whole genome DArTseq and SNP analysis for genetic diversity assessment in durum wheat from central fertile crescent. PLoS ONE, Vol. 12.
CrossRefDirect Link - Tharachand, C., C.I. Selvaraj and Z. Abraham, 2015. Molecular insights into the genetic diversity of Garcinia cambogia germplasm accessions. Braz. Arch. Biol. Technol., 58: 765-772.
CrossRefDirect Link