
Gehan G. Mostafa
Department of Horticulture (Ornamental Plants), Faculty of Agriculture, Beni Suef University, Egypt
Asian Journal of Crop Science
Year: 2016 | Volume: 8 | Issue: 2 | Page No.: 60-65







ABSTRACT
Background: Sunflowers are widely grown commercially for the oil and they are good lasting as cut flowers. The study aimed to improve the branching mutant of Helianthus annuus for their No. of inflorescences per plant as ornamental plant. Methodology: Three cycles of mass selection were applied from 2011 to 2013. Evaluation of their performance during two seasons was done during 2014 and 2015. Results: A gradual increase was found during the three cycles of mass selection (C1, C2 and C3) for most studied traits in both seasons as well as in combined analysis. No significant differences were found in the No. of florets per inflorescence, while a slightly increase was obtained. Mass selection had negative effect in the earliness of flowering. Number of inflorescences per plant was increased by 16.3, 19.7 and 61.4% for C1, C2 and C3, respectively. Positive and high correlation coefficients were found between the No. of inflorescences per plant and other traits except for the No. of seed per main head where moderate correlation was extended (0.66). Conclusion: Mass selection was a successful method to improve the branching mutant of Helianthus annuus and produce a new cultivar with better characteristics as ornamental plants.
PDF Abstract XML References Citation
Received: January 30, 2016;
Accepted: February 23, 2016;
Published: March 15, 2016
How to cite this article
Gehan G. Mostafa, 2016. Improvement in the Branching Mutant of Helianthus annuus by Mass Selection. Asian Journal of Crop Science, 8: 60-65.
DOI: 10.3923/ajcs.2016.60.65
URL: https://scialert.net/abstract/?doi=ajcs.2016.60.65
DOI: 10.3923/ajcs.2016.60.65
URL: https://scialert.net/abstract/?doi=ajcs.2016.60.65
INTRODUCTION
Sunflower (Helianthus annuus) is one of the most important species belongs to the family Asteraceae (Compositae). Sunflowers are widely grown commercially for the oil that is extracted from the seeds. Seeds are also used to feed birds. All sunflowers are good and long lasting as cut flowers1,2. Induced mutations have been applied for the past 40 years to produce mutant cultivars in sunflower by changing plant characteristics for significant increase in plant productivity3,4. Genetic variability was induced by mutagens treatments5.
Sodium azide treatments produced branched- mutant in Helianthus annuus in the previous studies2. Branching is initiated from axillary meristems in leaf axils on the primary shoot. These meristems give rise to axillary buds which remain dormant or grow out into a branch. Three phytohormones (auxin, cytokinen and strigolacton that is carotenoid-derived and inhibit bud outgrowth) and genes are associated with their homeostasis and signaling are thought to be largely responsible for the regulation of branching. Additionally, genes related to gibberellic acid and polyamine metabolism and genes encoding transcription factors play important roles in branch growth. Shoot branching is a major characteristic which can affect phenotypes such as flowering6.
Mass selection has been used for cultivar improvement in sunflower for many years and it was effective in developing cultivars with early maturity, higher oil percentage and resistance to diseases. The efficiency of mass selection depends on gene effects of the selected traits, their heritability, sample size and genotype-environment interaction. Mass selection is effective for characters controlled by additive genes7.
The present study was carried out to improve the number of inflorescences per plant and to evaluate the performance of three cycles of mass selection in the branching mutant of sunflower that having many inflorescences per plant.
MATERIALS AND METHODS
The study was carried out at the Nursery of Ornamental Plants, Faculty of Agriculture, South Valley University, Qena, Egypt from 2011 to 2015.
Plant materials and procedures: The branching mutant of Helianthus annuus which have many inflorescences/plant was used in this study. The mutant was obtained from previous study on the cv. Giza 102 after sodium azide treatment at 100 ppm in the M2 generation2. Seeds of the selected mutant as a base population were sown in the field on May, 2011 in a non-replicated area. The procedure of mass selection was done. Fifty plants with many inflorescences at axisymmetrical form along the stem and with largest stem diameter were selected (5% selection intensity). While, the other plants were discarded before inflorescences was opening. Equal parts of the seeds from the selected plants were bulked together. Seeds were sown on May, 2012. The same procedure of mass selection was done to form the second cycle (C2) which was sown in May, 2013. The same procedures of mass selection repeated to produce the third cycle of mass selection (C3).
On 15 May, 2014 and 2015, seeds of unbranched parent (cv. Giza 102), base population (branched mutant) and three cycles of mass selection (C1, C2 and C3) were sown to evaluate the selected mutant and estimate the response to selection. The seeds were sown in three replications. Each one contained three ridges and ten holes with a distance of 30 cm between. In each hole there were two seeds. After two weeks, thinning was made to maintain one plant/hole. The cultural practices were applied as recommended.
Recorded data: At flowering stage: Plant height, No. of leaves, stem diameter, leaf area, fresh weight of vegetative growth, flowering date (days from sowing seeds to showing color of the first inflorescence per plant), No. of inflorescences per plant, number of ray florets per inflorescence and inflorescence diameter were recorded. Chlorophyll content (SPAD unit) was estimated8. Number of seeds per main head and twenty seeds weight were recorded at the maturity stage.
Statistical analysis: Data were statistically analyzed using analysis of variance for Randomized Complete Block Design (RCBD) for separate analysis. Combined analysis for the two years was analyzed using split plot analysis. Comparison among means was done using least significant differences (LSD) at 0.05% and simple correlation coefficient between traits were done9. Realized gain% was estimated from combined means10 as follow:
Realized gain% for C1 = COX 100, for C2 = COX100, for C3 = COX100
RESULTS
Significant differences were observed among populations (parent, base population, C1, C2 and C3 mass selections) in both seasons for all traits except for the number of florets per inflorescence as shown in Table 1.
| Table 1: | Analysis of variance for separate and combined of the two evaluated seasons for all studied traits in the parent, base population and three cycles of mass selection in the branching mutants of Helianthus annuus |
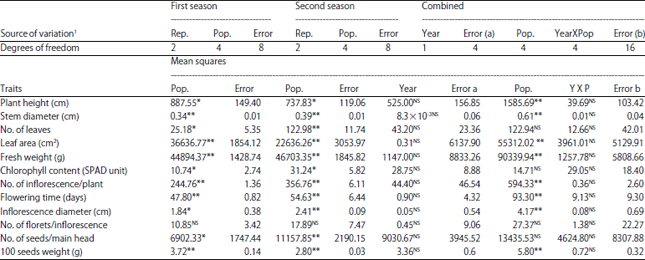 | |
| 1Rep: Replication, Pop: Populations, YXP: Interaction between year and population, NS,*,**: Not significant and significant at p = 0.05 and 0.01, respectively | |
The differences between populations in the combined analysis were significant for all traits except for the No. of leaves, florets per inflorescence and No. of seeds per main head. No significant differences were found for mean square of seasons and the interaction between seasons and populations for all studied traits.
Mean values of the studied traits for the parent, base population and three cycles of mass selection populations are illustrated in Table 2 and 3.
For all vegetative growth traits, a gradual increase was found from the first cycle of mass selection to the second and third one in both evaluation seasons as well as the combined analysis.
Realized gain% for the C1, C2 and C3 of mass selection relative to the base populations was 5.0, 8.1 and 12.6, respectively for plant height, 8.0, 32.0 and 40.0 for stem diameter, 6.6, 10.0 and 32.6, respectively for No. of leaves, 16.4, 54.0 and 121.9 for leaf area and 9.4, 56.0 and 59.8%, respectively for fresh weight.
With respect to chlorophyll content, no significant differences were found in the combined analysis, while significant differences were found in both seasons. Mean values of chlorophyll were decreased in the three cycles of mass selection compared to base population in the first season but an increase was found in the first cycle only in the second season. Response of the third cycle to selection relative to base population was 6%.
Number of inflorescences per plant was increased gradually during cycles of mass selection in both seasons and their combined as shown in Table 3. Mass selection improves No. of inflorescences per plant by 16.3, 19.7 and 61.4%, respectively. It had a negative effect on the earliness of flowering. Mass selection delayed flowering by 5.8, 10.0 and 12.9% as compared to base population. The latter recorded a significant decrease on the inflorescence diameter compared to their parent for both seasons and their combined values. However, mass selection cycles improve this trait with a realized gain 18.3, 30.6 and 40.8% compared to base population.
No significant differences were found in the No. of florets per inflorescence in both seasons and their combined values. Thus, a slightly increase was found. Where, realized gain% that estimated from combined analysis was 5.9, 7.2 and 11.6 for C1, C2 and C3, respectively.
The third cycle of mass selection increased significantly the No. of seeds per main head in the second season. No significant differences were found in the combined analysis. However, a slight increase was found. Response to selection was 17.8, 19.8 and 35.4% in the three cycles from C1-C3, respectively.
Mean values of 100 seeds weight were increased significantly during three cycles ranged from 3.49 g in the C1 to 4.63 g in the C3 of the combined analysis. Responses to selection compared to base population were 49.7, 84.5 and 98.7 for C1, C2 and C3, respectively.
Table 4 presents simple correlation coefficient between the studied traits calculated in the combined analysis for the two evaluation seasons of the three mass selection cycles, base population and the parent.
| Table 2: | Mean performance for separate and combined of the two evaluated seasons for plant height, stem diameter, No. of leaves, leaf area, fresh weight and chlorophyll content in the parent, base population and three cycles of mass selection in the branching mutant of Helianthus annuus |
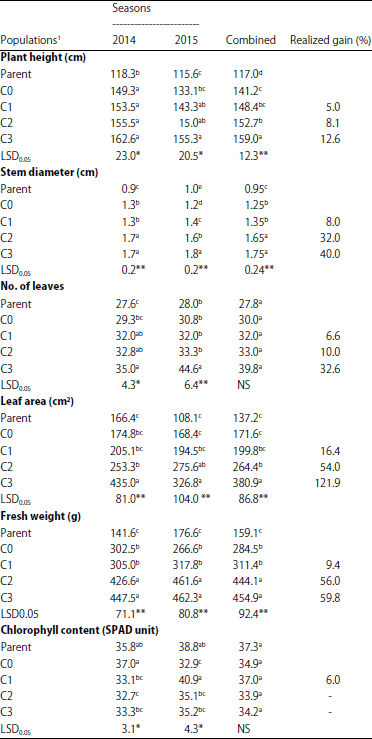 | |
1Parent: cv.Giza102, C0: Base population, C1, C2 and C3: First, second and third cycle of mass selection. Values in the same column not followed by the same letter are significantly different at the 5% level of probability, NS,*,**: Not significant and significant at p = 0.05 and 0.01, respectively | |
Positive and high correlation coefficients were found between No. of inflorescences per plant and all other traits except the No. of seeds per main head where moderate correlation was extend (0.66). The same observation was obtained between stem diameter and all other traits.
| Table 3: | Mean performance for separate and combined of the two evaluated seasons for the No. of inflorescences per plant, flowering date, inflorescence diameter, No. of florets/inflorescence, No. of seeds/main head and 100 seeds weigh (g) in the parent, base population and three cycles of mass selection in the branching mutant of Helianthus annuus |
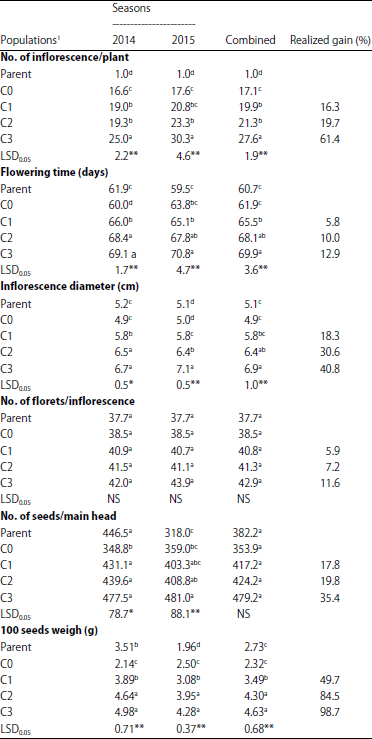 | |
1Parent: cv.Giza102; C0: Base population, C1, C2 and C3: First, second and third cycle of mass selection. Values in the same column not followed by the same letter are significantly different at the 5% level of probability, NS,*,**: Not significant and significant at p = 0.05 and 0.01, respectively | |
Highly and significant correlation were found between inflorescence diameter with each of number of seeds and 100 seeds weight, indicating its importance for the improvement of seed yield.
| Table 4: | Simple correlation coefficient between all studied traits in the combined values of the two evaluated seasons after three cycle of mass selection in the branching mutants of Helianthus annuus |
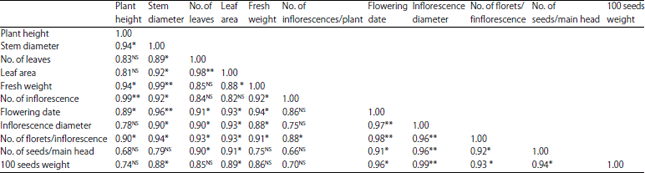 | |
| NS,*,**: Not significant and significant at p = 0.05 and 0.01, respectively | |
DISCUSSION
Through evaluation of the parent, base population, C1, C2 and C3 mass selection cycles in both evaluated seasons and combined analysis of them showed that, seasonal differences were insignificant for all studied traits. These results indicating that, the selected mutant after three cycle of mass selection was stable during seasons.
Mass selection cycles were effective in shifting the mean values towards the positive response. This may be due to selective random mating which might have helped in breaking closely linked genes, gene complexes or eliminates the deleterious recessive alleles which can be produced as a chemical mutagen effect7. These results are in accordance with the results of previous studies10-14.
The significant improvement in mean performance of most studied traits indicated that, these characteristics are under the influence of genetic control. Hence, mass selection was effective in improving the valuable traits by increasing the desired gene frequency and appearance of new favorable genotypes14,15. The effective mass selection tools in improving Helianthus annuus is referred to that these traits are under control of additive genes7. They observed a similar response for seed yield after two cycles of mass selection on Helianthus annuus. The positively and highly correlation coefficients that found between the No. of inflorescences per plant and most other traits, led to the success of the mass selection breeding program in improving the branching mutant, depending on the number of inflorescences per plant and stem diameter.
CONCLUSION
The population obtained after three cycles of mass selection is superior in most studied traits compared to the base population and the parent. This result indicated that mass selection was a successfull method to improve Helianthus annuus. More evaluation seasons in different locations were needed before registration as a new cultivar.
REFERENCES
- Gvozdenovic, S., S. Bado, R. Afza, S. Jocic and C. Mba, 2009. Intervarietal Differences in Response of Sunflower (Helianthus annuus L.) to Different Mutagenic Treatments. In: Induced Plant Mutations in the Genomics Era, Hu, Q.Y. (Ed.). Food and Agriculture Organization of the United Nations, Rome, pp: 358-360.
Direct Link - Mostafa, G.G., 2011. Effect of sodium azide on the growth and variability induction in Helianthus annuus L. Int. J. Plant Breed. Genet., 5: 76-85.
CrossRefDirect Link - Jain, S.M., 2005. Major mutation-assisted plant breeding programs supported by FAO/IAEA. Plant Cell Tissue Org. Cult., 82: 113-123.
CrossRefDirect Link - Natikar, P., K. Madhusudan, U. Kage, H.I. Nadaf and B.N. Motagi, 2013. Genetic variability studies in induced mutants of sunflower (Helianthus annuus L.). Plant Gene Trait, 4: 86-89.
CrossRefDirect Link - Sakin, M.A., A. Yildirim and S. Gokmen, 2004. The evaluation of agronomic traits of durum wheat (Triticum durum Desf.) mutants. Pak. J. Biol. Sci., 7: 571-576.
CrossRefDirect Link - Nambeesan, S.U., J.R. Mandel, J.E. Bowers, L.F. Marek and D. Ebert et al., 2015. Association mapping in sunflower (Helianthus annuus L.) reveals independent control of apical vs. basal branching. BMC Plant Biol., Vol. 15.
CrossRefDirect Link - Gowda, J. and A. Seetharam, 2008. Response to mass and SI selection for autogamy, seed yield and oil content in sunflower populations (Helianthus annuus L.). Helia, 31: 101-110.
CrossRefDirect Link - Yadawa, U.L., 1986. A rapid and nondestructive method to determine chlorophyll in intact leaves. HortScience, 21: 1449-1450.
Direct Link - Bakheit, B.R. and M.A. Ali, 2013. Improvement of single cut Egyptian clover (Trifolium alexandrinum L.) by mass and family selection methods. Asian J. Crop Sci., 5: 304-311.
CrossRefDirect Link - Bakheit, B.R., M.A. Ali and A.A. Helmy, 2011. Effect of selection for crown diameter on forage yield and quality components in alfalfa (Medicago sativa L.). Asian J. Crop Sci., 3: 68-76.
CrossRefDirect Link - Hassan, M.S. and S.H.M. Abd-El-Haleem, 2014. Effectivenss of gamma rays to induce genetic variability to improve some agronomic traits of canola (Brassica napus L.). Asian J. Crop Sci., 6: 123-132.
CrossRefDirect Link - Solieman, T.H.I. and E.I.M. Ragheb, 2014. Two selection methods and estimation of some important genetic parameters in broad bean (Vicia faba L.). Asian J. Crop Sci., 6: 38-48.
CrossRefDirect Link - Rashwan, A.M.A., 2015. Improvement of weight fruit and yield in super strain-B cultivar of tomato (Lycopersicon esculentum Mill) by mass selection. J. Am. Sci., 11: 45-50.
Direct Link