
J.G. Nyaanga
Department of Crops, Horticulture and Soils, Egerton University, P.O. Box 536, Egerton, Kenya
A.W. Kamau
Department of Crops, Horticulture and Soils, Egerton University, P.O. Box 536, Egerton, Kenya
R.S. Pathak
Department of Crops, Horticulture and Soils, Egerton University, P.O. Box 536, Egerton, Kenya
R.K. Tuey
Kenya Agricultural Research Institute, National Plant Breeding Centre, Private Bag, Njoro, Kenya
Asian Journal of Crop Science
Year: 2014 | Volume: 6 | Issue: 3 | Page No.: 281-288







ABSTRACT
Biological control and host plant resistance are key components of Integrated Pest Management (IPM). Little studies have however been done on their potential for integration in the management of cereal aphids in wheat. The impact of host plant resistance and coccinellid predators on cereal aphid population and wheat yield was therefore studied in outdoor cages. Two wheat varieties and two coccinellids predators Adonia variegeta and Chelomenes lunata were compared against a control. A mixed population of three cereal aphid species was inoculated onto wheat plants and newly emerged coccinellids were introduced in all plots except controls. Aphid counts were taken at five wheat growth stages. Repeated measure analysis indicated that there were significant differences in aphid counts between the cages with coccinellids and the control (F = 7.20; df 2, 12; MSE = 28; p<0.09) in 2008 and (F = 9.4; df 2, 12; MSE = 19; p<0.09) in 2009. The aphid numbers in the control (13.7) were approximately twice those in cages with beetles (7). Coccinellids and the resistant wheat variety showed a positive interaction at flowering stage (F = 25.3; df 60, 89; MSE = 3.9; p<0.01). Cages with coccinellids recorded more than 50% increase in wheat yield (F = 44; df 10, 17; MSE = 0.00004; p<0.001) in 2008 and (F = 57; df 10, 17; MSE = 0.000036; p<0.0001) in 2009 compared to the control. Wheat plant resistance and biological control could therefore be complementary strategies in IPM against cereal aphids.
PDF Abstract XML References Citation
Received: December 18, 2013;
Accepted: February 27, 2014;
Published: May 15, 2014
How to cite this article
J.G. Nyaanga, A.W. Kamau, R.S. Pathak and R.K. Tuey, 2014. Impact of Host Plant Resistance and Coccinellid Predators on Cereal Aphid Abundance and Wheat Yield. Asian Journal of Crop Science, 6: 281-288.
DOI: 10.3923/ajcs.2014.281.288
URL: https://scialert.net/abstract/?doi=ajcs.2014.281.288
DOI: 10.3923/ajcs.2014.281.288
URL: https://scialert.net/abstract/?doi=ajcs.2014.281.288
INTRODUCTION
Host-plant resistance and biological control are important components of Integrated Pest Management (IPM) (Mallah et al., 2001) and could act additively to suppress populations of agricultural pests (Biswas and Sing, 1998; Chacon et al., 2012). Plant breeders have however, for along time almost exclusively focused on selecting varieties with enhanced direct defenses against pests while biological control workers have mainly concentrated on improving natural enemy traits (Cortesero et al., 2000). There is an urgent need for a deliberate attempt to bridge these two pest management practices.
Cereal aphids are considered the most serious pests of wheat causing both direct and indirect damage (Curtis et al., 2002). Strategies for managing cereal aphids have over the years tended to focus on short-term single-technology interventions with over-reliance on synthetic insecticides. The development of Biological control and host plant resistance has taken little account of their potential for integration in the management of cereal aphids in wheat. Messina and Sorenson (2001) observed that even modest reductions in aphid population growth as might occur on tolerant-resistant plants can be sufficient to produce a synergistic level of pest suppression when combined with lacewing larvae predators. Soybean cultivars with moderately or densely pubescent trichomes showed positive influences on the abundance of some herbivores and their predators on Soybean (Dai et al., 2010). McCarville and O’Neal (2012) also demonstrated high compatibility of biological control with the soybean aphid plant resistance by reporting a 63% reduction in the Soybean aphid population growth rate using the combined effect of natural enemies and pyramided resistance.
Significant reductions in population growth rate and subsequent density of aphids have been reported by Fuentes-Contreras and Niemeyer (2000) when integrating a partial resistance wheat cultivar with the parasitoid Aphidius rhopalosiphi and the entomopathogenic fungus Pandora neoaphidi. The current study hypothesized that the joint action of coccinellids and partial wheat resistance can reduce the population of cereal aphids on wheat to less damaging levels and improve wheat yield. An experiment was therefore designed to test this hypothesis.
MATERIALS AND METHODS
The research was carried out in 2008 and 2009 at Egerton University situated in the Rift valley province of Kenya. A two factor experiment consisting two commercial wheat varieties, two coccinellid beetle species and a control (no beetles), was set in outdoor cages in a factorial Randomised Complete Block Design (RCBD) replicated three times. The wheat varieties included an aphid susceptible variety (Duma) and a partially resistant variety (Fahari). The coccinellid species used were Adonia variegata (Goeze) and Chelomenes lunata (Fabricius). Experimental plots measuring 1.5 mx2 m with an inter-row spacing of 20 cm were planted according to recommendations for commercial wheat production. A screen mesh enclosure (cage) was erected around each plot immediately after planting to exclude any aphids or predators flying in or out of the plots. The cages were made from wooden frames with polyester screen covers of mesh size 0.4 by 0.5 mM and thread thickness of 0.1 mM on the sides and fine wire mesh size 0.5 by 0.5 mM at the top.
Methodology: Twenty fourth instar cereal aphids (total sixty) from each of the three species; Ropalosiphum padi (Linnaeus), Metapolophium dirhodum (Walker) and Sitobion avenae (Fabricius) were collected from greenhouse cultures and randomly inoculated onto the wheat plants at Zadock growth stage 12 (2-leaf stage) in all the caged plots. The aphids were allowed to establish on the plants for two weeks (two generations) before two female coocinellid beetles with a maximum age difference of 24 h since eclosion were evenly distributed in all plots except controls. One adult male beetle was also placed in the plots with female beetles after 2 h.
Aphid counts were taken from ten plants and their tillers randomly selected in each plot at five growth stages namely; GS 22 (2-tiller stage), GS 32 (stem elongation stage) GS 45 (booting stage) 69 (flowering stage) GS 91 (ripening stage) (Zadoks et al., 1974). Wheat yield was taken from all plants and tillers within an area measuring 1m sq within the central rows of the plot.
The data on aphid counts was first transformed using the v (x+1) to homogenize and stabilize the variance before being subjected to analysis. Between and among treatment variation in aphid counts was analyzed using a repeated-measures analysis of variance (ANOVA) with sampling date (growth stages) as the repeated measure. The effect of time and the interaction of treatment by time were included within the ANOVA to test for differences in aphid counts and treatment differences across sample dates (SAS, 2002).
RESULTS
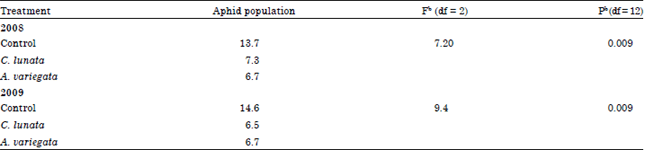
The aphid population in 2009 was generally but not significantly higher compared to 2008. Between treatments comparisons showed that there were significant differences in aphid counts between the cages which had no beetles (control) and those with beetles across the sampling dates in both 2008 (F = 7.20; df 2, 12; MSE = 28; p<0.09) and 2009 (F = 9.4; df 2, 12; MSE = 19; p<0.09) (Table 1). The aphid counts in the control cages (13.7) in 2008 was approximately twice the population in the cages whereby C. lunata recorded 7.3 and A. variegata 6.7 aphids. A similar trend was observed in 2009 whereby the aphid counts in control cages were 14.6 while the ones with beetles had 6.5 and 6.7 aphids for C. lunata and A. variegata, respectively. No significant differences in aphid counts was detected when comparing populations in the cages planted with Fahari or Duma wheat varieties in both years (F = 3.28; df 2, 12; MSE = 28; p<0.009) and (F = 1.09; df 2, 12; MSE = 19; p<0.009) (Table 2).
Among-treatment comparisons revealed that time had a significant influence on aphid counts and hence was the main contributing factor that brought about treatment differences. There was a positive interaction between the two coccinellids species and Fahari wheat variety at flowering wheat growth stage in both 2008 and 2009 (F = 25.3; df 60, 89; MSE = 3.9; p<0.01) (Fig. 1). The aphid population was below 10 aphids per plant in the cages planted with Fahari compared to more than 15 aphids per plant observed in the cages with Duma wheat variety (Fig. 2) irrespective of the interacting coccinellid species in 2009 the period which aphid population was generally higher.
The cages with beetles recorded a significantly higher wheat yield (F = 44; df 10, 17; MSE = 0.00004; p<0.001) in 2008 and F = 57; df 10, 17; MSE = 0.000036; p<0.0001) in 2009 compared to the cages that had no beetles. There was a more than 50% increase in wheat yield over the control for both wheat varieties (Fig. 2a, b) during the two seasons studied. Fahari recorded a significantly higher yield (0.097 and 0.101) in 2008 and 2009, respectively in the control plots compared to Duma which recorded 0.086 and 0.093 in 2008 and 2009, respectively.
| Table 1: | Effect of coccinellids on cereal aphid population in 2008 and 2009 |
 | |
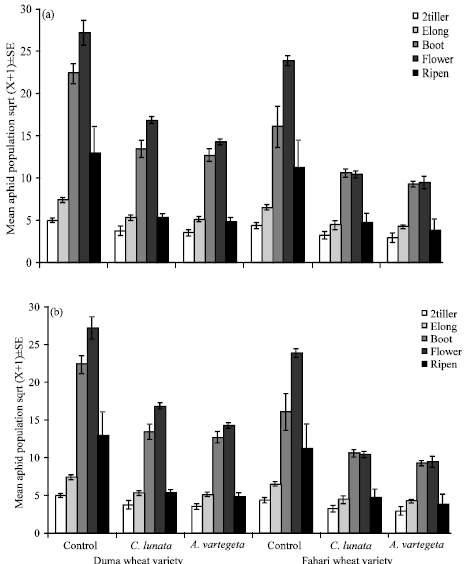 | |
| Fig. 1(a-b): | Mean aphid populations±SE with coccinellids (control, C. lunata and A. variegeta) at four growth stages (2-tiller, elongation, booting, flowering and ripening) of wheat variety Duma and Fahari in (a) 2008 and (b) 2009 aphid population was generally higher |
| Table 2: | Effect of wheat variety on cereal aphid population in 2008 and 2009 |
 | |
| a: Means were pooled across all sample dates (Growth stages), b: Values derived from a two-way ANOVA | |
There was however no significant difference in yield between Fahari and Duma with either of the coccinellid species tested in both years.
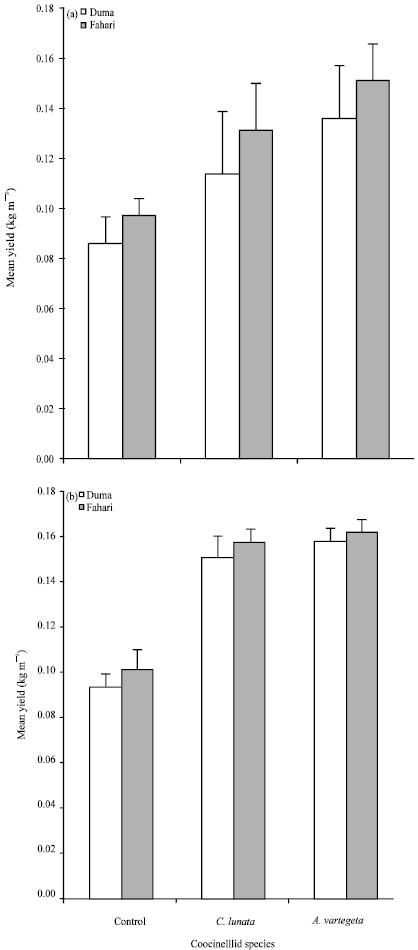 | |
| Fig. 2(a-b): | Effect of coccinellids and varieties on wheat yield in (a) 2008 and (b) 2009, respectively |
DISCUSSION
Based on the results obtained in this study, the two coccinellid beetles A. variegata and C. lunata can be considered efficient predators of cereal aphids and could contribute significantly to the implementation of IPM of these pests in wheat. This is consistent with studied by Nyaanga et al. (2012) and Dehkordi et al. (2013) who demonstrated that the coccinellid species H. variegata was an effective predator and hence a good biocontrol agent in the management of cereal aphids in wheat and cotton aphid (Aphis gossypii), respectively.
The significantly lower aphid numbers recorded on Fahari wheat variety is an indication of some level of compatibility between the partially resistant wheat variety (Nyaanga et al., 2006) and the two coccinellid species. Plant mediated factors that increase the efficiency of natural enemies on their prey have been extensively studied and reported. The plant traits that have effects on natural enemies of cereal aphids have been described by Brewer and Elliot (2004) as those that are innate, bred, or induced by aphid feeding. For instance, tritrophic effects of plant chemistry are important (Ode, 2006) in assessing the degree of compatibility between biological control and plant resistance approaches to pest control. Plants often release a blend of volatile organic compounds in response to damage by herbivorous insects that may serve as cues to locate those herbivores by natural enemies (De Boer et al., 2008; Dicke, 2009; Hare, 2011).
Significant reductions in aphid population were recorded during flowering wheat stage, a period when the control plots recorded the highest aphid population. This could be an indication of both functional and numerical response of the coccinellid predators to their aphid prey density. There is a correlation between aphid numbers and the abundance of coccinelid life stages (egg, larva and beetle) on host plants (Hossain et al., 2001; Butin et al., 2003; Dixon et al., 2005; Jogender, 2006; Mrosso et al., 2013). Aphid and coccinellid apperarance has also been observed to be highly dependent on the phenological stage of the host plant (Jarosik et al., 2003; Akhtar and Khaliq, 2003). The high presence of immature (larval) stages during flowering which is a critical wheat growth stage (Nyaanga et al., 2009) contributes most towards the suppression of aphid densities. This is because most of the coccinellids feeding takes place during their juvenile stages (Ruppert et al., 2003; Shah and Khan, 2013).
The higher wheat yield recorded from Fahari wheat variety may have been as a result of the lower aphid numbers recorded on this variety during flowering stage. Cereal aphids can cause serious yield losses particularly if the infestation occurs at heading. The results from this study suggest that even modest reductions in aphid population growth (as might occur on partially resistant wheat variety) can be sufficient to produce a synergistic level of pest suppression with biocontrol agents like coccinellids.
CONCLUSION
Planting of wheat varieties that have shown some resistance to aphids is important since even modest reductions in aphid population can be sufficient to produce synergistic effects when combined with biological control agents. The study emphasizes the necessity of bridging plant breeding and biological control practices to improve crop protection. Plant breeders should not only focus on selecting varieties with enhanced direct defenses against pests but also place special emphasis on breeding crop plants with natural enemy enhancing traits.
ACKNOWLEDGMENTS
We thank Egerton University and African Institute for Capacity Development (AICAD) for the financial support of this research.
REFERENCES
- Chacon, J.M., M.K. Asplen and G.E. Heimpel, 2012. Combined effects of host-plant resistance and intraguild predation on the soybean aphid parasitoid Binodoxys communis in the field. Biol. Control, 60: 16-25.
CrossRef - Mallah, G.H., A.K. Korejo, A.R. Soomro and A.W. Soomro, 2001. Population dynamics of predatory insects and biological control of cotton pests in Pakistan. J. Biol. Sci., 1: 245-248.
CrossRefDirect Link - Cortesero, A.M., J.O. Stapel and W.J. Lewis, 2000. Understanding and manipulating plant attributes to enhance biological control. Biol. Control, 17: 35-49.
CrossRef - Brewer, M.J. and N.C. Elliott, 2004. Biological control of cereal aphids in North America and mediating effects of host plant and habitat manipulations. Annu. Rev. Entomol., 49: 219-242.
CrossRef - Butin, E., J. Elkinton, N. Havill and M. Montgomery, 2003. Comparison of numerical response and predation effects of two coccinellid species on hemlock woolly adelgid (Homoptera: Adelgidae). J. Econ. Entomol., 96: 763-767.
CrossRefDirect Link - Fuentes-Contreras, E. and H.M. Niemeyer, 2000. Effect of wheat resistance, the parasitoid Aphidius rhopalosiphi and the entomopathogenic fungus Pandora neoaphidis, on the population dynamics of the cereal aphid Sitobion avenae. Entomol. Exp. Appl., 97: 109-114.
CrossRef - Biswas, S. and R. Singh, 1998. Interaction between host plant resistance and the biocontrol of a cereal aphid: A laboratory study. Biol. Agric. Hortic., 16: 25-36.
CrossRef - Dai, H., Y. Wang, Y. Du and J. Ding, 2010. Effects of plant trichomes on herbivores and predators on soybeans. Insect Sci., 17: 406-413.
CrossRef - De Boer, J.G., C.A. Hordijk, M.A. Posthumus and M.A. Dicke, 2008. Prey and non-prey arthropods sharing a host plant: Effects on induced volatile emission and predator attraction. J. Chem. Ecol., 34: 281-290.
CrossRef - Dicke, M., 2009. Behavioural and community ecology of plants that cry for help. Plant Cell Environ., 32: 654-665.
CrossRef - Dixon, A.F.G., V. Jarosik and A. Honek, 2005. Thermal requirements for development and resource partitioning in aphidophagous guilds. Eur. J. Entomol., 102: 407-411.
Direct Link - Hare, J.D., 2011. Ecological role of volatiles produced by plants in response to damage by herbivorous insects. An. Rev. Entomol., 56: 161-180.
CrossRef - Jarosik, V.A., A. Honek and A. Tichopad, 2003. Comparison of field population growths of three cereal aphid species on winter wheat. Plant Prot. Sci., 39: 61-64.
Direct Link - Hossain, M.A., M.A. Ahad, H.F. El-Taj, M.R. Amin and A.K.M. Azad, 2001. Prevalence and relative abundance of coccinellid beetles on mustard (Brassica campestris) crop. Pak. J. Biol. Sci., 4: 136-138.
CrossRefDirect Link - Akhtar, I.H. and A. Khaliq, 2003. Impact of plant phenology and coccinellid predators on the population dynamic of rose aphid Macrosiphum rosaeiformis das (Aphididae: Homoptera) on rose. Asian J. Plant Sci., 2: 119-122.
CrossRefDirect Link - Messina, F.J. and S.M Sorenson, 2001. Effectiveness of lacewing larvae in reducing russian wheat aphid populations on susceptible and resistant wheat. Biol. Control, 21: 19-26.
CrossRef - Mrosso, F., M. Mwatawala and G. Rwegasira, 2013. Functional responses of Cheilomenes propingua, C. lunata and C. sulphurea (Coleoptera: Coccinellidae) to predation on Aphis gossypii (Homoptera: Aphididae) in Eastern Tanzania. J. Entomol., 10: 76-85.
CrossRefDirect Link - Nyaanga, J.G., A.W. Kamau and J.K. Wanjama, 2006. Niche and temporal differences in colonization of wheat varieties by three species of cereal aphids. Asian J. Plant Sci., 5: 713-716.
CrossRefDirect Link - Nyaanga, J.G., A.W. Kamau, R.S. Pathak and R.K. Tuey, 2012. The effect of different cereal aphid species on the performance of two coccinellid predators. J. Entomol., 9: 41-49.
CrossRef - Ode, P.J., 2006. Plant chemistry and natural enemy fitness: Effects on herbivore and natural enemy iteractions. Ann. Rev. Entomol., 51: 163-185.
CrossRef - Dehkordi, S.D., A. Sahragard and J. Hajizadeh, 2013. The effect of prey density on life table parameters of Hippodamia variegate (Coleoptera: Coccinellidae) fed on Aphis gossypii (Hemiptera: Aphididae) under laboratory conditions. ISRN Entomol., (In Press).
CrossRef - Zadoks, J.C., T.T. Chang and C.F. Konzak, 1974. A decimal code for growth stages of cereals. Weed Res., 14: 415-421.
CrossRefDirect Link