
Mohamed F. El-Nady
Department of Agricultural Botany (Branch: Agricultural Botany), Faculty of Agriculture, Kafrelsheikh University, 33516, Kafr El-Sheikh, Egypt
Elsayed B. Belal
Department of Agricultural Botany (Branch: Agricultural Microbiology), Faculty of Agriculture, Kafrelsheikh Univ., 33516, Kafr El-Sheikh, Egypt
Asian Journal of Crop Science
Year: 2013 | Volume: 5 | Issue: 3 | Page No.: 222-237







ABSTRACT
The aim of the present study was to investigate the influence of Pendimethalin (PM) on growth and anatomical characteristics of Cucumis sativus and Echinochloa crus-galli plants. Moreover, Psudomonas putida and compost were evaluated for detoxification of PM in soil. Seeds were sown in pots containing either PM treated soil or PM and P. putida or compost treated soil. Twenty eight days later, the phytotoxicity bioassay was performed on the growing plants. PM significantly decreased germination and increased seedling mortality percentages. Cotyledonary leaf and hypocotyls and true leaves of C. sativus seedling treated with PM seemed to be dark green colors, swelling and brittleness. In addition, reduction in fresh and dry weights of the treated plants was observed. In contrary, chlorophyll contents were significantly increased. Reducing in number of xylem vessels was found not only in hypocotyl but also in the first true leaf of C. sativus plants. Insignificant differences were observed in the most growth and anatomical parameters between P. putida or compost applying in PM contaminated soil comparing with PM treated plants. P. putida and compost were effective in PM degradation in soil with half-life of 4.67 and 5 days, respectively. PM half-life was 51.9 days in untreated clay soil. The results suggested that bioremediation by P. putida and compost was proved as an effective method for detoxification of pendimethalin in soil.
PDF Abstract XML References Citation
Received: November 12, 2012;
Accepted: February 20, 2013;
Published: May 18, 2013
How to cite this article
Mohamed F. El-Nady and Elsayed B. Belal, 2013. Effect of Phytotoxicity of Pendimethalin Residues and its Bioremediation on Growth and Anatomical Characteristics of Cucumis sativus and Echinochloa crus-galli Plants. Asian Journal of Crop Science, 5: 222-237.
DOI: 10.3923/ajcs.2013.222.237
URL: https://scialert.net/abstract/?doi=ajcs.2013.222.237
DOI: 10.3923/ajcs.2013.222.237
URL: https://scialert.net/abstract/?doi=ajcs.2013.222.237
INTRODUCTION
Pendimethalin (N-(1-ethylpropyl)-2, 6-dinitro-3,-4-xylidine) has the empirical formula C13H19N3O4, a selective pre-emergent herbicide a dinitroaniline group, is used extensively for weed control in cotton, rice, soybean and tobacco (Smith et al., 1995). It has been reported that the use of pendimethalin (PM) before crop emergence or planting resulted in the production of stunting plant portions due to root and shoot growth inhibition. The reason of such inhibition was due to the alteration of cell division steps needed for chromosome separation and cell wall formation (Parka and Soper, 1977; Appleby and Valverde, 1988). Studies in terrestrial ecosystems showed that 10-20% of the herbicide vaporizes within the first week or two after application. The observation of phytotoxicity to crops and weeds 200 days after application confirms that the dissipation time of PM is high enough to harm plants far beyond the period during which it is intended to be active (Strandberg and Scott-Fordsmand, 2004). PM has been classified by the US Environmental Protection Agency (EPA) as a Persistent Bioaccumulative Toxic (PBT) (Kamrin, 1997; Megadi et al., 2010).
In order to protect rotation crops from injury, the use of PM should be minimized. As microorganisms can mineralize and detoxify pesticides and use them for their growth, bioremediation should be considered for accelerating the rate of elimination of pesticide from contaminated water and soil. Meanwhile, successful microorganisms used for bioremediation should have high degradation ability and should be stable under varied conditions, such as changes in pH and temperature. Therefore, it is necessary to investigate the effects of various environmental factors on the growth ability of the tested microorganisms (Pattanasupong et al., 2004). On the other hand, it was found that various materials were used as soil amendments, nutrients, to increase and enhance the degradation potential of xenobiotics such as Yard manure compost (Guo et al., 1991; Leoni et al., 1992; Liu et al., 1995; Gan et al., 1996; Zheng and Cooper, 1996; Grigg et al., 1997; Vidali, 2001) biogas slurry and compost (Belal et al., 2008). However, after remediation toxicity assessments are needed. Firstly, is providing valuable and complementary information to compound analysis. Secondly, the major advantage of toxicity tests is the direct assessment of the potential hazard to the environmental system by both original pollutants and its metabolites (Ahtiainen et al., 2000). Therefore, this study attempted to evaluate Psudomonas putida and compost in remediation of PM contaminated soil. In addition to confirm the complete detoxification of pendimethalin by measuring the toxicity of the treated soil in the presence of bacterial strain or compost against sensitive target weed (Echinochloa crus-galli) and non-target plant (Cucumis sativus).
MATERIALS AND METHODS
Chemicals: Pendimethalin (N-(1-ethylpropyl)-2, 6-dinitro-3,4-xylidine) standard was obtained from Ehrenstorfer (Germany). All other chemicals were of analytical grade.
Microbial degradation of the pendimethalin
Media: M9-Minimal medium as Mineral Salt Liquid (MSL) and Luria Bertani (LB) a complete medium were used through this study as described by Sambrook et al. (1989).
Source and identification of microorganism: Pseudomonas putida (E1) was isolated and identified in previous study (Belal and El-Nady, 2012).
Bioremediation of PM contaminated soil and phytotoxicity test: Pot experiments were conducted at the greenhouse of the Agriculture Botany Department, Faculty of Agriculture, Kafrelsheikh University, Egypt. The phytotoxicity bioassay of (PM) was performed on the contaminated soil after 28 days treatment with P. putida (E1) and compost. Cucumber plants Cucumis sativus L., cultivar Hisham and barnyardgrass Echinochloa crus-galli (Obtained from Rice Weeds Research Department, Rice Research and Training Centre, Sakah, Kafrelsheikh) were used as the test organisms. The phytotoxicity was determined as deformation in morph-physiological and histological measurements comparing to the treatments with P. putida (E1) and compost. All treatments were compared with control treatment (untreated soil).
Clay soil had no previous history of PM concentration was collected from top 12-15 cm randomly following standard procedure and sieved through 2 mm size sieve (Gupta, 2000). The experiments were conducted in 1000 g capacity pots (polyethylene pots, 20 cm inner diameter and 30 cm in depth), each having 1000 g dried clay soil. Soil was contaminated with PM (100 μg g-1 soil) at 2% moisture level in their respective treatment pot before one week from cucumber sowing.
P. putida (E1) was cultured onto mannitol salt agar (MSA)+pendimethalin for 7 days and then the growing colonies were washed with 3 mL sterilized Mineral Salt Liquid (MSL) medium. One hundred mL from cell suspension (107 CFU mL-1 for bacterial strains) was then used to inoculate 1 kg clay containing (100 μg g-1) from pendimethalin before one week from cucumber sowing, mixed well and kept under incubation for 28 days at temperature 30±2°C.
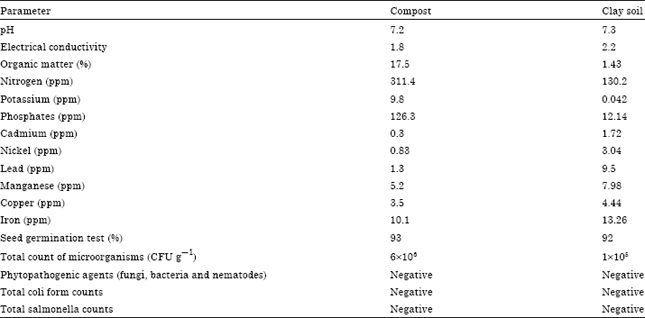
Compost was used as soil amendments. The calculated quantity i.e. 100 g of compost was applied before sowing of trial in respective treatment pot, mixed well and kept under incubation for 28 days at 30±2°C (Belal et al., 2008). Physicochemical characteristics of used clay soil and compost are presented in Table 1.
The soil used in this experiment was fertilized with nitrogen at rate a 360 kg ha-1 of urea fertilizer (contain 46% nitrogen). Super phosphate fertilizer (phosphorus 15%) was added at a rate of 240 kg ha-1 before planting. Potassium was not added because the Egyptian soil is rich in this element.
Five cucumber seeds of (Cucumis sativus L., cultivar Hisham) and barnyardgrass (Echinochloa crus-galli) were sown on 1st of October 2010 in each pot after one week from soil contamination with pendimethalin.
The residue half-live (RL50) for PM residues was calculated using the equation of Moye et al. (1987). Control pots of equal weight of soil and pesticide without any microbial population or compost were run in parallel at all intervals to assess a biotic losses as well as measuring of the botanical parameters on the tested plants.
| Table 1: | Physico-chemical characteristics of clay soil and compost (Belal and El-Nady, 2012) |
 | |
Growth characters and chlorophyll pigment determination: Germination percentage was determined at 15th day from sowing. Percentages of seedling mortality were calculated as percentages of total number of germinated seeds. For seedlings characters, samples were taken at 15 days from sowing to estimate seedling fresh and dry weights (dried in an electric oven at 70°C for 72 h) g plant-1. Chlorophyll a, b and total were determined in cotyledonary leaf and the first true leaf of cucumber (C. sativus) plants and in the second leaf of barnyardgrass (E. crus-galli) using spectrophotometer method as described by Moran and Porath (1980).
Histological parameters calculation: C. sativus hypocotyl specimens were taken from the middle region. The leaf specimens including the midrib were taken from the first true leaf. For E. crus-galli, leaf specimens including the midrib were taken from the second leaf from the plant tip. Specimens were taken on day 15th of treating. Specimens were taken on 10th day of sowing. Specimens were fixed in formalin alcohol acetic acid mixture (FAA, 1: 18: 1; v/v), washed and dehydrated in alcohol series. The dehydrated specimens were infiltrated and embedded in paraffin wax (52-54°C m.p.). The embedded specimens were sectioned using a rotary microtome (Leica RM 2125) at a thickness of 8-10 μm. Sections were mounted on slides and deparaffinized. Staining was accomplished with safranine and azur II (Gutmann, 1995) cleared in xylol and mounted in Canada balsam (Ruzin, 1999). Ten reading from 3 slides were examined with electric microscope (Leica DM LS) with digital camera (Lieca DC 300) and then photographed. The histological feature of the hypocotyl was thickness of hypocotyl, vascular and cortex tissues as well as number of vessels/bundle. Moreover, the histological features of the first true leaf were thickness of lamina, midrib region, midrib vascular bundle, mesophyll (palisade and spongy tissues) and vascular tissues (xylem and phloem) in addition to the No. of vessels/midrib vascular bundle. Also, leaf lamina thickness of E. crus-galli was calculated. The histological manifestation was calculated using Lieca IM 1000 image manager software. Lieca software was calibrated using 1 cm stage micrometer scaled at 100 μm increment (Leitz Wetzler, Germany 604364) at 4 and 10 X magnifications.
Statistical analysis: Data were subjected to statistical analysis of variance according to Gomez and Gomez (1984).
Analytical procedure: Extraction and determination of PM residues was carried out by the described method by Jazwa et al. (2009) at Central Agric. Pesticides Laboratory, Agricultural Research Center, Ministry of Agriculture and Land Reclamation, Egypt. PM residue in soil was monitored weekly after application date. At each sampling time four soil samples were taken from randomly selected pots of cucumber plants. At the end of that test, PM residues were determined. Soil samples were air-dried, ground and stored at room temperature prior to analysis but no more than three days. Subsamples (20 g) were extracted by shaking for one hour with 100 mL of dichloromethane-acetone mixture (9:1 v:v) on a rotary shaker. The extract obtained, was decanted by a layer of anhydrous sodium sulphate and the soil was rinsed two times with 10 mL of dichloromethane (Luke et al., 1975, 1981; Ambrus et al., 1981). The extract was cleaned using florisil (Ahtiainen et al., 2000). The analysis of the extract was performed using a Hewlett Packard 5890A Gas Chromatograph, equipped with Nitrogen-phosphorus Detector (GC-NPD). The column used in this study was an HP fused-silica capillary column coated with cross-linked methyl silicone (length 25 m, ID 0.31, film thickness 0.52 μm). Nitrogen was used as both the carrier and make-up gas at a flow rate of 30 mL min-1. Hydrogen was used at a flow rate of 3.5 mL min-1 and air at 120 mL min-1. The oven temperature was programmed as follows: initial temperature 150°C (1 min), rate of 10°C min-1 and final temperature 250°C. Recovery studies were carried out regularly by spiking analytical samples with stock solution of pendimethalin standard.
RESULTS AND DISCUSSION
Effects of pendimethalin (PM) contaminated soil on growth and histological parameters of C. sativus and E. crus-galli were studied. Investigated plant growth parameters were seed germination, fresh and dry weight and chlorophyll pigments contents. The detailed histological examinations included hypocotyl and the first true leaf in C. sativus and leaf thickness in E. crus-galli. In addition, P. putida and compost were evaluated for detoxification of PM in soil.
Phytotoxicity assessment: The effect of the remaining toxicity of PM in clay soil on germination, growth and anatomical characters of C. sativus and E. crus-galli plants was estimated after it treated with compost and P. putida (E1) as follow:
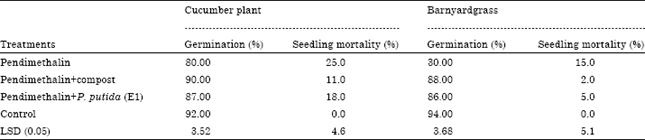
Germination characters: The results in Table 2 showed the influence of the remaining toxicity of PM in clay soil on germination and seedling mortality percentage of the two tested plants after it treated with compost and P. putida (E1). PM caused the highest value in reduction germination and increasing seedling mortality percentage comparing with control treatment. These parameters were improved with compost and P. putida (E1) treatment comparing with PM treatment. Compost treatment was more effective in reduction of seedling mortality comparing with P. putida (E1) treatment. The efficacy of compost and P. putida (E1) was similar in increasing of germination percentage of the tested plants comparing with PM treatment. All treatments were compared with unweedded treatment (control) after 15 days from sowing. Germinating plants absorb PM through epidermis of root, coleoptile or young shoots which come into close contact with the herbicide. Similarly, to other dinitroanilines, PM inhibit the formation of mitotic apparatus microtubules thus affecting chromosome movement and inducing formation of polyploid cells (Tarkowska et al., 1994). They stated that PM acts on fragmoplast causing formation of multinuclear and also on cortical microtubules causing isodiametric growth of cells and disturbance in secondary cell wall. These lead to the formation of swelling on root tips and at the base of the stem (Smeda and Vaughn, 1997). This may lead the inhibition of seed germination and increasing seedling mortality percentages.
Growth characters: Data presented in Table 3 and Fig. 1 illustrate the effect of the remaining toxicity of PM in clay soil on fresh and dry weight of C. sativus and E. crus-galli seedlings after it treated with compost and P. putida (E1).
| Table 2: | Effect of the remaining toxicity of pendimethalin in clay soil on cucumber (C. sativus) and barnyardgrass (E. crus-galli) seed germination and seedling mortality percentage after it was treated with compost and P. putida (E1) |
 | |
 | |
| Fig. 1(a-b): | Seedling 15 days after sowing of (a): C. sativus and (b): E. crus-galli, 1: Control (untreated), 2: PM+compost, 3: PM+P. putida (E1), 4: PM, Hypocotyl (H), CL: Cotyledonary leaf, P: Plumule, FTL: First true leaf, Bar = 1.5 and 3 cm |
| Table 3: | Effect of the remaining toxicity of PM in clay soil on hypocotyl length, fresh and dry weight of C. sativus and fresh and dry weight of E. crus-galli seedlings after it was treated with compost and P. putida (E1) |
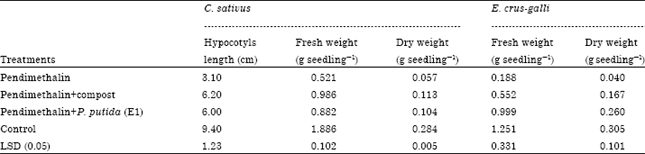 | |
Reduction in C. sativus hypocotyl length values were found in PM treatment. Fresh and dry weight/plant values were reduced in case of treatment with PM comparing with the other treatments. These parameters were increased with compost and followed by P. putida (E1). The control (without PM) treatment recorded the highest value for the measured plant parameters comparing with the other treatments. PM treatment reduced the measured botanical parameters more than the other treatments and this due to PM residues in soil which were 95, 30 and 25% with PM, compost and P. putida (E1) treatments, respectively. These plant parameters were improved gradually when PM residues were disappeared.
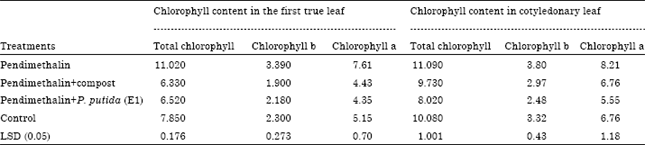
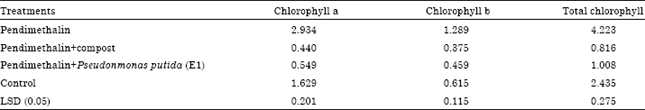
Chlorophyll pigments: Application of PM significantly increased chlorophyll pigment (chlorophyll a, chlorophyll b and total contents of chlorophyll) in cotyledonary and the first true leaf of C. sativus seedling compared with the other treatments (Table 4). Chlorophyll pigment contents in cotyledonary leaf were higher than in the first true leaf. Similar results were recorded in E. crus-galli (Table 5). It was interesting to note that, the increase in chlorophyll pigment contents was accompanied with the increase in mesophyll tissue thickness.
| Table 4: | Effect of the remaining toxicity of pendimethalin in clay soil on chlorophyll pigment (a, b and total chlorophyll) of cotyledonary and the first true leaves of C. sativus seedlings after it was treated with compost and P. putida (E1) |
 | |
| Table 5: | Effect of the remaining toxicity of pendimethalin in clay soil on chlorophyll pigment (a, b and total chlorophyll) in the second leaf of E. crus-galli plants after it was treated with compost and P. putida (E1) 15 days from sowing |
 | |
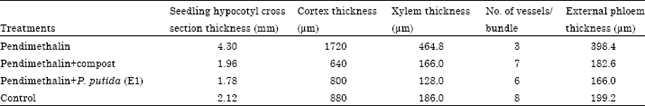
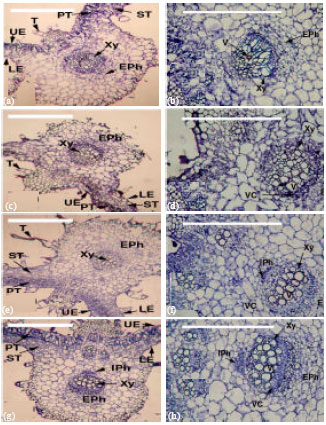
Anatomical characters: The hypocotyls internal structure of cucumber was similar to stems of dicotyledon plants. The hypocotyl structure of cucumber plants as seen in transverse sections consists of the epidermis, ground tissue and vascular system (Fig. 2). The regions between the bundles were parenchymatous. The vascular bicollateral bundles arranged in complete cylinder (Siphonostele: eustele). Two types of bicollateral vascular bundles were present, i. e., large and small bundles. Data presented in Table 6 revealed that, effect of the remaining toxicity of PM in clay soil on some anatomical parameters of cucumber seedling hypocotyl after it treated with compost and P. putida (E1). Application of PM increased seedling hypocotyl cross section, cortex and conductive vascular tissues (xylem and phloem) thickness compared with the other treatments. On the other hand, the lowest number of vessels per bundle was reduced by PM treated soil in comparison with the other treatments. Application of compost and P. putida (E1) decreased these anatomical parameters compared to control (Table 6).
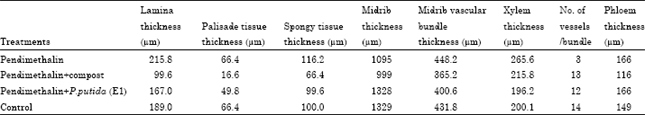
The leaf lamina internal structure of C. sativus plants was similar to other dicotyledons plants. It consists of upper and lower epidermis and mesophyll tissue, which differentiate into palisade and spongy parenchyma. Epidermis, one layer of completely arranged parenchymatous cells, which were flattened parallel to the leaf surface. The palisade parenchyma cells elongated and completely arranged. The spongy parenchymatous cells loosely arranged with numerous large intercellular spaces. Data presented in Table 7 and Fig. 3 indicated that, anatomical parameters of the first true leaf of C. sativus show similar trend as those of seedling hypocotyl. Lamina, palisade, spongy, conductive vascular tissues as well as midrib thickness were induced by PM soil treatment in comparing with other treatments. Number of vessels per bundle was 3 for PM treatment and 13, 12, 14 for compost, P. putida (E1) and control, respectively. Irregular xylem vessels and were noticed not only in hypocotyl but also in the midrib vascular bundle of the first true leaf treated with PM. Moreover, xylem vessel diameters seem to be smaller in comparing with other treatments.
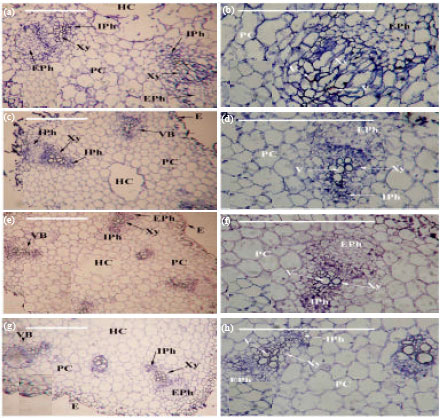 | |
| Fig. 2(a-h): | Cross sections through cucumber (C. sativus) hypocotyl, a-b: PM, c-d: PM+P. putida (E1), e-f: PM+compost, g-h: Control (untreated), HC: Hypocotyls cavity, Xy: Xylem, Eph: External phloem, Iph: Internal phloem, C = Cortex (Co), V: Vessel, Bar = 500 μm |
| Table 6: | Effect of the remaining toxicity of pendimethalin (PM) in clay soil on some anatomical parameters of cucumber seedling hypocotyl after it was treated with compost and P. putida (E1) |
 | |
| Table 7: | Effect of the remaining toxicity of pendimethalin in clay soil on some anatomical parameters of the first true leaf of cucumber plants after it was treated with compost and P. putida (E1) |
 | |
Concerning E. crus-galli leaf, internal structure of leaf lamina is similar to other moncotyledonous plants.
 | |
| Fig. 3(a-h): | Cross sections through the first true leaf of C. sativus plants, a-b: Pendimethalin, c-d: Pendimethalin+P. putida (E1), e-f: Pendimethalin+compost, g-h: Control (untreated), HC: Hypocotyls cavity, Xy: Xylem, Eph: External phloem, IPh: Internal phloem, VC: Vascular cambium, V: Vessel, UE: Upper epidermis, LE: Lower epidermis, T: Trichome, PT: Palisade tissue, ST: Spongy tissue, Bar = 500 μm |
| Table 8: | Effect of the remaining toxicity of PM in clay soil on E. crus-galli leaf lamina cross section thickness after it was treated with compost and P. putida (E1) |
 | |
Data in Fig. 4 and Table 8 indicted that, application of PM increased leaf lamina thickness and the cells of upper and lower epidermis seemed to be wider compared with untreated plants. On the other hand, application of PM in combination with each compost and P. putida (E1) caused a reduction in lamina thickness relative to the control. The lowest leaf lamina thickness was obtained by application of PM in combination with P. putida (E1). Leaf cells seem to be wider in each PM and in the combination with compost and P. putida (E1) than the control plants. It is interesting to indicate that, the internal growth parameters were concomitant with the growth parameters. No available literature was found concerning the anatomical differences, which might be useful for understanding the effect mechanisms of PM on cucumber plants.
Herbicides play an important role in the production of vegetables but their residues may cause numerous environmental problems. First of all, they may contaminate surface and groundwater through leaching and run-off.
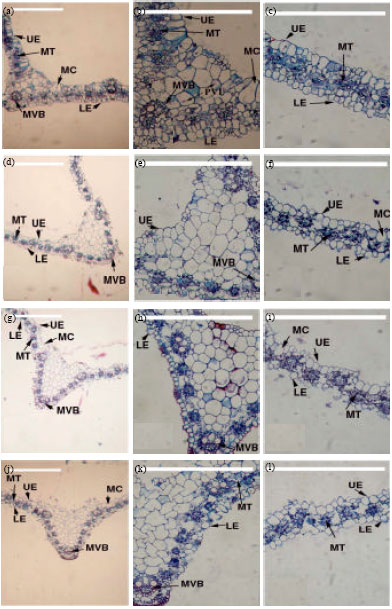 | |
| Fig. 4(a-l): | Cross sections through leaf lamina of E. crus-galli plants, a-c: PM, d-f: PM+compost, g-i: PM+P. putida (E1), j-l: Control (untreated), UE: Upper epidermis, LE: Lower epidermis, MT: Mesophyll tissue, MC: Motor cells, MVB: Midrib vascular bundle, Bar = 500 μm |
They may also remain on the soil surface and potentially affect quality and yield of the next crop cultivated on the same field. Finally, stable herbicides might be taken up by a plant forming unwanted residues. With regard to plants, PM shows differential toxicity to various species and there is a formulation-dependent toxicity to non target plant species.
PM is similar to other broad-spectrum herbicides in that it is phytotoxic to crop species to some extent. In the development of herbicides, screening experiments are conducted to ascertain extent of phytotoxicity to crops. The phytotoxicity of PM to crop species has been the focus in numerous experiments, for example, rice grain and straw yield (Devi and Gowda, 1985), root suppression of pampas grass (Green et al., 1997), effect of repeated application on cotton yield and quality, cotton fiber quality and yield (Keeling et al., 1996). Tylicki et al. (2010) observed inhibition of root elongation of Allium cepa after 48 h of incubation with PM. This effect was caused by the inhibition of mitoses varying from 1/3 to 1/2 in the case of 0.033, 0.066 and 0.099 g L-1 of pindimethalin and almost complete restriction of mitoses under higher concentrations. PM caused mitotic disturbances (c-metaphases, anaphasal and telophasal chromosome bridges, multipolar anaphases) and interphase abnormalities (micronuclei, multinuclear cells). This effect was irreversible during a 48 h postincubation in water. Mitotic disturbances were caused by abnormalities in the organization of the tubulin cytoskeleton. It suggests that even small amounts of PM can be a danger for dividing cells and embryos.
There is indication that dinitroaniline herbicides (include PM) inhibit photosynthesis, oxidative phosphorylation, protein, nucleic acid and lipid synthesis (Moreland et al., 1972). Cotyledonary leaf and hypocotyls and first true leaf of C. sativus seedling treated with PM seem to be dark green colors, swelling and brittleness. PM treatment caused reduction in primary root length and number of lateral roots. This may be due to the ridicule is the first organ to come directly contact with PM in the soil. Compost and P. putida (E1) disappeared the dark green colors and improved of the lateral roots. Smith (2006) recorded, that PM markedly inhibited the growth of both seedling weeds and crops. It was found that, dinitroaniline herbicides kill seedling weeds by inhibiting the development of lateral roots in susceptible plants, stunting the above-ground parts, with the development of a dark green color, swelling and brittleness of the stem or seedling hypocotyl (Parka and Soper, 1977). Severe crop phytotoxicity and damage symptoms reported in literature range from reduced or inhibited germination, reduced root length, protein and nucleic acid contents of root tips, injured flowers, to complete crop failure and residual persistence of herbicides in crop and soil (Henderson and Webber, 1993; Sinha et al., 1996). PM caused seedling mortality (Aluka, 1997), but doesn't prevent seedling emergence (Akobundu, 1984; Smith, 2004). In the present study, applications of PM reduced seedling emergence lower than the other treatments. The higher seedling phytotoxicity could be attributed to PM concentration in soil. Thus, a carryover of herbicide residues from one crop season to following one may occur even through the development of modern herbicides has been directed toward a short half-live in the environment (Fayez and Kristen, 1996).
Bioremediation of pendimethalin contaminated soil test: Results in Fig. 5 show the degradation rate of PM by P. putida (E1) and compost in clay soil. PM was degraded similar by P. putida (E1) and compost. PM half-lives were 4.67 and 5 days for P. putida (E1) and compost in clay soil, respectively. PM half-live was 51.9 days in untreated clay as control treatment. The loss of PM treatment was 23% and this may be due to evaporation, drift or leaching. The trend of degradation rate of PM by bacterial strain and compost was similar in the tested soil. The obtained results showed that the bacterial strain and compost play an outstanding role in degradation of PM in clay soil. Exhibited increasing in loss of PM after initial phase (7 days) and thereafter degradation of the PM was increased gradually till end of the incubation time 28 days and this may be due to accumulation of biodegradation products.
Biodegradation of pesticides in soil was reported with microorganisms and compost (Liu et al., 1995; Belal et al., 2008). Previous studies by (Karpouzas and Walker, 2000) have reported the isolation of two ethoprophos-degrading P. putida strains, which were also able to degrade cadusafos but in a less efficient way compared to the Flavobacterium and Sphingomonas strains. Flavobacterium strains have been reported to be responsible for the degradation of carbofuran (Chaudhry and Ali, 1988).
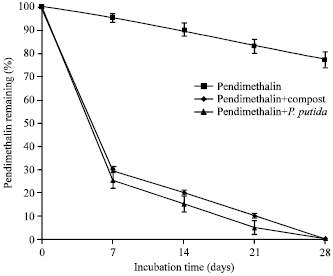 | |
| Fig. 5: | Bioremediation of PM contaminated soil by P. putida (E1) and compost and their effects on population of microorganisms in soil |
More potent strains that degraded PM rapidly were obtained from a soil samples which PM had been applied or exposed for a number of years or the time an enrichment technique. This indicates that repeated applications or exposure of soil or mature compost to xenobiotic compounds for a long period of time can result in the evolution of microorganism's capability of degrading these compounds rapidly and more extensively.
Although addition of these bioprocessed materials has been an integral part of sustainable agriculture practices and offers a good nutrient source for microbes (Laine and Jorgensen, 1996) and enhancers of microbial activity include moisture, inorganic nutrients and oxygen. There are many well-established bioremediation technologies applied commercially at contaminated sites. One of such technology is the use of compost material and biogas slurry. Compost is rich sources of microorganisms, which can degrade contaminants to innocuous compounds such as carbon dioxide and water.
Earlier studies have also reported that bioprocessed materials such as compost and biogas slurry were used to degrade of atrazine herbicide in contaminated soil using various bioprocessed materials (Liu et al., 1995). Due to their high organic matter content, all bioprocessed materials accelerated cadusafos and carbofuran breakdown. Earlier studies have also reported high microbial biomass in soil that received the organic carbon amendment (Devi and Gowda, 1985), who found that addition of compost provided a rich source of microorganisms. Kulshrestha and Singh (1992) observed that 11-14% of PM degradation could be attributed to microbial transformation in sandy soil after 91 day. Smith et al. (1979) reported that, after application to soil, PM may dissipate through evaporation, drift, leaching and runoff. A laboratory experiment simulating winter conditions showed that as much as 10% of the applied pendimethalin (0.6 mg kg-1 applied) evaporated if it was applied on the soil surface. Nayak et al. (1994) investigated the effect of PM on populations of bacteria, fungi and actinomycetes in sesame soil (sandy loam, pH 5.8, available N, P and K 21, 23.7 and 53.75 kg ha-1, respectively) at Bhubaneshwar, India. The dilution plate method was used to enumerate populations of bacteria, fungi and actinomycetes from soil samples. It was found that PM (0.5 kg ha-1) significantly reduced bacteria (61%) after 25 days but not after 50 and 75 days, at which time a slight stimulation was noted as compared with the unweeded control. Fungi were significantly reduced by 19% after 25 days and stimulated after 50 and 75 days as compared with unweeded control. Actinomycetes were substantially reduced by 21% after 25 days and stimulated after 50 and 75 days. Sidhu et al. (1985) and Barua et al. (1991) studied the effect of PM on populations of fungi, bacteria and actinomycetes. A significant decrease was observed on the first few days after the application, but after a period of 6 weeks, recovery to the level of the control was reached or almost reached. Bacteria were almost unaffected after 42 days, while actinomycetes were the most one.
CONCLUSION
In the present study, bioremediation of PM-contaminated soil was studied by addition of pure culture from P. putida (E1) and compost in 28 days. P. putida (E1) and compost showed high ability in PM degradation. There was no toxicity of PM detected in clay soil after it treated with P. putida or compost on C. sativus (non-target crop) and E. crus-galli (target plants), therefore these residues did not affected the following economical crops. It was observed that clay soil without any amendment (i.e. control) showed least degradation of PM. PM significantly decreases germination rate and increases in seedling mortality rate of the tested plants. The results suggest that bioremediation by P. putida (E1) strain and compost were considered to be the effective method for detoxification of PM in soil system.
ACKNOWLEDGMENT
This study was supported by Kafrelsheikh University (Researches Support Fund), Egypt.
REFERENCES
- Ambrus, A., J. Lantos, E. Visi, I. Csatlos and L. Sarvari, 1981. General method for determination of pesticide residues of plant origin, soil and water, I. Extraction and cleanup. J. Assoc. Off. Anal. Chem., 64: 733-742.
Direct Link - Appleby, A.P. and B.E. Valverde, 1988. Behavior of dinitroaniline herbicides in plants. Weed Technol., 3: 198-206.
Direct Link - Barua, A.S., J. Saha, S. Chaudhury, A. Chowdhury and N. Adityachaudhury, 1991. Degradation of pendimethalin by soil fungi. Pestic. Sci., 29: 419-425.
CrossRef - Chaudhry, G.R. and H.D. Ali, 1988. Bacterial metabolism of carbofuran. Applied Environ. Microbiol., 54: 1414-1419.
Direct Link - Liu, X., A.M. Cole and L. Zhang, 1995. Remediation of pesticide contaminated soil by planting and composting addition. Compost Sci. Utilizat., 3: 20-30.
Direct Link - Gan, J., R.I. Becker, W.C. Koskinen and D.D. Buhler, 1996. Degradation of atrazine in two soils as a function of concentration. J. Environ. Qual., 25: 1064-1072.
Direct Link - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedures for Agricultural Research. 2nd Edn., John Wiley and Sons Inc., Hoboken, New Jersey, ISBN: 978-0-471-87092-0, Pages: 704.
Direct Link - Green, J.C., G.J. Keever, C.H. Gilliam, C.K. Palmer, J.W. Olive and D.J. Eakes, 1997. Effects of preemergence-applied herbicides on pampas grass grown in containers. J. Environ. Hortic., 15: 77-80.
Direct Link - Grigg, B.C., N.A. Assaf and R.F. Turco, 1997. Removal of atrazine contamination in soil and liquid systems using bioaugmentation. Pestic. Sci., 50: 211-220.
CrossRef - Guo, L., T.J. Bicki, A.S. Felsot and T.D. Hinesly, 1991. Phytotoxicity of atrazine and alachlor in soil amended with sludge, manure and activated carbon. J. Environ. Sci. Health B Pest. Food Contam. Agric. Wastes, 26: 513-527.
CrossRef - Gutmann, M., 1995. Improved staining procedures for photographic documentation of phenolic deposits in semithin sections of plant tissue. J. Microsc., 179: 277-281.
CrossRef - Henderson, C.W.L. and M.J. Webber, 1993. Phytotoxicity to transplanted lettuce (Lactuca sativa) of three pre-emergence herbicides: Metolachlor, pendimethalin and propachlor. Aust. J. Exp. Agric., 33: 373-380.
CrossRef - Jazwa, A., E. Szpyrka and J. Sadlo, 2009. Disappearance of pendimethalin in soil and its residue in ripe fennel. J. Cent. Eur. Agric., 10: 153-158.
Direct Link - Karpouzas, D.G. and A. Walker, 2000. Factors influencing the ability of Pseudomonas putida strains epI and II to degrade the organophosphate ethoprophos. J. Applied Microbiol., 89: 40-48.
CrossRefPubMedDirect Link - Keeling, J.W., P.A. Dotray and J.R. Abernathy, 1996. Effects of repeated applications of trifluralin and pendimethalin on cotton (Gossypium hirsutum). Weed Technol., 10: 295-298.
Direct Link - Kulshrestha, G. and S.B. Singh, 1992. Influence of soil moisture and microbial activity on pendimethalin degradation. Bull. Environ. Contam. Toxicol., 48: 269-274.
CrossRef - Laine, M.M. and K.S. Jorgensen, 1996. Straw compost and bioremediated soil as inocula for the bioremediation of chlorophenol-contaminated soil. Applied Environ. Microbiol., 65: 1507-1513.
Direct Link - Leoni, V., C. Cremisini, R. Giovinazzo, G. Puccetti and M. Vitali, 1992. Activated sludge biodegradation test as a screening method to evaluate persistence of pesticides in soil. Sci. Total Environ., 123-124: 279-289.
CrossRef - Luke, M.A., J.E. Froberg, G.M. Doose and H.T. Masumoto, 1981. Improved multiresidue gas chromatographic determination of organophosphorus, organonitrogen and organohalogen pesticides in produce, using flame photometric and electrolytic conductivity detectors. J. Assoc. Official Anal. Chem., 64: 1187-1195.
PubMedDirect Link - Luke, M.A., J.E. Froberg and H.T. Masumoto, 1975. Extraction and cleanup of organochlorine, organophosphate, organonitrogen and hydrocarbon pesticides in produce for determination by gas-liquid chromatography. J. Assoc. Off. Anal. Chem., 58: 1020-1026.
PubMedDirect Link - Megadi, V.B., P.N. Tallur, R.S. Hoskeri, S.I. Mulla and H.Z. Ninnekar, 2010. Biodegradation of pendimethalin by Bacillus circulans. Indian J. Biotechnol., 9: 173-177.
Direct Link - Moran, R. and D. Porath, 1980. Chlorophyll determination in intact tissues using N, N-demethylformamide. Plant Physiol., 65: 478-479.
CrossRefPubMedDirect Link - Moreland, D.E., F.S. Farmer and G.G. Hussey, 1972. Inhibition of photosynthesis and respiration by substituted 2,6-dinitroaniline herbicides: II. Effects on responses in excised plant tissues and treated seedlings. Pestic. Biochem. Physiol., 2: 354-363.
CrossRef - Nayak, B.S., J.C. Prusty and S.K. Mohanty, 1994. Effect of herbicides on bacteria, fungi and actinomycetes in sesame (Sesamum indicum) soil. Indian. J. Agric. Sci., 64: 888-890.
Direct Link - Parka, S.J. and O.F. Soper, 1977. The physiology and mode of action of the dinitroaniline herbicides. Weed Sci., 25: 79-87.
Direct Link - Pattanasupong, A., H. Nagase, M. Inoue, K. Hirata, K. Tani, M. Nasu and K. Miyamoto, 2004. Ability of microbial consortium to remove pesticide, carbendazim and 2,4- dichlorophenoxyacetic acid. World J. Microbiol. Biotechnol., 20: 517-522.
CrossRef - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link - Sidhu, P.S., S.C. Bhandari, S.P. Mehra and H.S. Gill, 1985. Effect of fluchloralin, oxadiazon and pendimethalin on soil microflora and nodulation in groundnut. J. Res. Punjab Agric. Univ., 22: 33-38.
Direct Link - Smith, A.E., A.J. Aubin and T.C. McIntosh, 1995. Field persistence studies with emulsifiable concentrate and granular formulations of the herbicide pendimethalin in Saskatchewan. J. Agric. Food Chem., 43: 2988-2991.
CrossRef - Smith, M.A.K., 2006. Comparing weed and crop seedling response to pre-emergence pendimethalin application in Corchorus olitorius and Abelmoschus esculentus. Crop Prot., 25: 1221-1226.
CrossRef - Smith, M.A.K., 2004. Pendimethalin phytotoxicity and seedling weed control in Indian spinach (Basella alba L.). Crop Prot., 23: 201-204.
CrossRef - Strandberg, M. and J.J. Scott-Fordsmand, 2004. Effects of pendimethalin at lower trophic trophic levels-a review. Ecotoxicol. Environ. Saf., 57: 190-201.
CrossRef - Ahtiainen, J., R. Valo, M. Jarvinen and A. Joutti, 2000. Microbial toxicity tests and chemical analysis as monitoring parameters at composting of creosote-contaminated soil. Ecotoxicol. Environ. Saf., 53: 323-329.
CrossRef - Tylicki, A., E. Kowalska-Wochna, A. Oscilowicz and R. Ilasz, 2010. Mitodepressive and cytotoxic effects of short-term exposure to relatively small doses of pendimethalin evaluated by allium test, polish. J. Environ. Stud., 19: 193-199.
Direct Link - Zheng, S.Q. and J.F. Cooper, 1996. Adsorption, desorption and degradation of three pesticides in different soils. Arch. Eviron. Contam. Toxicol., 30: 15-20.
CrossRef