
M. A. Ali
Department of Agronomy, Faculty of Agriculture, South Valley University, Qena, Egypt
Asian Journal of Crop Science
Year: 2012 | Volume: 4 | Issue: 1 | Page No.: 23-31







ABSTRACT
The objective of the study was to compare the effectiveness of pedigree and independent culling levels methods for early selection in the F3 and late selection in the F5 generations. The experiments were conducted at South Valley University Experimental Farm at Qena during 2007 through 2010 summer seasons. The F3, F4, F5 and F6 generations of the grain sorghum hybrid (Giza 15 x Dorado) were studied. Highly significant differences were observed among F3 families for all studied traits. After three cycles of selection, the genetic variability retained after independent culling level was larger and more pronounced than that after single trait selection. However, the genetic variability retained in the F6 was larger after late than after early selection. Late pedigree selection in the F5 increased grain yield/plant by 9.57% compared to 12.70% from the bulk sample after three cycles of selection for this trait started early in the F3. Selection for early flowering and 1000-grain weight improved these traits but decreased yield in both early and late selection. Late selection in the F5 generation in these materials was more efficient than early selection in the F3 families. Pedigree selection though proved to be an efficient selection procedure for improving the selected traits. However, it was accompanied with some deleterious effects in some unselected related traits. ICL caused moderate increases in most measured traits that were comparable for early and late selection.
PDF Abstract XML References Citation
Received: September 05, 2011;
Accepted: November 14, 2011;
Published: December 31, 2011
How to cite this article
M. A. Ali, 2012. Effectiveness of Selection in the F3 and F5 Generations in Grain Sorghum. Asian Journal of Crop Science, 4: 23-31.
DOI: 10.3923/ajcs.2012.23.31
URL: https://scialert.net/abstract/?doi=ajcs.2012.23.31
DOI: 10.3923/ajcs.2012.23.31
URL: https://scialert.net/abstract/?doi=ajcs.2012.23.31
INTRODUCTION
Grain sorghum (Sorghum bicolor (L.) Moench) is one of the most important cereal crops in Egypt in the summer season. It ranks fourth after wheat, rice and maize (Smith and Frederikson, 2000). Grain sorghum production is concentrated in Upper Egypt, particularly in Assiut and Sohag provinces. Haussmann et al. (2000) stated that grain sorghum can be grown under a wide range of environments i.e., low soil fertility, poor land establishment, hot weather, drought stress and salinity. Therefore, efforts are currently devoted to increase the cultivated area of grain sorghum in Upper Egypt in newly reclaimed desert land.
High-yielding, tall sorghum cultivars were crossed to shorter sorghum varieties to develop improved high-yielding cultivars with a shorter stature (Rosenow and Dahlberg, 2000). An important aspect of breeding in sorghum is the development of dual purpose varieties. Therefore, it is desirable to have plants which have high grain yield. This attribute is dependent upon several components; hence, assessment of the contribution of different morphological characters to productivity is of vital objective in sorghum breeding. However, grain yield is a complex character which is influenced by the fluctuation of environmental conditions (Biswas et al., 2001).
Plant breeders are continuously searching for more reliable and efficient selection procedures. The practical value of a plant is affected by several traits. Selection for yield is one of the most difficult challenges of plant breeding. However, selection based upon yield per se should be the most efficient method of increasing the mean yield of a population (Wells and Kofoid, 1986). Yield components and plant traits contributing to grain yield are important for breeding strategies (Singh and Chaudhary, 2006). The pedigree selection method can be used to identify superior genotypes for grain yield in a cultivar development program.
There are opposing views on the genetic theory that underlies selection for quantitative characters, such as grain yield. First is that the phenotype of the heterozygote is not a reliable guide to the lines which might be derived from it. Second is as the proportion of homozygote in early generations is very small, selection should be delayed until later generations (Allard, 1960). Selection for yield potential using yield of single plants in early generations (F3 or F4) within crosses, however, has been ineffective (Golabadi et al., 2006). The objectives of the present study were: (1) to compare the effectiveness of early selection in the F3 and late selection in the F5 generations using the pedigree and independent culling levels methods and (2) to investigate the relationship between the F3 to F5 generations in a segregating population of grain sorghum originated from the cross between Giza 15 and Dorado grain sorghum varieties.
MATERIALS AND METHODS
The population (Giza 15 x Dorado) used in this study was developed by author at South Valley University Experimental Farm at Qena in 2006 summer season. The materials included 100 F3 families traced back to a random sample F2 single plants originated from the cross (Giza 15 x Dorado). The field experiments were conducted at South Valley University Experimental Farm at Qena during 2007 through 2010 summer seasons. The soil type was sandy loam. Some physical and chemical properties of a representative soil sample of the experimental site have been presented in Table 1.
Early selection: The seeds of each F3 plant were divided into two parts: the first part of the seeds was sown during the 2007 summer season in a randomized complete block design of three replications. Each family was grown in a single row plot in each replicate. Rows were 4 m long and 60 cm wide with hills spaced 20 cm apart within a row. Hills were over seeded and after full emergence, hills were thinned to two plants per hill. All cultural practices were applied as recommended for grain sorghum production in Upper Egypt. Days to 50% flowering were recorded on a plot mean basis, as days from planting to the day of 50% flowering. The earliest head of each plot was labeled and bagged. At maturity, individual plant data were recorded on ten random plants from the middle part of each plot. Other recorded traits included plant height (cm), panicle length (cm), panicle width (cm), 1000-grain weight (g) and grain yield/plant (g).
| Table 1: | Some physical and chemical properties of a representative soil sample of the experimental site† |
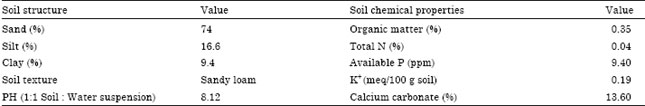 | |
| †: Data were kindly supplied by the Department of Soils, Faculty of Agriculture, South Valley University, Qena, Egypt | |
Analysis of variance for the randomized complete blocks design was carried out according to Snedecor and Cochran (1980). Single trait selection was applied between and within families after the analysis of variance of each selection criterion. Seeds of the best plant from each of the best 20 families for each selection criterion viz., flowering date, 1000-grain weight and grain yield/plant were saved to grow the F4-families. In addition, the best plant from the best 20 families was selected based on independent culling levels for six traits (flowering date, plant height, panicle length (cm), panicle width (cm), 1000-grain weight and grain yield/plant).
In 2008 summer season, the parents, the F4-families selected for each selection criterion and the F4-random bulk sample were sown in a randomized complete blocks design of three replications. The data were recorded as in the previous season. Each group of 20 families for each selection criterion and by independent culling levels was analyzed separately. The best plant from the best 10-families for each selection criterion and independent culling levels were saved. The same procedure was repeated until the F6-generation.
Late selection: In 2007 summer season, the second part of the saved seeds was planted to give the F4 families for late (F5) selection. Each plant was sown in a single row in a nursery in a non replicated trial along with the parents and a random bulk sample. Rows were (100 rows) 4 m long, 60 cm apart and 20 cm between hills within a row.
In 2008 summer season, a bulk seed from each family was used to give the next generation (F4). In the F5-generation, three replications were sown from the bulk seed of each family, along with the parents and the F5-random sample. Plot size and row and hill spacing were similar to the early selection experiment. Seeds of the best plant from the 10-best families for each selection criterion were saved for evaluation in the F6-generation.
Statistical analysis: Analysis of variance of trait means and covariance between pairs of traits were calculated according to Snedecor and Cochran (1980). Differences between means were tested by the revised LSD according to El-Rawi and Khalafalla (1980). Genotypic and phenotypic coefficients of variation were estimated on plot mean basis according to Burton (1952). Broad sense heritability was estimated following Walker (1960).
The observed gain from selection as percent deviation of the mean of selected families from the better parent and from the bulk sample means, for each selection criterion and the correlated traits were computed.
RESULTS AND DISCUSSION
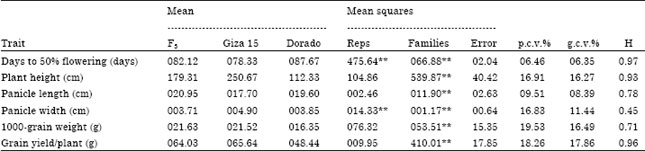
Base population: Analysis of variance revealed significant differences (p<0.01) among families of the base population for all traits under study viz., days to 50% flowering; plant height, panicle length, panicle width, 1000-grain weight and grain yield/plant (Table 2). This indicated the presence of genetic variability among the selected families for these traits.
The genotypic coefficient of variability (Table 2) was high for plant height (16.27%), panicle width (11.44%), 1000-grain weight (16.49%) and grain yield/plant (17.86%). Moderate variability was also observed for days to 50% flowering (6.35%) and panicle length (8.39%). This may be due to the genetic diversity between the crossed parents. Giza 15 headed after 78.33 days, while Dorado headed after 87.67 days from sowing. Plant height also varied from 112.33 cm for Dorado to 250.67 cm for Giza 15. The 1000-grain weight ranged from 16.35 g for Dorado to 21.52 g for Giza 15.
| Table 2: | Mean performance, mean squares, genotypic and phenotypic coefficients of variability and heritability in broad sense of six traits in the base population (F3 generations) |
 | |
| *, **Significant and highly significant at 0.05 and 0.01 level of probability, respectively | |
Grain yield/plant varied from 48.44 g for Dorado to 65.64 g for Giza 15. Differences between the parents appeared to be responsible for most of the differences among the F3 families. The magnitudes of phenotypic and genotypic coefficients of variability, which are characteristics of the original population, have a considerable effect on early generation selection. Response to selection is expected to be greater when selection is practiced in populations having both larger phenotypic and genotypic variances.
Large estimates of broad sense heritability were obtained for all traits under study except panicle width (Table 2). However, estimates of heritability seemed to be biased upwards due to the confounding effects of years and locations of the estimates of genetic variance obtained from one location for one season (O’Brien et al., 1978). Mahdy et al. (1999a, b) reported estimates of broad sense heritability for some field traits in F3 families of grain sorghum. It ranged from 72.9 to 95.7% and 99.4 to 99.5% for days to 50% flowering and plant height, respectively. However, it ranged from 83.4 to 94.7%, 60.7 to 94.0%, 96.3 to 97.4% and 96.1 to 97.4% for panicle length, panicle width, 1000-grain and grain yield/plant, respectively. Our results are in line with those observed by Biswas et al. (2001) and Ali (2002, 2011a, b).
Early selection in the F3-generation
Single trait selection: Means and genotypic coefficients of variability of cycles 1, 2 and 3 (F4, F5 and F6) of selection for each of flowering date, 1000-grain weight and grain yield/plant and correlated traits are presented in Table 3. Pedigree selection for days to 50% flowering decreased the genotypic coefficient of variability (g.c.v.) from 4.61% after the first cycle to 2.09% after the third cycle. However, selection for 1000-grain weight and grain yield/plant greatly decreased the genetic variability of these traits from 9.31% and 13.16% after the first cycle to 4.14% and 3.89% after the third cycle, respectively. This indicates that early selection in the F3 generation rapidly decreased the g.c.v. of the selection criterion after three cycles. Mahdy et al. (1999a, b) and Ali (2002, 2011a, b) and reached the same conclusion after two cycles of pedigree selection. In contrast, Ali et al. (2006) reported increased g.c.v. after two cycles of pedigree selection for grain yield/plant and its correlated traits viz., flowering date, plant height and 1000-grain weight.
The observed gains and correlated responses to selection measured as the deviation of the overall cycle mean from the better parent and the bulk sample are presented in Table 4. After the third cycle, pedigree selection for days to 50% flowering improved earliness by -14.75% over the bulk sample. However, selection for earliness was accompanied by deleterious effects on the other correlated traits reaching, -0.44, -2.27, -5.09, -5.68 and -4.46% for plant height, panicle length, panicle width, 1000-grain weight and grain yield/plant, respectively. Selection for 1000-grain weight resulted in an observed gain of 14.19% in this trait over the bulk sample.
| Table 3: | Means and genotypic coefficients of variability estimates for traits of grain sorghum during three cycles of pedigree selection and selection by independent culling levels (ICL) |
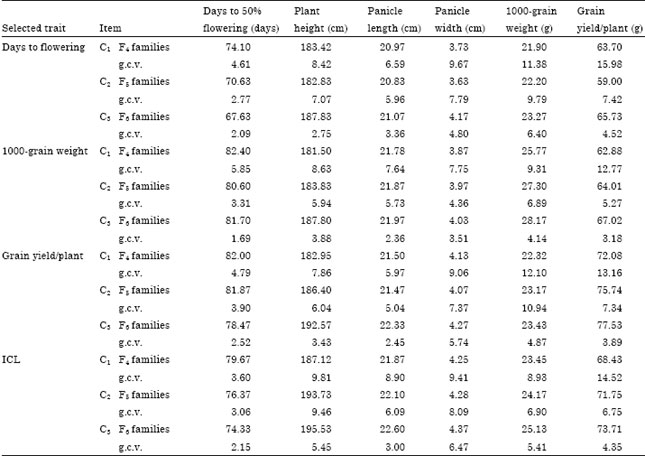 | |
| g.c.v.: Genotypic coefficients of variability | |
However, selection for 1000-grain weight was accompanied with favourable decrease in plant height (-0.46%) and a 1.90% increase in panicle length from the bulk sample. While, it was accompanied with increase in flowering date (2.98%) and but adversely affected panicle width (-8.12%) and grain yield/plant (-2.58%) from the bulk sample. Selection for grain yield/plant resulted in a 12.70% gain over the bulk sample. Three cycles of pedigree selection for grain yield/plant also increased plant height and panicle length and caused a favourable decrease in days to 50% flowering and negatively affected panicle width and 1000-grain weight. Mahdy et al. (1999a, b), reported that grain yield/plant was increased by 25.74 and days to flowering were decreased by -4.83 from the bulk sample, respectively, after two cycles of pedigree selection in a segregating population. Similar results were reported by Ismail et al. (1996), Ali (2002), Benmoussa and Achouch (2005) and El-Morshidy et al. (2010).
Selection by independent culling levels: Means and genotypic coefficients of variability of cycles 1, 2 and 3 of selection by the independent culling levels for the studied traits are illustrated in Table 3. After three cycles of selection by independent culling levels, the g.c.v. of days to flowering decreased to 2.15% compared with 2.09% for single trait selection and 5.41% for 1000-grain weight compared with 4.14% and to 4.35% for grain yield/plant compared with 3.89% in single trait selection for these traits.
| Table 4: | Realized gains and correlated responses from pedigree selection and independent culling levels (ICL) measured in percentage from the bulk sample and the better parent |
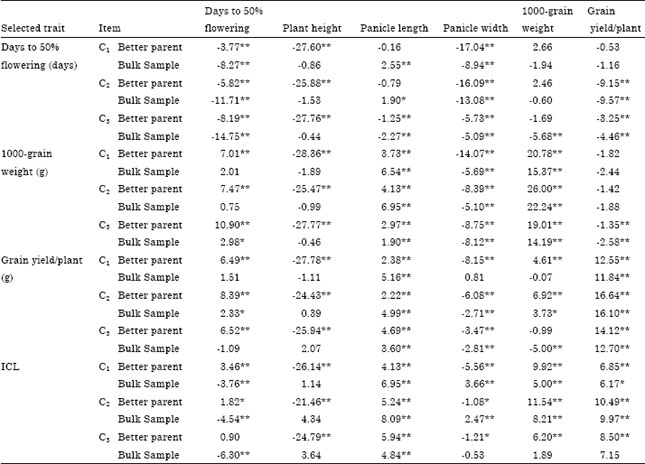 | |
| ***,**Significant at 0.05 and 0.01 levels of probability, respectively. Negative value meant that the overall cycle mean were earlier, shorter, smaller, lower, lighter and lower than the better parent and the bulk sample mean for flowering date, plant height, panicle length, panicle width, 1000-grain weight and grain yield/plant, respectively | |
Therefore, it may be concluded that genetic variability retained after selection by independent culling levels was more pronounced and larger than that after single trait selection. These results were expected because single trait selection was directed to one trait (one selection criterion), whereas selection by independent culling levels was directed to all studied traits. Ismail et al. (1996) and Mahdy et al. (1999a, b) found that the g.c.v. of each of flowering date, 1000-grain weight and grain yield/plant was larger after independent culling levels than after single trait selection.
The results presented in Table 4 show that the observed gains from three cycles of selection by ICL reached -6.30, 1.89 and 7.15% relative to the bulk sample for flowering date, 1000-grain weight and grain yield/plant, respectively, compared with -14.75, 14.19 and 12.70% for single trait selection. It may be concluded, therefore, that single trait selection is an efficient procedure for improving each of the studied traits, but was accompanied with deleterious decreases in some other studied traits. In contrast, ICL showed intermediate genetic gain for all studied traits. Similar results were reported by Ismail et al. (1996), Mahdy et al. (1999a, b) and El-Morshidy et al. (2010).
| Table 5: | Means and genotypic coefficient of variation estimates for the studied traits of F6 families after pedigree selection and selection by independent culling levels (ICL) |
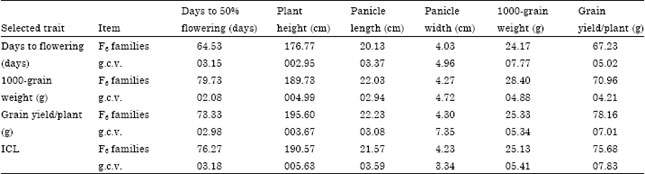 | |
| g.c.v.: Genotypic coefficient of variation | |
| Table 6: | Observed gains from and correlated responses to pedigree selection and selection by independent culling levels (ICL) measured in percentage from the mean of bulk sample and the better parent mean |
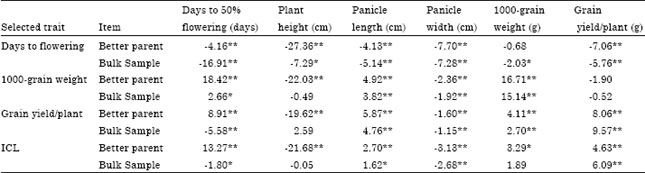 | |
| *,**Significant at 0.05 and 0.01 levels of probability, respectively. Negative value meant that the overall cycle mean were earlier, shorter, smaller, lower, lighter and lower than the better parent and the bulk sample mean for flowering date, plant height, panicle length, panicle width, 1000-grain weight and grain yield/plant, respectively | |
Late selection in the F5-generation
Single trait selection: Single trait selection was applied on the 100 F5-families resulting from bulk propagation of F3-families. Data in Table 5 indicate that the g.c.v. was high for the selection criteria in the late compared to early selection and accounted by 3.15, 4.88 and 7.01% for flowering date, 1000-grain weight and grain yield/plant, respectively, compared to 2.09, 4.14 and 3.89% for the same traits in the same generation (F6) and after three cycles of early selection. These results show that early selection in the F3 generation rapidly minimized and decreased the genetic variability than late selection applied in the F5-generation. Fredrickson and Kronstad (1985) suggested that selection for quantitative traits of low heritability must be delayed until later generations.
The observed gain from one cycle of single trait selection (Table 6) was -16.91, 15.14 and 9.57% for days to 50% flowering, 1000-grain weight and grain yield/plant, respectively.
While it was -14.75, 14.19 and 12.70% for the bulk sample for the respective traits of the bulk sample in the early selection.
This, it may be concluded that selection in the F5 in materials used in this study was more efficient than early selection. However, the genetic variability in each generation was greater in late than in early selection. Kontt and Kumar (1975), Sneep (1977) and Ismail et al. (1996) also concluded that early selection would be of doubtful value.
Selection by independent culling levels: After one cycle of selection by ICL the genotypic coefficient of variation indicated greater genetic variation than late and/or single trait selection for all traits under study.
The observed gain achieved from one cycle of ICL was -1.80, -0.05, 1.62, -2.68, 1.89 and 6.09% from the bulk sample for flowering date, plant height, panicle length, panicle width, 1000-grain weight and grain yield/plant, respectively. Similar results were reported by Ismail et al. (1996) and Mahdy et al. (1999a, b) and El-Morshidy et al. (2010).
CONCLUSIONS
It may be concluded that the late selection in the F5 generation in the materials used in this study was more efficient than early selection in the F3 families. However, pedigree selection, though proved to be an efficient selection procedure for improving the selected traits, it was accompanied with some deleterious effects in some unselected related traits. In addition, ICL caused moderate increases in most measured traits that were comparable for early and late selection.
REFERENCES
- Ali, M.A., 2011. Response to pedigree selection for earliness and grain yield in spring wheat under heat stress. Asian J. Crop Sci., 3: 118-129.
CrossRefDirect Link - Ali, M.A., 2011. Pedigree selection for grain yield in spring wheat (Triticum aestivum L.) under drought stress conditions. Asian J. Crop Sci., 3: 158-168.
CrossRefDirect Link - Allard, R.W., 1960. Principles of Plant Breeding. 1st Edn., John Wiley and Sons Inc., New York pp: 372-372.
CrossRefDirect Link - Benmoussa, M. and A. Achouch, 2005. Effect of water stress on yield and its composantsof some cereals in Algeria. J. Cent. Eur. Agric., 6: 427-434.
Direct Link - Biswas, B.K., M. Hasanuzzaman, F. El-Taj, M.S. Alam and M.R. Amin, 2001. Simultaneous selection for fodder and grain yield in sorghum. J. Biol. Sci., 1: 321-323.
CrossRefDirect Link - Golabadi, M., A. Arzani and S.A.M.M. Maibody, 2006. Assessment of drought tolerance in segregating populations in durum wheat. Afr. Agric. J. Res., 1: 162-171.
Direct Link - Haussmann, B.I.G., A.B. Obilana, P.O. Ayiecho, A. Blum, W. Schipprack and H.H. Geiger, 2000. Yield and yield stability of four population types of grain sorghum in a semi-arid area of Kenya. Crop Sci., 40: 319-329.
Direct Link - Kontt, D.R. and J. Kumar, 1975. Comparison of early generation yield testing and single seed descent procedure in wheat breeding. Crop Sci., 15: 295-299.
Direct Link - Singh, G.P. and H.B. Chaudhary, 2006. Selection parameters and yield enhancement of wheat (Triticum aestivum L.) under different moisture stress conditions. Asian J. Plant Sci., 5: 894-898.
CrossRefDirect Link - Snedecor, G.W. and W.G. Cochran, 1980. Statistical Methods. 7th Edn., Iowa State University Press, Iowa, USA., ISBN-10: 0813815606, Pages: 507.
Direct Link - Sneep, J., 1977. Selection for yield in early generations of self-fertilizing crops. Euphytica, 26: 27-30.
CrossRef - Walker, J.T., 1960. The use of a selection index technique in the analysis of progeny row data. Empire Cotton Grow. Rev., 37: 81-107.
Direct Link