
Salem S. Alghamdi
Department of Plant Production, College of Food and Agricultural Sciences, King Saud University, P.O. Box 2460, Riyadh 11451, Saudi Arabia
Asian Journal of Crop Science
Year: 2009 | Volume: 1 | Issue: 2 | Page No.: 66-76







ABSTRACT
The present investigation was carried out under insect-free cages during the three successive seasons 2004-2006. A diallel cross excluding reciprocals among eight faba bean genotypes was used to estimate the heterotic effects of F1 crosses and F2 populations relative to their respective mid and better parents and combining ability analysis for yield and some of its variables. Highly significant differences among the tested entries were detected for different traits, indicating wide genetic variability for all traits. Heterosis percentages relative to mid-parents were significantly positive in several crosses with a range of 15.6-19.7, 38.0-59.8, 85.0-131.4, 74.3-79.4, 54.4-127.2 and 69.8-142.0% for plant height, number of branches per plant, number of pods per plant, number of pods per main stem, number of seeds per plant and seed yield per plant, respectively. However, heterosis percentages relative to better parent were significantly positive in some crosses and recorded a range of 19.0-19.0, 54.2-73.7, 62.9-97.7 and 47.6-129.9% for plant height, number of pods per plant, number of seeds per plant and seed yield per plant, in the same order. Some crosses exhibited significantly negative heterosis compared to respective mid-parent for days to flowering (from -9.0-23.5%). The ratios of
PDF Abstract XML References Citation
How to cite this article
Salem S. Alghamdi, 2009. Heterosis and Combining Ability in a Diallel Cross of Eight Faba Bean (Vicia faba L.) Genotypes. Asian Journal of Crop Science, 1: 66-76.
DOI: 10.3923/ajcs.2009.66.76
URL: https://scialert.net/abstract/?doi=ajcs.2009.66.76
DOI: 10.3923/ajcs.2009.66.76
URL: https://scialert.net/abstract/?doi=ajcs.2009.66.76
INTRODUCTION
Faba bean (Vicia faba L.) is one among the most important nutritive seed legumes and widely considered as a good resource of protein, starch, cellulose and minerals (Haciseferogullari et al., 2003) for human in developing countries and for animal feed, mainly for horses, chickens and pigeons, in industrialized countries. In the Middle East and most part of Mediterranean, China and Ethiopia, faba bean constitutes the main dish on the breakfast and dinner tables, particularly for low income groups. Faba, a diploid species of 2n = 2x = 12 chromosomes, is botanically classified on the basis of seed shape and weight into three common categories, small rounded seeds (1 cm long), var. equina with medium sized seeds (1.5 cm) and var. major with large broad flat seeds (2.5 cm) (Tamas et al., 1998). Primarily, faba bean is a self-pollinating plant with significant levels of out-cross and inter-cross ranging from 20-80% (Suso and Moreno, 1999) depending on genotype and environmental effects. Plants that have greater access to pollinators usually produce more pods per plant, more seeds per pods; longer pods and heavier seeds than encaged plants (Aouar-Sadli et al., 2008).
The crop is becoming important in Saudi Arabia where it is consumed as fresh bean pods, fresh seeds or dry seeds. The normal growing season for faba beans is winter and it is harvested earlier before the onset of high temperature. Despite the significance of faba bean in the Saudi Arabia, annual supply from farm doesn’t cover the increasing demand of the growing population, a reason that justifies annual import of large quantities of seeds. Faba bean is one of the most efficient fixers of the atmospheric nitrogen and hence, can contribute to sustain or enhance total soil nitrogen fertility through biological N2-fixation (Lindmann and Glover, 2003). It can thus successfully be incorporated into plant rotation in Saudi Arabia intensive wheat production areas. Low production of faba bean in the Kingdom can be related to several constraints, including the lack of research emphasis on this vital crop, lack of high yielding varieties and abiotic stresses inflicted by harsh environmental conditions (hot winds, high temperature, drought and salinity problems).
Therefore, the way to the expansion faba bean production as profitable crop in Saudi Arabia lies in giving more emphasis upon research focused on identification of high-yielding genotypes of good adaptation to the local production conditions. In this regards, College of Food and Agricultural Sciences, King Saud University, Riyadh, Saudi Arabia initiated breeding program on faba bean in the early nineties to solve some of the persisting problems in faba bean production in the Kingdom of Saudi Arabia.
Literarily, genetic improvement of crop desired traits depends on the nature and magnitude of genetic variability and interactions involved in the inheritance of these traits which can be estimated using diallel cross technique. This technique of crosses has been widely employed to estimate genetic variances among parents of random individuals or inbred lines from a random-mating populations through making crosses among different individuals to produce a new genetic combination of which its performance negatively or positively may exceed that of the parents a phenomenon known as heterosis.
Exploitation of heterosis through synthetics and ultimately hybrids could pay off improving yield potential and its components in faba beans. Superiority of hybrids over the mid and better parents for seed yield was found to be associated with manifestations of heterotic effects in main yield components i.e., number of branches, number of pods, number of seeds per plant and seed index. It is clearly stated from the literature that heterotic effects ranges from significantly positive to significantly negative for different traits and were very pronounced in F1 of faba beans especially crosses among widely divergent materials and less occurred in hybrids between local varieties (Darwish et al., 2005; El-Hady et al., 2006, 2007; Duc, 1997; Schill et al., 1998; Bond and Crofton, 1999; Filippetti et al., 1999; Gasim and Link, 2007; Link et al., 2008).
In addition, an inference can be made from diallel crosses about combining ability of the parents, a general concept considered collectively for classification of an inbred line relative to its crosses performance. Such information is helpful for breeders to identify the best combiners which may be hybridized either to exploit heterosis or to build up favourable fixable genes. Several researchers have stated the significance of both general and specific combing ability effects for yield and other important traits of faba beans (Abdalla et al., 2001; Attia et al., 2002; Attia and Salem, 2006; Ghaouti and Link, 2008). Kunkaew et al. (2006) established that seed yield of azuki bean and some of its yield component characters were poly-genetically controlled by additive gene effects. Although study of heterosis of various faba cultivars has been done, further and detailed information is lacking about the segregant generation heterosis, therefore, the present investigation aimed to understand the nature of gene action and relative magnitude of heterosis and combining ability of eight faba bean diverse genotypes including F1 and F2 generations using diallel cross mating design.
MATERIALS AND METHODS
Eight diverse faba bean genotypes (Table 1) were crossed in an 8x8 diallel mating design excluding reciprocals under insect free cage at Dirab Agricultural and Experimental Research Station, College of Food and Agricultural Sciences, King Saud University, Riyadh, Saudi Arabia during 2004/05 growing season. Due to insufficient hybrids seeds of some crosses, re-hybridization was made during 2005/06 season and F2 seeds were propagated from the resulted F1 plants. The parents along with their derived F1’s (28) and F2 populations (28) were planted in 2006/07 growing season under insect free cage in a randomized complete block design (RCBD) with three replicates. Seeds were planted on ridges, 2.5 m long, 60 cm between, with single seeded hills, 20 cm apart, representing each parent as well as their F1’s and F2’s by one, one and three ridges, respectively. Fertilizers were applied at monthly intervals starting at two weeks from seedlings emergence at the following rates: 40 kg N ha-1 as urea, 40 kg P2O5 ha-1 as triple super phosphate and 20 kg K2O ha-1 as potassium sulphate.
Data and harvest were carried out on guarded twelve plants in each experimental plot (parents, F1’s and F2’s). The following traits were recorded: days to flowering, plant height (cm), number of branches per plant, number of pods per plant, number of pods per main stem, number of seeds per plant, seed yield per plant (g) and 100-seed weight (g). Data were subjected to regular analysis of RCBD on plot mean basis to test genotype variances following statistical model, considering cultivar as fixed effects:
| Yij = μ + gi + bj + eijk | (1) |
Where:
| yij | = | Observation of ith treatment in the jth block (i = 1, 2, ..., g = 6; j = 1, 2, ..., b = 10) |
| μ | = | General mean |
| gi | = | Effect of the ith cultivar |
| bi | = | Effect of the jth cultivar |
| eijk | = | Experimental error |
The heterotic effects of F1 crosses and F2 populations were estimated as percentage over mid and better parents using the following formula:
| (2) |
| (3) |
| Table 1: | Origin and some features of eight faba bean genotypes |
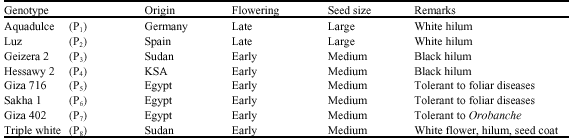 | |
Where:
| (4) |
The LSD for heterosis was computed following the formulae by Bhatt (1971).
Combining ability effects and variances were calculated according to Griffing (1956), method 2, model 1 (all possible combinations excluding reciprocals) as follow:
| Mij = μ + GCAi + GCAj + SCAij + eijk | (5) |
Where:
| Mij | = | Observation of ith cultivar in the jth block |
| μ | = | General mean |
| GCAi | = | Effect of the ith cultivar |
| GCAj | = | Effect of the jth cultivar |
| SCAij | = | Combined effect of two cultivars |
| eijk | = | Experimental error |
The estimates of variance components of GCA and SCA were calculated as follows:
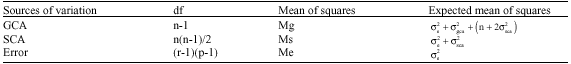 |
Where:
| Mg | = | Mean squares of GCA |
| Ms | = | Mean squares of SCA |
| Me | = | Mean squares of error |
| p | = | No. of populations |
| r | = | No. of replicates |
| n | = | No. of parental lines |
The standard error (SE) of the estimated general and specific combining ability effects were calculated as follows:
| (6) |
| (7) |
RESULTS AND DISCUSSION
Significance of Variances and Mean Performance
Mean squares of genotypes in F1 and F2 generations revealed highly significant differences among tested genotypes for all characters under investigation which indicate a wide genetic variability for studied characters and hence, the feasibility for genetic improvements using such genetic pools of faba beans.
| Table 2: | Observed mean squares of variances for different studied traits in F1 and F2 generations |
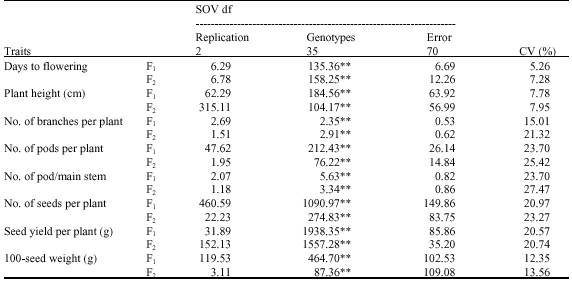 | |
| **Significant at p<0.01 level | |
On the other hand, as expected results of both populations F1 and F2 showed lower coefficient of variability (CV %) revealing that these traits have not been greatly affected by environmental factors (Table 2). The change in the value of CV from low in the F1’s generation to moderate level in the F2 generation for number of branches per plant and number of pods per main stem implied the possibility of gene segregation in a manner that an advance generation of faba bean could be under large effect of environmental factors, therefore, attention should be taken when considering such trait for selection at later generations.
As far as mean performance is concerned. The parent Geizera 2 followed by Triple white possessed the earliest plants and recorded 36.6 and 43.9 days, respectively (Table 3). Meanwhile, the two parental genotypes: Luz and Aquadulce exhibited the latest plants with 68.5 and 65.1 days, in the same order. The two parental genotypes Giza 402 and Hessawy 2 had tallest plants and recorded 108.8 and 105.5 cm, respectively, whereas the shortest plants belong to Luz (90.9 cm), followed by Aquadolce (89.9 cm). Regarding number of branches per plant, the parent Luz recorded the highest number of branches per plant (6.2) and Triple white had the lowest one (2.7). For number of pods per plant, the two parents, Triple white and Giza 402 recorded the highest number of pods (30.1 and 21.9, in the same order). Moreover, the parental genotypes Triple white had the highest values of pod number main stem (6.7). With respect to number of seeds per plant, results showed that the parental genotypes Triple white followed by Geizera 2 had the highest number of seeds per plant (62.9 and 54.1, respectively).
For seed yield per plant and 100-seed weight the two parents: Luz and Aquadulce possessed the highest values of both seed yield per plant (42.2 and 38.38) and 100-seed weight (149.1 and 130.5), respectively. It could be noticed that the parent Triple white is superior in number of pods per plant, number of pods per main stem and number of seeds per plant and the two parents Aquadulce and Luz are superior for seed yield per plant and 100-seed weight. Comparing the performance of crosses to corresponding highest parents, one (P3xP8), four (P3xP5, P3xP6, P3xP7 and P3xP8) and seven crosses (P1xP6, P2xP6, P2xP7, P3xP5, P3xP6, P4xP5 and P5xP6) significantly exceeded the highest parental genotypes for number of pods per plant, number of seeds per plant and seed yield per plant, in the same order. Whereas, one F2 cross (P2xP3) had the earliest plants (34.2 days). However, none of the crosses exceeded their highest parents for 100-seed weight. It could be concluded that the above mentioned parents and crosses would prospect in faba bean breeding and therefore may be valuable for improving seed yield via its component characters.
| Table 3: | Mean performance of eight faba bean parental genotypes and their F1 and F2 generations for yield and some of its components |
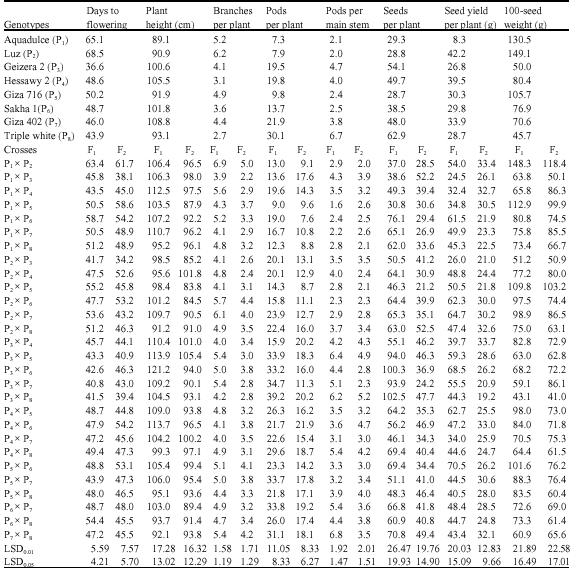 | |
Heterotic Effects
Values of heterosis percentages relative to mid (MP) parents were significantly positive in four, five, eight, two, twelve and eight crosses with a range of 15.6-19.7; 38.0-59.8; 85.0-131.4; 74.3-79.4; 54.4-127.2 and 69.8-142.0% for plant height, number of branches per plant, number of pods per plant, number of pods per main stem, number of seeds per plant and seed yield per plant, respectively (Table 4). However, heterosis percentages relative to the better parents (BP) were significantly positive in one, five, six and eight crosses with a range of 19.0-19.0; 54.2-73.7; 62.9-97.7 and 47.6-129.9% for plant height, number of pods, number of seeds per plant and seed yield per plant, respectively. Based on the two estimates of heterosis percentage, one cross (P3xP6), five crosses (P3xP5, P3xP6, P3xP7, P5xP7 and P6xP7), six (P1xP6, P3xP5, P3xP6, P3xP7, P3xP8 and P5xP6) and eight crosses (P1xP6, P2xP6, P2xP7, P3xP5, P3xP6, P3xP7, P4xP5 and P5xP6) exhibited significantly positive heterotic effects over both mid and better parents for plant height, number of pods per plant, number of seeds per plant and seed yield per plant. None of the tested crosses significantly exceeded its parents for 100-seed weight. With respect to days to flowering, seven crosses exhibited significantly negative heterosis compared to the respective mid-parents and recorded a range of -9.0 to -23.5%. It could be suggested that the heterotic effects for seed yield was associated with other components.
| Table 4: | Heterotic percentage of F1 relative to mid (MP) and better (BP) parents for different studied traits |
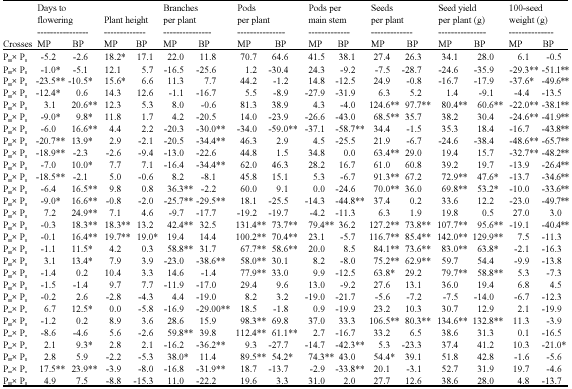 | |
| *,**Significant at 0.05 and 0.01 levels of probability, respectively | |
Moreover, various cross combinations exhibited different degrees of F1 superiority i some traits based on the genes in parental combinations that may contribute directly or indirectly to the characters. Different values of heterosis might be due to the genetic diversity of the parents with non-allelic interaction which increase or decrease the expression of heterosis (Hayman, 1958). The heterosis estimates for the majority of the traits indicate that there was sufficient genetic divergence among the parents assessed, resulting in a favorable situation for breeding (Barelli et al., 2000). Even in the absence of epistasis, multiple alleles at a locus lead to either positive or negative heterosis (Cress, 1966). Pronounced and favourable heterosis have been obtained by several researchers for faba bean traits which varied according to the cross combinations and traits (Duc, 1997; Stelling, 1997; Schill et al., 1998; Abdulmula et al., 1999; Bond and Crofton, 1999; Filippetti et al. ,1999; Abdalla et al., 2001; Attia et al., 2002; Darwish et al., 2005; Attia and Salem, 2006; El-Hady et al., 2006; Gasim and Link, 2007; Ghaouti and Link, 2008; Link et al., 2008).
Combining Ability
Mean squares of both GCA and SCA estimates were highly significant (or significant) in both generations for all the studied traits. Moreover, the variances due to GCA were larger than those for SCA for all variables except for seed yield per plant in F1 generation (Table 5). The ratio of o2gi/o2si estimates exceeded the unity for number of pods per plant (in F2), number of pods per main stem (in F1) and 100-seed weight (in both generations). This indicates that most of the genetic variation among the investigated genotypes for the mentioned traits appears to be under additive gene actions. A direct selection could thus be useful for improving these traits. However, low o2gi/o2si ratios (less than unity), revealed the predominance of non-additive gene action for days to flowering, plant height, number of branches per plant, number of seeds per plant and seed yield per plant. It could be concluded that both the additive and dominance components seemed to have an important role in controlling operating the inheritance of the studied traits, although the contribution of each component bean faba varied according to trait and generation.
| Table 5: | Mean squares due to genotypes, general (GCA), specific (SCA) combining ability and ratios of additive (σ2g) to non additive (σ2s) gene effects for yield and some of its components |
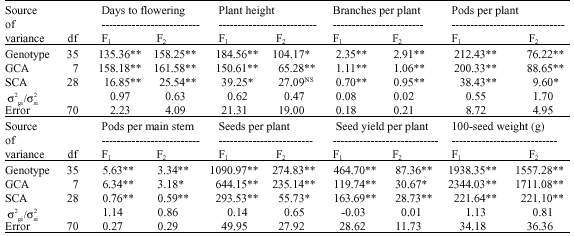 | |
| *,**Significant at 0.05 and 0.01 levels of probability, respectively | |
| Table 6: | General combining ability effects (^gi) for various traits |
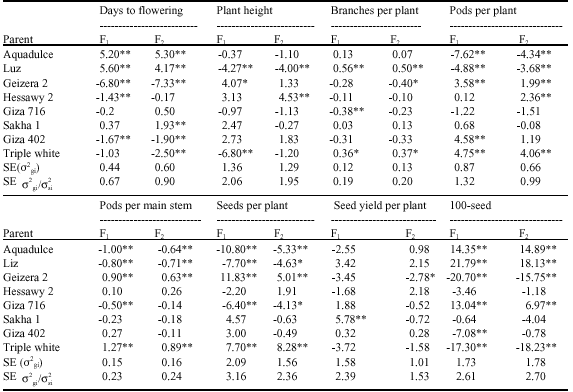 | |
| *,**Significant at 0.05 and 0.01 levels of probability, respectively | |
These findings are coherent with that of Bakheit et al. (2002), where they compared top test cross and six population mating designs and found that both of additive and non additive gene effect were significant for most of the studied traits. Comparisons between GCA effects (σ2gi) associated with each parent (Table 6), revealed that the parent P1 (Aquadulce) and P5 (Giza 716) showed highly significant positive (σ2gi) effects for 100-seed weight, whereas, P2 (Luz) had positive (σ2gi) effects for number of branches per plant and 100-seed weight. Moreover, P3 (Geizera 2) and P8 (Triple white) showed highly significant positive (σ2gi) effects for number of pods per plant, number of pods per main stem and number of seeds per plant. On the other hand, the three parents: P3 (Geizera 2), P7 (Giza 402) and P8 (Triple white) showed negative (σ2gi) effects for days to flowering and could be considered as sources for earliness in breeding program.
| Table 7: | Estimates of specific combining ability effects (σ2Sij) |
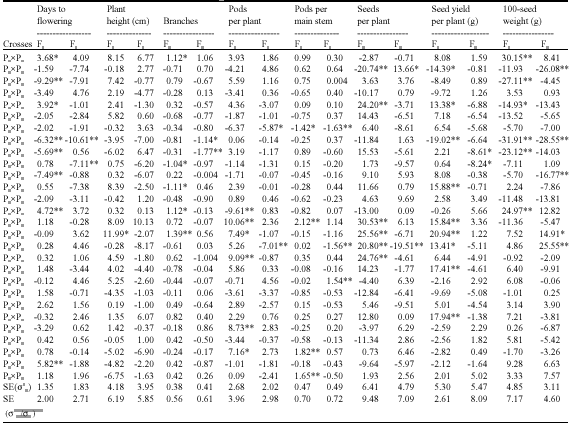 | |
| *,**Significant at 0.05 and 0.01 levels of probability, respectively | |
This findings is highly important for breeding early faba bean cultivars to be grown during the cool season in Soudi Arabia. These results suggest that the mentioned parental genotypes were good combiners for improving most studied traits. The significant relation between combining ability results and the mean performance of parental genotypes indicates the efficiency of phenotypic performance for detecting the potentiality of parents for inclusion in cross breeding programs. Four F1’s (P1xP4, P2xP3, P2xP4 and P2xP6) as well as two F2’s (P2xP3 and P2xP5) had highly significant negative (σ2sij) effects for days to flowering (Table 7). Only one F1 (P3xP6) exhibited significant positive (σ2sij) for plant height. Moreover, three F1’s (P1xP2, P3xP4 and P3xP6) showed significant positive (σ2sij) effects for number of branches per plant (Table 7). With respect to number of pods per plant, five F1’s (P3xP5, P3xP6, P3xP8, P5xP7 and P6xP7) had significant positive (σ2sij) effects. Regarding number of pods per main stem, three crosses (P3xP5, P6xP7 and P7xP8) showed significant positive (σ2sij) effects. Five F1’s (P1xP6, P3xP5, P3xP6, P3xP7 and P3xP8) and only one F2 (P1xP3) had significant positive (σ2sij) effects for number of seeds per plant. Whereas, two F1’s (P1xP2 and P3xP4) along with two F2’s (P3xP6 and P3xP7) showed significant positive (σ2sij) effects for 100-seed weight. Moreover, seven F1’s (P1xP6, P2xP7, P3xP5, P3xP6, P3xP7, P4xP5 and P5xP6) possessed significant positive (σ2sij) for seed yield per plant. Thus SCA for seed yield per plant seemed to be influenced by SCA for yield components.
It is evident from the results that some yield components are more important for yield expression than others. In the selection program, however adjustments up to the desired levels of each component may have to be made in order to obtain the maximum seed yield potential. GCA effects seemed to provide appropriate criterion for detecting the validity of a line in hybrid combination (or synthetic variety) but SCA effects may be related to heterosis (Peng and Virmani, 1999). It seemed that GCA effects were generally unrelated to SCA values of their corresponding crosses. In a cross showing high SCA, it might include only one good combiner, such combinations would show desirable transgressive segregations, providing that the additive genetic system present in the crosses are acting in the same direction to reduce un-derisible plant characteristics and maximize the characters in view (Abdalla et al., 1999). Therefore, most of the earlier crosses may be of importance in traditional breeding programs. These results are in full agreement with Abdalla et al. (2001), Attia et al. (2002), Zeid (2003), Darwish et al. (2005), Attia and Salem (2006), El-Hady et al. (2006) and El-Hady et al. (2007).
In conclusion, the present results are highly promising to breed faba bean cultivars, hybrids or synthetics possessing genetic factors for earliness and high yield potential. This findings is valuable to utilize the cool season before the hot dry season.
REFERENCES
- Abdelmula, A.A., W. Link, E.V. Kittlitz and D. Stelling, 1999. Heterosis and inheritance of drought tolerance in faba bean (Vicia faba L.). Plant Breed., 118: 485-490.
CrossRefDirect Link - Aouar-Sadli, M., K. Louadi and S. Doumandji, 2008. Pollination of the broad bean (Vicia faba L. var. major) (Fabaceae) by wild bees and honey bees (Hymenoptera: Apoidea) and its impact on the seed production in the Tizi-Ouzou area (Algeria). Afr. J. Agric. Res., 3: 266-272.
Direct Link - Bakheit, B.R., M.Z. El-Hifny, M.M. Eissa and S.B. Ragheb, 2002. Triple test cross and six population techniques for partitioning the components of genetic variance in faba bean (Vicia faba). J. Agric. Sci., 139: 61-66.
Direct Link - Barelli, M.A.A., M.C.G. Vidigal, A.T. Junior, S.P.V. Filho and C.A. Scapim, 2000. Combining ability among six common bean cultivars adapted to the north west of Parana state, Brazil. Bragantia Campinas, 59: 159-164.
Direct Link - Bhatt, G.M., 1971. Heterotic performance and combining ability in a diallel cross among spring wheats (Triticum aestivum L.). Aust. J. Agric. Res., 22: 359-360.
CrossRefDirect Link - Cress, C.E., 1966. Heterosis of the hybrid related to gene frequency differences between two populations. Genetics, 53: 269-274.
Direct Link - Filippetti, A., G.H. Azadegan and C.D. Pace, 1999. Breeding strategies for seed protein content and trypsin inhibitors inferred from combining ability and heterosis in test-crosses of Vicia faba. Plant Breed., 118: 411-416.
CrossRefDirect Link - Ghaouti, L. and W. Link, 2008. Local vs. formal breeding and inbred line vs. synthetic cultivar for organic farming: Case of Vicia faba L. Field Crops Res.
Direct Link - Griffing, B., 1956. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci., 9: 463-493.
Direct Link - Haciseferogullari, H., I. Gezer, Y. Bahtiyarca and H.O. Menges, 2003. Determination of some chemical and physical properties of Sakiz faba bean (Vicia faba L. var. major). J. Food Eng., 60: 475-479.
CrossRefDirect Link - Kunkaew, W., S. Julsrigival, C. Senthong and D. Karladee, 2006. Estimation of heterosis and combining ability in azukibean under highland growing condition in Thailand. Chiang Mai Univ. J., 5: 163-168.
Direct Link - Link, W., C. Balko and F.L. Stoddard, 2008. Winter hardiness in faba bean: Physiology and breeding. Field Crops Res.
Direct Link - Peng, J.Y. and S.S. Virmani, 1999. Combining ability for yield and four related traits in relation to breeding in rice. Oryza, 27: 1-10.
Direct Link - Stelling, D., 1997. Heterosis and hybrid performance in topless faba beans (Vicia faba L.). Euphytica, 97: 73-79.
CrossRefDirect Link - Suso, M.J., M.T. Moreno and A.E. Melchinger, 1999. Variation in outcrossing rate and genetic structure on six cultivars of Vicia faba L. as affected by geographic location and year. Plant Breed., 118: 347-350.
CrossRefDirect Link - Tamas, E., M. Savatti and R. Sestras, 1998. Some biological aspects in Vicia faba L. Not. Bot. Horti Agrobot. Cluj-Napoca, 28: 17-19.
Direct Link
Teshome Legese Ya'e Reply
your paper is very appreciable having to date information to the resoure poor like me. So it is better if accessed freely online.
salem
ok u will access the paper freely online
bye
salem
Teshome Legese Ya'e Reply
, I am working in one of the agriculture research in Eth. on highland pulse and oil crops improvement.
salem Reply
sure Eth. is great place for most legumose. faba bean is a common crop over there.
good luck
salem
Paul Gibson
Salem--
I'd love to get your email address. I was happy to see this article--one of my students at Makerere Univ in Uganda (where Pauline and I are now) cited your article. I've thought about you often, and wanted to get back in touch.
Sincerely,
Paul
Samir Rabie El Sayed Abo-Hegazy Reply
* 2004-2006 is not winter season,, if winter season, it have to be from 2004/2005 to 2006/2007
** why he did not use Vicia faba in the abestract??? because this a breeding papper
*** somtimes writing et al., and sometimes et al. only??
M.K.AL-Kummer Reply
We are in iraq grow faba bean in a very large areas all roun the country .yet we have limited reseach work
on faba bean breeding Programme,therefore this work is a very helpfull for post graduate student proposal.
If you can send a copu to us.
With best regards
Prof.dr.Majid AL-Kummer
University of Mosul
IRAQ