
F.J. Al- Saffar
Department of Anatomy, Head of Histology Section/College of Veterinary Medicine, Baghdad University, Iraq
LiveDNA: 964.4860
ORCID: 0000-0003-3447-5736
Asian Journal of Cell Biology
Year: 2015 | Volume: 10 | Issue: 3 | Page No.: 66-79







ABSTRACT
The study aimed to assess histology of rat kidneys and explore the renal oxidative stress post treatment with zerumbone via evaluation of some renal metabolizing enzymes and assay of prostaglandin E2 in the serum. To conduct such research, fifty adult male Sprague Dawley rats were randomly assigned into five equal groups. Rats in the first and second groups were administered 2 mL kg–1 body weight of 0.2 and 0.4% w/v zerumbone, respectively. Third group were treated with Celecoxib and served as positive control and rats in the fourth group were received corn oil and served as negative control whereas rats of the fifth group were left without treatment as a basal normal group. Microscopic findings revealed normal picture of the renal tissue and no signs of morphological changes. Biochemical analysis showed that the microsomal content of Cytochrome P450 was significantly induced in the first three groups compared to fourth and fifth groups. Activity of cytosolic glutathione-S-transferase enzyme was induced significantly in zerumbone treated groups. Non significant induction of total glutathione enzyme in the animals treated with zerumbone or celecoxib was detected. Similarly, there was no significant elevation in the level of malondialdehyde in the first three groups compared to fourth and fifth groups. Significant reduction of prostaglandin hormone following four weeks of oral administration of zerumbone and celecoxib compared to the negative control group was observed. Interestingly the levels of hormone in second group showed comparable level to those measured in the normal group.
PDF Abstract XML References Citation
Received: July 17, 2015;
Accepted: August 29, 2015;
Published: October 06, 2015
How to cite this article
F.J. Al- Saffar, 2015. Effects on Renal Histology, Renal Metabolizing Enzymes and Oxidative Stress Biomarkers in Rats Administered Zerumbone Against Osteoarthritis. Asian Journal of Cell Biology, 10: 66-79.
DOI: 10.3923/ajcb.2015.66.79
URL: https://scialert.net/abstract/?doi=ajcb.2015.66.79
DOI: 10.3923/ajcb.2015.66.79
URL: https://scialert.net/abstract/?doi=ajcb.2015.66.79
INTRODUCTION
Plants are considered nature’s remedies and they have been used throughout the world since ancient times as food, drugs and remedies against various diseases and disorders (Geethalakshmi et al., 2010; Aisha et al., 2011; Pandey et al., 2011; Padam et al., 2012). The study of the extracts or compounds obtained from the rhizomes of the plants attracted several medicinal investigators. Zingiber zerumbet Smith or wild ginger is one of such herbal candidate, widely cultivated in the tropical and subtropical countries for its medicinal efficiencies (Al-Zubairi et al., 2010). The rhizomes of wild ginger are frequently employed in folk medicine against inflammation, diarrhea, rheumatic pain and its young shoots and inflorescence are used as food condiments (Prakash et al., 2011). Several researches documented that the rhizomes of Zingiber zerumbet Smith consist large amount of an oily hydrocarbon compound called zerumbone (Bhuiyan et al., 2009; Abdelwahab et al., 2012; Chen et al., 2011).
Extensive studies were carried out through the last decade on zerumbone anti-inflammatory (Sulaiman et al., 2010), anti-oxidant (Ibrahim et al., 2010) and anti-cancer efficiencies (Al-Zubairi et al., 2010). Previously it was established that the dietary administration of zerumbone can suppress colon carcinogenesis due to its property to suppress the expression cyclooxygenase-2; an enzyme enhances the cell proliferation of the colonic mucosa (Tanaka et al., 2001). Previous researches dealt with various human colonic adenocarcinomas were showed suppressive effect of zerumbone against inflammatory and oxidative stress parameters that are free radical generations, inducible nitric oxide synthase expression and tumor necrosis factor-α release (Murakami et al., 2002). Recently, the immunoreactivity of the synovial membranes and damaged articular surfaces of the knee joint were improved post oral administration of zerumbone against experimental knee osteoarthritis in the rats (Al-Saffar et al., 2010, 2011a).
The use of natural products from the herbs has gained an interest among the world’s population which led to the development of herbal supplements to reduce symptoms of the diseases and to improve the quality of life. Unfortunately, some adverse effects may rise besides their therapeutic efficiencies (Foster et al., 2002). Therefore, the determination and evaluation of some metabolizing enzymes and oxidative stress biomarkers in the renal tissue or other important vital organs should kept in consideration upon using such herbal or natural products.
Kidneys possess most of metabolizing enzymes against xenobiotics and are thus able to make an important contribution to the body’s metabolism of drugs and foreign compounds. In general, the catalytic activities of the various renal enzymes are lower than those of the liver but with some exceptions, such as; enzymes involved in the processing of glutathione conjugates (Lock and Reed, 1998). Oxidative stress of the xenobiotics may influence the metabolism of cells in vital organs such as kidney, heart, liver and nervous tissue (Mandelker, 2008). Xenobiotics such as drugs, chemical carcinogens, pesticides, environmental pollutants, ingested natural products are undergoing a number of hepatic and extra-hepatic enzymatic reactions referred to as biotransformation, which may be categorized into phase I and phase II reactions. Metabolizing enzymes first activates xenobiotics and then conjugate their active secondary metabolites with either glucuronic or sulfuric or glutathione and subsequently excretes them in the bile or urine (Prakash and Vas, 2008). However, biotransformation may either detoxify the compounds, or may produce more toxic intermediate metabolites than the parents compounds themselves (Lechon-Moliner and Donato, 2007; Malhi et al., 2010). In fact, the effect of oxidative stress is hazardous as it does not show any symptoms so that its detection is really difficult by means of the common laboratory procedures (Kataria et al., 2010).
Cytochrome P450 (CYP450) isoenzymes are superfamily of hemoprotein enzymes found on the membrane of endoplasmic reticulum. They are considered main metabolizing enzymes because they are responsible for about 75% of phase I dependent drug metabolism (Ingelman-Sundberg, 2004; Budriang et al., 2011; Sirisangtrakul and Sripanidkulchai, 2011; Wangcharoenrung and Warisnoicharoen, 2011). These isozymes catalyze most reactions of phase I through the oxidation, hydroxylation and demethylation of their substrates (Kalra, 2007). They render foreign compounds more water-soluble and thus facilitate their excretion (Mandelker, 2008). The inducibility of the CYP450s by a large number of compounds is an important property because it enables the living organism to detoxify a wide range of exogenous xenobiotics. Paradoxically, induction of these enzymes have potential risk factor because many of their reactions may activate the procarcinogens to their critical carcinogenic structure and make them fit to interact with the nucleophilic sites in the DNA (Koul et al., 2009). If the metabolites of phase I reactions are sufficiently polar, they may be readily excreted at this point. However, many products of phase I reactions are not eliminated rapidly and undergo a subsequent reaction of the phase II in which an endogenous substrate combines with the newly incorporated functional group to form a highly polar conjugate.
Glutathione S-Transferase (GST), which is one of the important phase II metabolizing enzymes is a large family of isozymes, play a role in intracellular transport of hydrophobic molecules through their conjugation with the reduced form of the intracellular glutathione (Korkina et al., 2009). Large number of toxic xenobiotics conjugates glutathione producing inactive glutathione conjugates and such conjugates have higher molecular weights and being more water soluble and more likely excreted in the urine and bile than the parent compounds themselves (Zhu et al., 1998). Failure of these toxic xenobiotics to be conjugated with glutathione left them free to attack the cellular DNA, RNA and proteins (Abraham and Singh, 1999). The reduced form of glutathione and GSTs are able to protect the cells from the harmful free reactive intermediate compounds by capturing them before they attack the vital nucleophilic sites of the active cells. Glutathione S-transferases are found in highest concentration in the kidney, liver, intestine and lungs. The cytosolic GSTs, which comprise over 95% of total cellular GSTs are the major isoforms involved in xenobiotics metabolism. Fundamentally, these enzymes catalyses the nucleophilic attack of the sulphur atom of glutathione on the electrophilic group of xenobiotics and ultimately decreases their reactivity and render them more water soluble so that facilitate their subsequent removal from the body (Sau et al., 2010).
Malondialdehyde (MDA) is a relevant indicator or biomarker for lipid peroxidation which is enhanced in the presence of oxidative stress in the living organisms. Its concentrations are elevated in different diseases due to the damage caused by the generated free radicals species. Hence, its determination is considered the most common approach for lipid peroxidation and evaluation of the oxidative stress (Ediz et al., 2011). Polyunsaturated fatty acids of the cell membranes are very susceptible to the free radical attack and eventually form many stable products such as lipid hydroperoxides, lipid peroxides, hydrocarbons and aldehyde, which are implicated in many pathological disorders such as atherosclerosis, cancers and diabetes (Chaudhary et al., 2007). Malondialdehyde is frequently determined spectrophotometrically after its reaction with thiobarbituric acid at high temperature and low acidic media and measuring the absorbance of reaction mixture at 532 nm (Lefevre et al., 1998). Many previous investigations on the oxidative stress established significant raise of lipid peroxidation, whereas, dissimilarly, glutathione and other endogenous enzymatic antioxidants are depleted, indicating strong correlation between these two oxidative stress biomarkers (Idonije et al., 2011; Nagy et al., 2011; Rahimi et al., 2012). The inhibitory outcome of antioxidants against lipid peroxidation was thought to be performed by either preventing the accumulation of reactive oxygen species or by preventing the peroxidation chain reaction (Choi and Cho, 2009; Karafakioglu et al., 2011).
Glutathione (GSH) antioxidant is an ubiquitous molecule present in all cellular parts where it conducts various functions ranging from the detoxification to the protection of cells from the damage of oxidative stress. Antioxidant property of GSH is due to its thiol group, which is reducing agent that can be reversibly oxidized and reduced. It protects the cell membranes from the oxidative damage by its ability to remove the produced and accumulated reactive free species (Dehghan et al., 2010). The level of GSH in the tissue is primarily dependent on the rate of biosynthesis and utilization in oxidation/reduction reactions (Anand et al., 2011). Cellular glutathione is usually maintained in its reduced form by the glutathione reductase and in turn it reduces other metabolites and enzymes systems such as the ascorbate in the glutathione ascorbate cycle, glutathione peroxidase and can react directly with oxidants (Foyer and Noctor, 2011; Long et al., 2012). Oxidants such as carbon tetrachloride and Aminoglycoside antibiotics can cause renal oxidative damage because it induces lipid peroxidation and oppositely, it reduces the levels of the renal GSH and other enzymatic antioxidants (Adaramoye, 2009; Kandeel et al., 2011). Antioxidants that acts against oxidants are either enzymatic such as superoxide dismutase, catalase and GSH peroxidase or non enzymatic such as selenium, zinc, uric acid and vitamin C and E (Shuid et al., 2011; Hosseini and Abdollahi, 2012). Antioxidants are beneficial health substances because of their vital role against different diseases and disorders (Budhiyanti et al., 2011; Prasong, 2011; Saboo et al., 2011). Cellular concentration of glutathione may decreased in association with the processes of aging and pathogenesis of many diseases (Mahmoudabad et al., 2008) and on another hand, its level may affected by the growth, nutritional status and hormonal/stress levels (Ballatori et al., 2009). To our knowledge, there is no research conducted before to detect the oxidative stress on kidney tissue associated with osteoarthritis regulated with zerumbone administration. Currently the study aimed to explore the effect of oral application of zerumbone on renal oxidative stress enzymes such as microsomal Cytochrome 450, cytosolic glutathione-S-transferase and oxidative stress biomarkers such as glutathione and malondialdehyde.
MATERIALS AND METHODS
Experimental design: The study was performed on fifty adult male Sprague Dawley rats randomly assigned into five groups (n = 10). The rats were individually caged in air-conditioned animal room at 22°C and were given commercial pellet and tap water ad libitum. To induce osteoarthritis (OA), rats were anesthetized and injected intraarticularly with 50 μL of monosodium iodoacetate (MIA) diluted with saline at a concentration of 60 mg mL–1 (Sigma, USA) in their right knee joints at day 0 (Al-Saffar et al., 2009).
Zerumbone preparation and protocol of treatment: Preparation of zerumbone was implemented and well described according to the previous publication (Al-Saffar et al., 2010). Oral treatment was performed daily starting on day 16 using feeding catheter and lasted on day 43 according to the following protocol. Rats in the first (ZI) and second (ZII) groups were administered 2 mL kg–1 body weight of 0.2 and 0.4% w/v zerumbone in corn oil, respectively. Third group (CEL) were treated with celecoxib in a dosage of 30 mg kg–1 body weight diluted in 5% carboxyl methyl cellulose and served as a positive control and rats in the fourth group (CO) were received corn oil in a dosage of 2 mL kg–1 body weight and served as a negative control whereas rats of the fifth group (B) were left without treatment as a basal normal group.
Prostaglandins assay and kidneys collection: Upon euthanasia, 5 mL of blood was collected from rats to estimate proinflammatory prostaglandin (PG) E2 concentrations at three different periods that are before OA induction, end of OA induction and end of the treatment periods. Blood was collected through the cardiac puncture and left for one hour then centrifuged at 3000 rpm for 10 min. Sera were collected and stored at -20°C until further use. Enzyme immunoassay kit for PGE2 detection (Assay Design purchased from USA, Catalog No. 900-001) was used to perform hormone assays. Thereafter, rats were dissected and kidneys were removed for both histological and toxicological examinations. For general morphological examination, kidneys were fixed in 10% neutral phosphate-buffered formalin solution. Following dehydration in an ascending series of ethanol (70, 80, 96, 100%), tissue specimens were cleared in xylene and embedded in paraffin. Tissue sections of 6 μm were stained with hematoxylin and eosin (H and E). Light microscopy was used to detect and evaluate glomerulus or tubular changes in the renal tissue that may occur.
Biochemical analysis: Determination of protein in the renal microsomal pellet, cytosolic fraction and kidney homogenate was assayed using BCATM protein assay kit (Thermo Scientific, USA), which was performed according to the manufacturer’s instructions.
Preparation of renal microsomal pellet and cytosolic fraction were prepared according to the method previously described (Al-Saffar et al., 2011b). Upon euthanasia, kidneys were excised and washed in cold 1.15% KCl then one gram of each kidney was mixed with 10 times to its weight with 1.15% KCl, minced well and homogenized thoroughly, centrifuged at 104 g for 20 min. A volume of buffer A [0.10 M Tris-acetate buffer (pH 7.4) containing 0.10-M KCl, 1.0 mM Ethylenediamine Tetraacetic Acid (EDTA) and 20 mM Butylated Hydroxytoluene (BHT)] equal to the discarded sediment was added to the supernatant and centrifuged for 60 min at 105 g (Beckman, XL-80 ultracentrifuge, 70.1 Ti rotor) to yield supernatant (contain the cytosolic fraction which was saved for subsequent uses) and a microsomal pellet. A volume of buffer B (0.10 M potassium pyrophosphate buffer (pH 7.4) containing 1.0 mM EDTA and 20 mM BHT) equal to that saved supernatant was added to the pellet and suspended, homogenized and recentrifuged at 105 g for 60 min. The resulted pellet was homogenized in 2 mL of buffer C (10 mM Tris-acetate buffer (pH 7.4) containing 1.0 mM EDTA and 20% glycerol (w/v)) and stored at -80°C.
Determination of CYP P450 was conducted according to the method of Omura and Sato (1964). Aliquot from microsomal pellet was diluted with 0.05 M PBS (pH 7.4) to obtain sample contain protein concentration approximately 2 mg mL–1. Aliquot of 2 mL from this sample was transferred for each reference and sample cuvettes. Their baseline densities were determined using a spectrophotometer by scanning them from 420-490 nm. Carbon monoxide was bubbled gently into the sample cuvette for 20 sec and then a small pinch of powdered sodium dithionite was added. The cuvette was covered with a parafilm and inverted many times and then bubbled with carbon monoxide again for 20 sec. The reference cuvette was treated only with a small pinch of sodium dithionite. The cuvettes were shaken vigorously and their densities were determined with the spectrum 450 nm. The quantity of CYP450 was calculated from the differences of Optical Densities (OD) and the molar extinction coefficient (91 mM–1 cm–1) as in the following equation:
*amount of CYP 450 per each 1 mg of protein of microsomal pellet.
Determination of cytosolic GST activity was implemented according to the method of Habig et al. (1974). The procedure was conducted by the preparation of two cuvettes (sample and reference) each of 3 mL from the followings: 2.2 mL of 0.1 M potassium phosphate buffer (pH 6.5), 0.1 mL of 30 mM CDNB substrate (1-chloro-2, 4-dinitrobenzen from Aldrich Chemical Co.) and 0.1 mL of 30 mM GSH. Both cuvettes were left for 1 min at room temperature (25°C) and read with spectrophotometer at 340 nm. An aliquot of 0.6 mL of the cytosol was added to the sample cuvette and mixed thoroughly. Readings were read after 1, 2, 3, 4 and 5 min for both cuvettes at 340 nm with spectrophotometer. The specific activity of GST was expressed as nmol of conjugate formed min–1 mg–1, which is calculated from the differences of Optical Densities (OD) and the molar extinction coefficient (9.6 mM–1 cm–1) as in the following equation:
Determination of MDA was implemented according to the method of Ohkawa et al. (1979). Homogenates of 10% were prepared from kidneys by mincing 1 g of their tissue in 10 times to this weight with 1.15% KCl. Aliquot of 0.1 mL from the homogenate mixed with 0.2 mL of 8.1% sodium dodecyl sulfate, 1.5 mL of 20% acetic acid solution (pH 3.5) and 1.5 mL of 0.8% aqueous solution of thiobarbituric acid. The mixture was finally made up to 4.0 mL with distilled water and heated at 95°C for 60 min. After cooling with tap water, 1.0 mL of distilled water and 5.0 mL of n-butanol were added and the mixture was shaken vigorously and centrifuged at 4000 rpm for 10 min, the upper butanol layer was separated and read with spectrophotometer at 532 nm against pure n-butanol. The concentration of MDA was determined by comparing the absorbance to that of the standard curve with a range from 1-100 nM of authentic MDA (product of Sigma Co.) and expressed as μ mol mg–1 of protein.
Determination of total GSH concentration was measured in the kidney homogenate according to the method of Sedlak and Lindsay (1968) which is well described by Alsaif (2009). About 2 mL of 10% trichloacetic acid was added to a 2 mL of homogenate and centrifuged at 600 g for 15 min. Then 2 mL of supernatant was added to 2.0 mL of 0.32 mol L–1Na2HPO4. About 5 min prior to the measurement of absorbance, 50 μL aliquot of 0.01 mol L–1 Ellman’s solution (5, 5-dithiobis-2-nitrobenzoic acid) was added. The absorbance was determined at 412 nm. The concentration of GSH was determined by comparing the absorbance to that of the standard curve with a range from 1-100 nM of authentic GSH (product of Sigma Co.) and expressed as μ mol mg–1 of protein.
Statistical analysis: Statistical calculations were carried out with the SPSS 15.0 for windows software package. Data was expressed as Mean±SEM and analyzed with one way ANOVA and confirmed with student t-test (Table 1). The comparisons were considered significant if p<0.05.
| Table 1: | Renal concentrations of proteins, CYP450, GST, GSH, MDA and PGE2 in ZI (treated with 2 mL kg–1 b.wt. of 0.2% w/v zerumbone diluted in corn oil), ZII (treated with 2 mL kg–1 b.wt. of 0.4% w/v zerumbone diluted in corn oil), CEL (treated with 30 mg kg–1 b.wt. diluted in 5% carboxyl methyl cellulose), CO (treated with 2 mL kg–1 b.wt. corn oil) and B (normal untreated) groups |
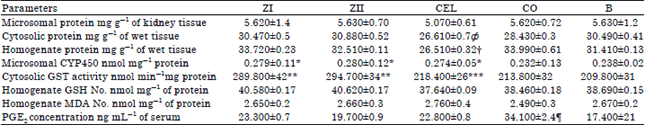 | |
Ф lowest cytosolic protein post treatment with the Celecoxib, *Higher microsomal CYP450 in treated groups with zerumbone and Celecoxib than, untreated and normal groups, **Higher cytosolic GST activity at groups treated with zerumbone than other groups, ***Lower cytosolic GST activity at groups treated with Celecoxib than the zerumbone, treated groups, but nearly equal to those untreated and normal groups, NO: Non significant differences between all groups, ¶Higher elevation of PGE2 concentration in the group with induced osteoarthritis and no treated than the other groups | |
RESULTS
Clinical and gross observations: Clinically, rats in the corn oil treated group were showed swelled right knee joints accompanied with signs of lameness, whereas rats in the other treated groups showed unapparent joint swelling with no signs of abnormal gait. Food and water consumption of rats allocated in ZI, ZII and CEL but not CO groups was similar to those of the normal Basal (B) group during the course of treatment. Body weight gain of rats was measured starting on day 0 until the day 43 which was the day of rat’s euthanasia (Fig. 1). Body weight was dropped suddenly following OA induction in all rats except those in the normal Basal (B) group which was left without OA induction and was untreated. Three days later on, the mean body weight sustained to that before OA period. Following the treatment with zerumbone the body weight showed gradual gain which was significantly comparable to those of the normal basal group. The gain of body weight was insignificantly lower in celecoxib treated group and significantly lower in corn oil treated group compared to those of the basal group. Upon euthanasia, kidneys dissected from all treated groups were showed normal size and coloration compared to those of the normal basal group. All kidneys were found glistening, pale brown to reddish in color and approximately 12-15 g weights with absolute absence of gross degenerative changes.
Microscopic examination of kidney: Light microscopy revealed no signs of glomerular or renal tubular degeneration in the renal cortex and the absence of abnormal histology picture in the medullary tissue. Current findings revealed total absence of vascular changes and there were no inflammatory cells infiltration in the interstitial tissues of the different examined parts of the renal tissues. Both proximal and distal renal tubules showed clear lumina with total absence of a myeloid formation with no signs of tubular necrotic changes in the cortices of all kidneys of rats incorporated in this study (Fig. 2), except those of CEL group where mild cellular changes in the proximal convoluted renal tubules were observed. The lining epithelium of these tubules showed loss of brush border and some pyknotic and necrotic nuclei.
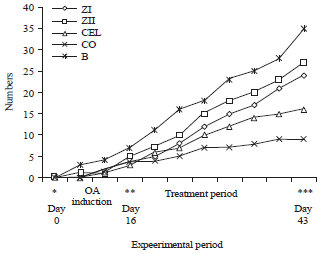 | |
| Fig. 1: | Body weight gain in gram (No.) throughout the experimental period. Obvious loss of body weight was observed on day 0 [the day of osteoarthritis (OA) induction] in all groups except the normal basal group (B). During the treatment period the weight gain in zerumbone treated groups (ZI and ZII) was comparable to the normal basal group (B), whereas, it was lower insignificantly (p>0.05) in celecoxib (CEL) but lower significantly (p<0.05) in Corn Oil (CO) treated groups |
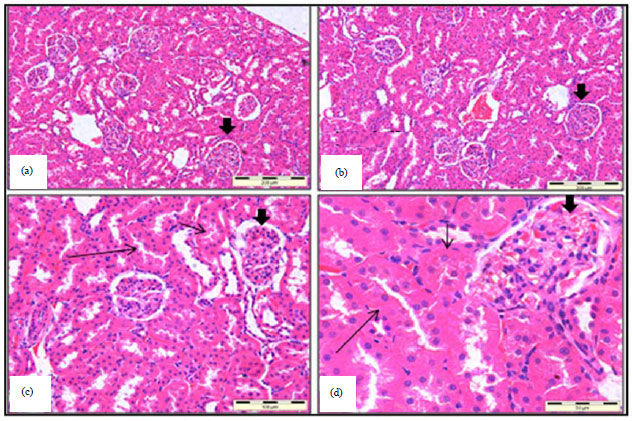 | |
| Fig. 2(a-d): | Histological sections from (a) ZI, (b) ZII, (c) CO and (d) B groups showed normal pictures of Glomeruli (thick arrows), proximal convoluted tubules (short thin arrows) and distal convoluted tubules (long thin arrows), Hematoxylin and Eosin |
Results of biochemical analysis: Renal protein assay, content and activities CYP450 and GST enzymes, levels of GSH and MDA oxidative stress biomarkers in different renal tissue preparations as well as serum PGE2 concentration were statistically analyzed and summarized in Table 1. Assay of protein showed no significant differences in microsomal pellet concentration between the designated groups in this study. While concentration of protein in both cytosolic fraction as well as renal homogenate revealed significant lowering in those of CEL group versus the others. Protein determined in the kidney homogenate was higher than the cytosolic fraction and both were significantly higher than the microsomal pellets content. The microsomal content of CYP450 was significantly induced in ZI, ZII and CEL groups compared to CO and B groups. Activity of cytosolic GST enzyme was significantly induced in ZI and ZII groups too but not in CEL group, which was not significantly affected as compared to either CO or B groups.
Determination of total GSH in the kidney homogenates of the different groups revealed no significant induction of this antioxidant enzyme in the animals from ZI, ZII or CEL groups compared to CO and B groups. Similarly, there was no significant elevation in the level of MDA in ZI, ZII and CEL groups compared to CO and B groups.
Assay of the inflammatory PGE2 showed a significant reduction of this hormone following four weeks of oral administration of zerumbone in ZI, ZII and CEL compared to the CO. Interestingly the levels of PGE2 in ZII group showed comparable level to those measured in the B group.
DISCUSSION
The current study intended to explore the status of renal oxidative stress post oral administration of zerumbone to recommend longer course or higher dosage regimen of this interesting remedy to alleviate OA or other diseases without possible unfavorable side effects. The present work is, to our knowledge, the first study that reports the in vivo the effect of zerumbone on renal metabolizing enzymes and oxidative stress biomarkers. The study provided fundamental information on the responses of the metabolizing enzymes and oxidative stress biomarkers next to the oral application of zerumbone against experimental osteoarthritis in rats because this herbal agent possesses many important medicinal properties; thus it deserves to be well investigated. Regarding this aspect, recently documented plausible chondroprotective effects of zerumbone orally administrated against experimentally induced knee osteoarthritis in rats (Al-Saffar et al., 2010).
Currently, gross and microscopic findings revealed glistening kidneys with normal appearance and absence of renal degenerative changes indicating the absence of renal damage in rats treated with both doses of zerumbone (ZI, ZII). These findings were consonance with those of the bioanalysis measurements in which levels of MDA in the renal tissues were insignificantly changed compared with those levels observed in the normal basal group. Absence of significant elevation in MDA levels indicates absence of lipid peroxidation which in turn indicates absence of oxidative damage post zerumbone oral administration. On other hand, current data showed significant induction of CYP450 and GST enzymes post oral administration of the two different doses of zerumbone (ZI, ZII). The inducibility of these enzymes is an important property because it enables the living organism to detoxify various exogenous compounds (Raza et al., 1998). This inducibility was accompanied with insignificant elevation of renal total GSH antioxidant. The current doses and courses of zerumbone application appeared within the favored therapeutic limit with antioxidant property and lack of the lipid peroxidation. The insignificant differences of MDA concentrations in the kidney tissue indicates that its level was insufficient to cause renal tubular damage or degeneration which reflect absence of lipid peroxidation or oxidative stress in responses to the different doses and course of treatments applied in this study. Previous researches established antioxidant property of zerumbone (Ibrahim et al., 2010). The two different doses of zerumbone used in this study induced the level of antioxidant glutathione in the rat kidney tissue homogenates. Zerumbone induce GSH in the treated groups due to its antioxidant property but the elevation of its level was insignificant compared to the normal basal untreated group regarding the dose of zerumbone administered in this study. Current applied doses of zerumbone indicate that this important medicinal herb can be used in higher concentration in the future for better therapeutic efficiency against some diseases or health disorders. Anti-inflammatory effect of zerumbone was well identified in this study because the levels of PGE2 in the sera of rats treated with zerumbone were significantly inhibited next to the 4 weeks of treatment with this remedial herb.
Quantitative enzyme measurement of the total CYP450 in renal microsome of the normal control and zerumbone treated groups was lower than those observed previously in the liver microsomes (Al-Saffar et al., 2011b). In fact, enzymes activities are tissue dependant and they are varying from tissue to other depending on the duration and severity of the pathological condition (Anand et al., 2011). Microsomal CYP450 levels in the current findings were in a consonance with the previous investigations in the rat kidney tissues (Goodin et al., 2000).
Current outcome showed zerumbone inducibility toward CYP450 in rat kidney microsome. The findings showed significant induction of CYP450 enzyme in groups treated with either zerumbone (ZI, ZII) or celecoxib (CEL). The inducibility of this enzyme is an important property because it enables the living organism to detoxify various exogenous compounds (Raza et al., 1998). The induction of this enzyme by the given dosage regimen and course of zerumbone in the current study should be kept in consideration when supplemented with other drugs or therapies against osteoarthritis or other diseases. The premise is based on the probable occurrence of drugs interaction and that the inducibility may enhance the metabolism of these substances, which are substrates to CYP450 or some of its isozymes. Currently, renal cytosolic GST was induced significantly in zerumbone treated groups (ZII, ZII) because of its potential inducibility to some isozymes of GST against lipid peroxidation and subsequently play a role in the protection against oxidative stress (Nakamura et al., 2004). The inducibility of zerumbone toward GST enzyme is in consonance with recently published findings (Abdul et al., 2008).
Nonsteroidal Anti-Inflammatory Drugs (NSAIDs) are the most currently used drugs to alleviate pain and inflammatory symptoms raised from the rheumatic and other musculoskeletal disorders. They are excellent inhibitors for the cyclooxygenase (COX) enzymes which are required for the synthesis of inflammatory prostaglandins (PGs). Celecoxib is deliberated for acute pain, menstrual cramps, pain and inflammation of osteoarthritis and rheumatoid arthritis but its application with the higher doses may cause cellular degeneration in liver and tubular damage in the kidney (Kockaya et al., 2010). Previous investigations postulated that celecoxib exert minor unfavorable effects compared to the other traditional NSAIDs that are inhibitors for both COX-1 and COX-2. However, recent works revealed that prolonged course with higher doses of celecoxib against OA or rheumatic arthritis cause generalized and peripheral edema and hypertension indicating renal dysfunction due to the damages in both cortical and medullary tissues of the kidneys (Zhao et al., 2001).
Actually, celecoxib, a selective cyclo-oxygenase-2 inhibitor so that prevents the formation of prostaglandins which is responsible for maintenance of renal function, accordingly its activity may predispose to the development of renal damage (Gupta et al., 2007). Celecoxib induce significantly lipid peroxidation and the activities of glutathione-S-transferase and on contrary it decreases the reactive thiol and glutathione in the renal tissue, which indicates inversely relationship between these two biomarkers (Sozer et al., 2011). Our results had showed different findings as it exposed neither lipid peroxidation nor renal damage in the kidneys following oral administration of zerumbone or celecoxib.
The present study revealed that the chronic administration of celecoxib led to a significant increase in GST activity. As GST catalyzes the conjugation of glutathione, the increase in GST activity would reduce glutathione levels. Thus present results showing reduced glutathione and increased GST activity corroborate each other (Gupta et al., 2007). Similarly, we found Celecoxib induced GST and reduced GSH.
CONCLUSIONS
The current study aims to explore the status of renal oxidative stress post oral administration of zerumbone to recommend longer course or higher dosage regimen of this interesting remedy to alleviate OA or other diseases without any possibility of adverse side effects.
REFERENCES
- Chen, B.Y., D.P.C. Lin, K.C. Su, Y.L. Chen and C.Y. Wu et al., 2011. Dietary zerumbone prevents against ultraviolet B-induced cataractogenesis in the mouse. Mol. Vision, 17: 723-730.
PubMedDirect Link - Abdelwahab, S.I., A.B. Abdul, Z.N.M. Zain and A.H.A. Hadi, 2012. Zerumbone inhibits interleukin-6 and induces apoptosis and cell cycle arrest in ovarian and cervical cancer cells. Int. Immunopharmacol., 12: 594-602.
CrossRefDirect Link - Abdul, A.B.H., A.S. Al-Zubairi, N.D. Tailan, S.I. Abdel-Wahab, Z.N.M. Zain, S. Ruslay and M.M. Syam, 2008. Anticancer activity of natural compound (Zerumbone) extracted from Zingiber zerumbet in human hela cervical cancer cells. Int. J. Pharmacol., 4: 160-168.
CrossRefDirect Link - Abraham, S.K. and S.P. Singh, 1999. Anti-genotoxicity and glutathione S-transferase activity in mice pretreated with caffeinated and decaffeinated coffee. Food Chem. Toxicol., 37: 733-739.
CrossRefDirect Link - Adaramoye, O.A., 2009. Comparative effects of vitamin E and kolaviron (a biflavonoid from Garcinia kola) on carbon tetrachloride-induced renal oxidative damage in mice. Pak. J. Biol. Sci., 12: 1146-1151.
CrossRefPubMedDirect Link - Aisha, A.F.A., Z.D. Nassar, M.J. Siddiqui, K.M. Abu-Salah, S.A. Alrokayan, Z. Ismail and A.M.S.A. Majid, 2011. Evaluation of antiangiogenic, cytotoxic and antioxidant effects of Syzygium aromaticum L. extracts. Asian J. Biol. Sci., 4: 282-290.
CrossRefDirect Link - Al-Saffar, F.J., S. Ganabadi, H. Yaakub and S. Fakurazi, 2009. Collagenase and sodium iodoacetate-induced experimental osteoarthritis model in sprague dawley rats. Asian J. Scient. Res., 2: 167-179.
CrossRefDirect Link - Al-Saffar, F.J., S. Ganabadi, S. Fakurazi, H. Yaakub and M. Lip, 2010. Chondroprotective effect of zerumbone on monosodium iodoacetate induced osteoarthritis in rats. J. Applied Sci., 10: 248-260.
CrossRefDirect Link - Al-Saffar, F.J., S. Ganabadi, S. Fakurazi and H. Yaakub, 2011. Zerumbone improved immunoreactivity of neuropeptides in monosodium iodoacetate induced knee osteoarthritis in rat. Afr. J. Biotechnol., 10: 3646-3653.
Direct Link - Al-Saffar, F.J., S. Ganabadi, S. Fakurazi and H. Yaakub, 2011. Response of hepatic metabolizing enzymes and oxidative stress in orally administrated zerumbone against MIA-induced osteoarthritis in rats. J. Anim. Vet. Adv., 10: 566-573.
CrossRefDirect Link - Alsaif, M.A., 2009. Beneficial effects of rutin and vitamin C coadministration in a streptozotocin-induced diabetes rat model of kidney nephrotoxicity. Pak. J. Nutr., 8: 745-754.
CrossRefDirect Link - Al-Zubairi, A.S., A.B. Abdul, M. Yousif, S.I. Abdelwahab, M.M. Elhassan and S. Mohan, 2010. In vivo and in vitro genotoxic effects of zerumbone. Caryologia, 63: 11-17.
CrossRefDirect Link - Anand, P., D. Rajakumar, M. Jeraud, A.J.W. Felix and T. Balasubramanian, 2011. Effects of taurine on glutathione peroxidase, glutathione reductase and reduced glutathione levels in rats. Pak. J. Biol. Sci., 14: 219-225.
CrossRefPubMedDirect Link - Ballatori, N., S.M. Krance, S. Notenboom, S. Shi, K. Tieu and C.L. Hammond, 2009. Glutathione dysregulation and the etiology and progression of human diseases. Biol. Chem., 390: 191-214.
CrossRefPubMedDirect Link - Bhuiyan, M.N.I., J.U. Chowdhury and J. Begum, 2009. Chemical investigation of the leaf and rhizome essential oils of Zingiber zerumbet (L.) Smith from Bangladesh. Bangladesh J. Pharmacol., 4: 9-12.
CrossRefDirect Link - Budhiyanti, S.A., S. Raharjo, D.W. Marseno and I.Y.B. Lelana, 2011. Free radical scavenging, metal chelating and singlet oxygen quenching activity of fractionated brown seaweed Sargassum hystrix extract. J. Biol. Sci., 11: 288-298.
CrossRefDirect Link - Budriang, C., P. Rongnoparut and J. Yuvaniyama, 2011. An expression of an insect membrane-bound cytochrome P450 CYP6AA3 in the Escherichia coli in relation to insecticide resistance in a malarial vector. Pak. J. Biol. Sci., 14: 466-475.
CrossRefPubMedDirect Link - Chaudhary, D.P., R.K. Boparai and D.D. Bansal, 2007. Implications of oxidative stress in high sucrose low magnesium diet fed rats. Eur. J. Nutr., 46: 383-390.
CrossRefDirect Link - Choi, E.Y. and Y.O. Cho, 2009. Effect of vitamin B6 deficiency on antioxidative status in rats with exercise-induced oxidative stress. Nutr. Res. Pract., 3: 208-211.
CrossRefDirect Link - Dehghan, A., M. Arabi, S. Nahid and M. Aminlari, 2010. Changes of serum reduced and oxidized glutathione in heat stressed ram. Asian J. Anim. Vet. Adv., 5: 472-477.
CrossRefDirect Link - Ediz, L., H. Ozkol, I. Tekeoglu, Y. Tuluce, E. Gulcu and I. Koyuncu, 2011. Increased oxidative stress in patients with familial Mediterranean fever during attack period. Afr. Health Sci., 11: S6-S13.
Direct Link - Foster, B.C., S. Vandenhoek, K.Y. Li, R. Tang, J.W. Budzinski and A. Krantis 2002. Effect of several Chinese natural health products of human cytochrome P450 metabolism. J. Pharm. Pharmaceut. Sci., 5: 185-189.
Direct Link - Foyer, C.H. and G. Noctor, 2011. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol., 155: 2-18.
CrossRefDirect Link - Geethalakshmi, R., D.V.L. Sarada and P. Marimuthu, 2010. Evaluation of antimicrobial and antioxidant potentials of Trianthema decandra L. Asian J. Biotechnol., 2: 225-231.
CrossRefDirect Link - Lechon-Moliner, M.J.G. and M.T. Donato, 2007. In vitro investigation of drug metabolism and toxicity in man. Anales Real Academia Nacional Farmacia, 4: 5-26.
Direct Link - Goodin, M.G., R. Walker and R.J. Rosengren, 2000. Effect of low concentrations of paracetamol on PGE2 production in the rat kidney. Aust. J. Ecotoxicol., 6: 85-88.
Direct Link - Habig, W.H., M.J. Pabst and W.B. Jakoby, 1974. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem., 249: 7130-7139.
CrossRefPubMedDirect Link - Hosseini, A. and M. Abdollahi, 2012. Antioxidants as an appropriate approach to improve the outcome of pancreatic islet isolation: Evidences from animal studies. Asian J. Anim. Vet. Adv., 7: 540-541.
CrossRefDirect Link - Ibrahim. M.Y., A.B.H. Abdul, T.A.T. Ibrahim, S.I. AbdelWahab, M.M. Elhassan and S. Mohan, 2010. Attenuation of cisplatin-induced nephrotoxicity in rats using zerumbone. Afr. J. Biotechnol., 9: 4434-4441.
Direct Link - Idonije, O.B., O. Festus, O. Okhiai and U. Akpamu, 2011. Comparative study of the status of a biomarker of lipid peroxidation (Malondialdehyde) in patients with Plasmodium falciparum and Plasmodium vivax malaria infection. Asian J. Biol. Sci., 4: 506-513.
CrossRefDirect Link - Kalra, B.S., 2007. Cytochrome P450 enzyme isoforms and their therapeutic implications: An update. Indian J. Med. Sci., 61: 102-116.
CrossRefPubMedDirect Link - Kandeel, M., I. Abdelaziz, N. Elhabashy, H. Hegazy and Y. Tolba, 2011. Nephrotoxicity and oxidative stress of single large dose or two divided doses of gentamicin in rats. Pak. J. Biol. Sci., 14: 627-633.
CrossRefDirect Link - Karafakioglu, Y.S., I.H. Cigerci and A.F. Fidan, 2011. The effects of taurine on aminolevulinic acid dehydratase activity in nonylphenol-induced toxicity. Asian J. Anim. Vet. Adv., 6: 96-100.
CrossRefDirect Link - Kataria, N., A.K. Kataria, R. Maan and A.K. Gahlot, 2010. Evaluation of oxidative stress in brucella infected cows. J. Stress Physiol. Biochem., 6: 19-25.
Direct Link - Kockaya, E.A., G. Selmanoglu, K. Kismet and M.T. Akay, 2010. Pathological and biochemical effects of therapeutic and supratherapeutic doses of celecoxib in Wistar albino male rats. Drug Chem. Toxicol., 33: 410-414.
CrossRefDirect Link - Korkina, L., M.G. Scordo, I. Deeva, E. Cesareo and C. de Luca, 2009. The chemical defensive system in the pathobiology of idiopathic environment-associated diseases. Curr. Drug Metab., 10: 914-931.
CrossRefDirect Link - Koul, A., L. Tanwar and N. Arora, 2009. Celecoxib administration exhibits tissue specific effect on 3H-benzo(a)pyrene-DNA adduct formation in cigarette smoke inhaling mice. Indian J. Exp. Biol., 47: 83-90.
Direct Link - Lefevre, G., M. Beljean-Leymarie, F. Beyerle, D. Bonnefont-Rousselot, J.P. Cristol, P. Therond and J. Torreilles, 1998. [Evaluation of lipid peroxidation by measuring thiobarbituric acid reactive substances]. Annales Biologie Clinique, 56: 305-319, (In French).
PubMedDirect Link - Lock, E.A. and C.J. Reed, 1998. Xenobiotic metabolizing enzymes of the kidney. Toxicol. Pathol., 26: 18-25.
CrossRefDirect Link - Long, L.K., J. Yang, Y. An and G. Liu, 2012. Disruption of a glutathione reductase encoding gene in Acremonium chrysogenum leads to reduction of its growth, cephalosporin production and antioxidative ability which is recovered by exogenous methionine. Fungal Genet. Biol., 49: 114-122.
CrossRefDirect Link - Mahmoudabad, A.B.Z., M. Saberi and J. Pirzad, 2008. Critical role of GSH in sulfur mustard-induced oxidative stress and cytotoxicity in human skin fibroblast cell line. Iran. J. Pharmaceut. Res., 7: 35-41.
Direct Link - Malhi, H., M.E. Guicciardi and G.J. Gores, 2010. Hepatocyte death: A clear and present danger. Physiol. Rev., 90: 1165-1194.
CrossRefDirect Link - Mandelker, L., 2008. Introduction to oxidative stress and mitochondrial dysfunction. Vet. Clin. North Am.: Small Anim. Pract., 38: 1-30.
CrossRefDirect Link - Murakami, A., D. Takahashi, T. Kinoshita, K. Koshimizu and H.W. Kim et al., 2002. Zerumbone, a Southeast Asian ginger sesquiterpene, markedly suppresses free radical generation, proinflammatory protein production and cancer cell proliferation accompanied by apoptosis: The α,β-unsaturated carbonyl group is a prerequisite. Carcinogenesis, 23: 795-802.
CrossRefPubMedDirect Link - Nagy, S.E.R., M.M. Nadia, M.M. Azza, Z.M. Naema and Z.R. Maha, 2011. Protection against oxidative damage induced by Schistosoma mansoni using susceptible/resistant nucleoproteins from Biomphalaria alexandrina Snails. Asian J. Biol. Sci., 4: 445-456.
CrossRefDirect Link - Nakamura, Y., Y. Chiho, A. Murakami, H. Ohigashi, T. Osawa and K. Uchida, 2004. Zerumbone, a tropical ginger sesquiterpene, activates phase II drug metabolizing enzymes. FEBS Lett., 572: 245-250.
CrossRefPubMedDirect Link - Ohkawa, H., N. Ohishi and K. Yagi, 1979. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem., 95: 351-358.
CrossRefPubMedDirect Link - Omura, T. and R. Sato, 1964. The carbon monoxide-binding pigment of liver microsomes. I. Evidence for its hemoprotein nature. J. Biol. Chem., 239: 2370-2378.
PubMedDirect Link - Pandey, A., R.K. Gupta and R. Srivastava, 2011. Curcumin-the yellow magic. Asian J. Applied Sci., 4: 343-354.
CrossRefDirect Link - Prakash, R.O., A. Rabinarayan and M.S. Kumar, 2011. Zingiber zerumbet (L.) Sm., a reservoir plant for therapeutic uses: A review. Int. J. Pharma World Res., 2: 1-23.
Direct Link - Prasong, S., 2011. Screening of antioxidant activity of some Samia ricini (Eri) silks: Comparison with Bombyx mori. J. Biol. Sci., 11: 336-339.
CrossRefDirect Link - Rahimi, H.R., M. Gholami, H.R. Khorram-Khorshid, F. Gharibdoost and M. Abdollahi, 2012. On the protective effects of IMOD and silymarin combination in a rat model of acute hepatic failure through anti oxidative stress mechanisms. Asian. J. Anim. Vet. Adv., 7: 38-45.
CrossRefDirect Link - Raza, H., A. John, M.S. Lakhani, I. Ahmed and W. Montague, 1998. Multiplicity and tissue specific expression of camel Cytochrome P450(s). Comp. Biochem. Physiol. Part C: Pharmacol. Toxicol. Endocrinol., 121: 205-211.
CrossRefDirect Link - Saboo, S.S., G.G. Tapadiya and S.S. Khadabadi, 2011. Antioxidant and hepatoprotective potential of isolated fraction of Revia hypocrateriformis. J. Biol. Sci., 11: 428-432.
CrossRefDirect Link - Sau, A., F.P. Tregno, F. Valentino, G. Federici and A.M. Caccuri, 2010. Glutathione transferases and development of new principles to overcome drug resistance. Arch. Biochem. Biophys., 500: 116-122.
CrossRefDirect Link - Sedlak, J. and R.H. Lindsay, 1968. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman's reagent. Anal. Biochem., 25: 192-205.
CrossRefPubMedDirect Link - Shuid, A.N., S. Mohamad, N. Mohamed, S.A. Mokhtar, N. Muhammad and I.N. Soelaiman, 2011. Bone oxidative changes during early fracture healing of postmenopausal osteoporosis rat model. Asian J. Anim. Vet. Adv., 6: 1193-1203.
CrossRefDirect Link - Sirisangtrakul, W. and B. Sripanidkulchai, 2011. Interference of thai traditional medicine (yahom ampanthong) on hepatic cytochrome p450 enzymes and pentobarbital-induced sleeping in mice. Pak. J. Biol. Sci., 14: 91-98.
CrossRefDirect Link - Sozer, S., G. Diniz and F. Lermioglu, 2011. Effects of celecoxib in young rats: Histopathological changes in tissues and alterations of oxidative stress/antioxidant defense system. Arch. Pharmacal Res., 34: 253-259.
CrossRefDirect Link - Sulaiman, M.R., E.K. Perimal, M.N. Akhtar, A.S. Mohamad and M.H. Khalid et al., 2010. Anti-inflammatory effect of zerumbone on acute and chronic inflammation models in mice. Fitoterapia, 81: 855-858.
CrossRefPubMedDirect Link - Ingelman-Sundberg, M., 2004. Human drug metabolising cytochrome P450 enzymes: Properties and polymorphisms. Naunyn-Schmiedeberg's Arch. Pharmacol., 369: 89-104.
CrossRefDirect Link - Tanaka, T., M. Shimizu, H. Kohno, S.I. Yoshitani and Y. Tsukio et al., 2001. Chemoprevention of azoxymethane-induced rat aberrant crypt foci by dietary zerumbone isolated from Zingiber zerumbet. Life Sci., 69: 1935-1945.
CrossRefPubMedDirect Link - Wangcharoenrung, L. and W. Warisnoicharoen, 2011. Change in mRNA expression of human cytochrome P450 by gold nanoparticles. J. Biol. Sci., 11: 173-180.
CrossRefDirect Link - Zhu, H.Z., X.L. Zhang and Y.S. Chen, 1998. Expression of glutathione S-transferase placental mRNA in hepatic preneoplastic lesions in rats. World J. Gastroenterol., 4: 38-40.
PubMedDirect Link - Zhao, S.Z., M.W. Reynolds, J. Lefkowith, A. Whelton and F.M. Arellano, 2001. A comparison of renal-related adverse drug reactions between rofecoxib and celecoxib, based on the world health organization/uppsala monitoring centre safety database. Clin. Therapeut., 23: 1478-1491.
CrossRefDirect Link - Gupta, S., P. Sarotra, R. Aggarwal, N. Dutta and N. Agnihotri, 2007. Role of oxidative stress in celecoxib-induced renal damage in wistar rats. Dig. Dis. Sci., 52: 3092-3098.
CrossRefDirect Link - Padam, B.S., H.S. Tin, F.Y. Chye and M.I. Abdullah, 2012. Antibacterial and antioxidative activities of the various solvent extracts of banana (Musa paradisiaca cv. Mysore) inflorescences. J. Biol. Sci., 12: 62-73.
CrossRefDirect Link