
S. Nahid
Department of Biochemistry, University of Shiraz, Shiraz, Iran
M. Haghkhah
Department of Pathobiology, School of Veterinary Medicine, University of Shiraz, Shiraz, Iran
M. Saeb
Department of Biochemistry, University of Shiraz, Shiraz, Iran
M. Aminlari
Department of Biochemistry, University of Shiraz, Shiraz, Iran
Asian Journal of Biotechnology
Year: 2011 | Volume: 3 | Issue: 3 | Page No.: 275-279







ABSTRACT
An efficient method to perform gene disruption plasmid by overlap extension PCR for site directed mutagenesis in Brucella abortus is demonstrated in this study. This method is based on three standard PCR and one fusion PCR reactions for construction of gene disruption construct and then insertion into a vector for performing of gene disruption plasmid. The present method in despite of conventional methods, is not restriction site dependent and require only one step host cell cloning.
PDF Abstract XML References Citation
Received: June 02, 2010;
Accepted: August 09, 2010;
Published: November 04, 2010
How to cite this article
S. Nahid, M. Haghkhah, M. Saeb and M. Aminlari, 2011. An Efficient Method for Gene Disruption in Brucella abortus by Overlap Extension PCR. Asian Journal of Biotechnology, 3: 275-279.
DOI: 10.3923/ajbkr.2011.275.279
URL: https://scialert.net/abstract/?doi=ajbkr.2011.275.279
DOI: 10.3923/ajbkr.2011.275.279
URL: https://scialert.net/abstract/?doi=ajbkr.2011.275.279
INTRODUCTION
Brucella abortus is an intracellular pathogen that causes abortion in bovines and can infect humans (Metcalf et al., 1994). A major area of interest is the determination of what genes contribute in virulence and persistence of wild or attenuated live vaccine strains. One of the most frequently employed tools for understanding gene function in bacteria is targeted gene disruption by replacement of the gene of interest with sequences coding for selectable markers that is facilitated by efficient homologous recombination (Wendland, 2003). Making of a gene disruption vector has been done by amplification of a fragment of target gene using PCR and insertion of PCR product into suitable vector, then insertion of a marker gene into a unique restriction site that resides approximately in the center of inserted gene fragment (Sambrook et al., 1989; Halling et al., 1991; Ugalde et al., 2003; Hornback and Roop, 2006). This method can be inflexible or time consuming because it requires using the existing restriction sites, or engineering additional unique site for inserting selectable marker.
In yeast, PCR-based construction of a gene disruption construct has been established (Ho et al., 1989; Horton et al., 1989). This widely used method involves the synthesis of a marker cassette flanked by short homology regions as linear DNA (Dillon and Rosen, 1990). In contrast to yeast, most bacteria are not readily transformable with linear DNA because of the presence of intracellular exonucleases that degrade linear DNA and hence the gene disruption construct should be inserted into a suitable plasmid vector to make a gene disruption vector.
Here we describe a procedure based on overlap extension PCR strategy and its application to disrupt Isocitrate lyase encoding gene (aceA) in Brucella abortus. This technique can be used in any system where homologous recombination requires longer regions than those that can be incorporated into synthetic oligonucleotides and will effectively eliminate the needing of restriction sites.
MATERIALS AND METHODS
Primary PCR reactions: Primers (1 and 2) was designed to amplify the kanamycin resistance cassette (KanR) from pET-28 vector. In parallel PCR reactions, the upstream (primers 3 and 4) and downstream (primers 5 and 6) segments of aceA gene was also amplified from Brucella abortus genomic DNA (sequence of primers has been shown in Table 1). Amplifications were performed using 0.1 ng plasmid and 10 ng genomic DNA. Each PCR reaction was performed in 50 μL that contained 20 mM Tris-HCl (pH 8.8), 10 mM (NH4)2SO4, 10 mM KCl, 1% Triton X-100, 1 mg mL-1 BSA, 500 nM of each primer, 0.2 mM of each dNTPs, 3.5 mM MgSO4 and 1 unit of Pfu DNA polymerase. The reactions were first heated to 95°C for 5 min, then 34 cycles of: 95°C for 1 min, 55°C for 1 min, 72°C for 2 min. Extensions were continued at 72°C for further 10 min.
Each PCR product was then size-fractionated on a 1% agarose gel in TAE buffer. The corresponding band was excised and then purified with DNA extraction kit (Fermentas) following the manufacturer protocol.
Fusion PCR reaction: As shown in Fig. 1, KanR amplification primers (1 and 2) have 5'-tail complementary to primers 4 and 5, respectively. Thus amplified segments in primary PCR reactions act as primer in fusion PCR. In the fusion PCR, the three purified PCR products generated in the primary PCR were added in equimolar (1-ng) amounts and primers 3 and 6 were used only. PCR reaction conditions were the same as primary PCR except that extension time changed to 3 min.
Construction of disruption plasmid. Fusion PCR product (disruption construct) was inserted into pUC18 through sacI cut site that carried by flanking primers 3 and 6 (Fig. 1). The resulting plasmid was cloned in E. coli HB101 (kanamycin was used as selectable marker) and harvested by alkaline lysis method.
Gene disruption: The gene disruption plasmid was introduced into Brucella abortus strain 119-3 by heat shock method (Chung et al., 1989) and transformants were selected based on its resistance to kanamycin and sensitivity to ampicillin. The aceA::kan mutation was confirmed by nested PCR using primers 3 and 6 as flanking and 1 and 2 as internal primers.
| Table 1: | Sequence of primers. The segments that shown as italic underlined letters in primers 1 and 2 complementary to primers 4 and 5, respectively. The segments that shown as italic underlined letters in primers 3 and 6 are sacI cut site |
 | |
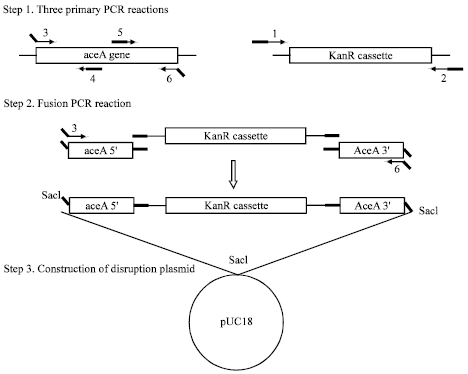 | |
| Fig. 1: | Schematic illustration of overlap extension PCR and construction of gene disruption plasmid. Bolded region in primer 1 overlaps with primer 4 and bolded region in primer 2 overlaps with primer 5 in fusion PCR. Bolded and oblique region in primers 3 and 6 carry sacI cut site |
RESULTS AND DISCUSSION
The strategy for constructing disruption plasmid is shown in Fig. 1. The procedure requires a total of fore PCR reaction: three standard PCR reactions and one fusion PCR reaction. The three standard PCR reactions are performed in step 1. These include one reaction to amplify the marker of choice and two reactions to amplify the regions flanking the site of marker insertion in the disruption target. In step 2, the fusion PCR reactions fuse the marker gene to upstream and downstream flanking segments to produce gene disruption construct. In three primary PCR reactions and fusion PCR reaction, the size of products observed on 1% agarose gel electrophoresis was in good agreement with the predicted size (Fig. 2).
Targeted disruption of the aceA gene and construction of ΔaceA strain was confirmed by nested PCR (Fig. 2). Susceptibility of mutant strain to ampicillin shows that gene disruption plasmid is unable to replicate in Brucella abortus.
Our method for disrupting Brucella abortus chromosomal gene is a modified use of the PCR overlap technique originally described by Ho et al. (1989) and Horton et al. (1989) and then developed by other researchers (Amberg et al., 1995; Kuwayama et al., 2002; Davidson et al., 2002) for gene disruption in yeast. The most common technique used for gene disruption in brucella is based on amplification and insertion of target gene into a suitable vector (vector should not be able to replicate in Brucella sp.) and after cloning in E. coli, marker cassette should be inserted via a unique cut site in approximately middle of inserted segment (Halling et al., 1991). In this report we have introduced a simple PCR method for the generation of a gene disruption construct and subsequently, gene disruption plasmid. This method requires only one standard PCR and one fusion PCR and is not depend on cut site existence and host cell cloning.
 | |
| Fig. 2: | Purified PCR Products on 1% agarose gel (TBE buffer) stained by BioAtlas. Lane 2, 3: upper and lower segments of aceA; Lane 4: KanR; Lane 5: fusion PCR product (gene disruption construct); Lane 6: wild type aceA PCR product by flanking primers; Lane 7: mutant aceA PCR product by flanking primers; Product of nested PCR in ÄaceA strain. Lane 1,9: DNA ladder |
This method allows researchers to design the exact configuration and sequence of the homologous regions flanking a selectable marker cassette by choosing the primer binding site at the target locus.
There are several important technical points that should be taken into consideration. First, all primers should be designed to have almost the same Tm value, which is crucial for annealing and amplification of the final PCR product in fusion PCR. Secondly, a high fidelity DNA polymerase should be used to ensure that the selectable marker gene product will function properly in vivo. Thirdly, purification of the three PCR products should be down without any contamination of the primers, as contaminated, tagged primers inhibit the precision of the fusion process.
In summary, this report introduces the PCR-based method for construction of gene disruption plasmid in Brucella abortus. This technique effectively eliminates the needing of restriction sites and multiple host cell cloning.
REFERENCES
- Amberg, D.C., D. Botstein and E.M. Beasley, 1995. Precise gene disruption in Saccharomyces cerevisiae by double fusion polymerase chain reaction. Yeast, 11: 1275-1280.
PubMedDirect Link - Chung, C.T., S.L. Niemela and R.H. Miller, 1989. One-step preparation of competent Escherichia coli: Transformation and storage of bacterial cells in the same solution. PNAS USA, 86: 2172-2175.
Direct Link - Davidson, R.C., J.R. Blankenship, P.R. Kraus, M.J. Berrios and M.C. Hull et al., 2002. A PCR-based strategy to generate integrative targeting alleles with large region of homology. Microbiology, 148: 2607-2615.
Direct Link - Dillon, P.J. and C.A. Rosen, 1990. A rapid method for the construction of synthetic genes using the polymerase chain reaction. BioTechniques, 9: 298-300.
PubMed - Halling, S.M., P.G. Detilleux, F.M. Tatum, B.A. Judge and J.E. Mayfield, 1991. Deletion of the BCSP31 gene of Brucella abortus by replacement. Infection Immunity, 59: 3863-3868.
PubMed - Ho, S.N., H.D. Hunt, R.M. Horton, J.K. Pullen and L.R. Pease, 1989. Site-directed mutagenesis by overlap extension using the polymerase chain reaction. Gene, 77: 51-59.
PubMed - Hornback, M.L. and R.M. Roop, 2006. The Brucella abortus xthA-1 gene product participates in base excision repair and resistance to oxidative killing but is not required for wild-type virulence in the mouse model. J. Bacteriol., 188: 1295-1300.
CrossRef - Kuwayama, H., S. Obara, T. Mario, M. Katoh, H. Urushihara and Y. Tanaka, 2002. PCR-mediated generation of a gene disruption construct without the use of DNA ligase and plasmid vectors. Nucleic Acids Res., 30: E2-E2.
PubMedDirect Link - Ugalde, J.E., D.J. Comerci, M.S. Leguizamon and R.A. Ugalde, 2003. Evaluation of Brucella abortus Phosphoglucomutase (pgm) mutant as a new live rough-phenotype vaccine. Infection Immunity, 71: 6264-6269.
Direct Link - Wendland, J., 2003. PCR-based methods facilitate targeted gene manipulations and cloning procedures. Curr. Genet., 44: 115-123.
PubMed