
B. Shaptadvipa
Department of Plant Breeding and Genetics, Assam Agricultural University, Jorhat-785013, India
R.N. Sarma
Department of Plant Breeding and Genetics, Assam Agricultural University, Jorhat-785013, India
Asian Journal of Biochemistry
Year: 2009 | Volume: 4 | Issue: 2 | Page No.: 45-54







ABSTRACT
Amylose content was determined in 41 traditional glutinous rice varieties of Assam classed as Birain, Bora and Chokuwa group during 2004-06. Average apparent amylose content in 6 accessions of Chokuwa (9.368%) was higher than 20 accessions of Bora group (0.502%) and 15 accessions of Birain (0.191%) genotypes. Mahsuri, a non-glutinous rice variety contained intermediate amylose content (21.2%). Eight SSR markers were used to assess genetic variability. The size of amplified fragments ranged from 100 to 500 bp. Among all genotypes, average Polymorphism Information Content (PIC) was 0.923. The average genetic similarity within the Birain accessions ranged from 0.119 to 0.571. Within Bora-Chokuwa accessions, similarity value ranged from 0.047 to 0.667. The average similarity was 0.228, which reflected that the Bora group could be more diverse than the Birain group. Amylose content is said to be highly influenced by environmental conditions. Since, Birain accessions were from the same Barak valley agro-climatic condition and Bora as well as Chokuwa were from the Brahmaputra valley, an analysis was made with corresponding pair-wise relative rate of increase (%) in apparent amylose contents as well as corresponding values of pair-wise Jaccard’s co-efficient of similarity among the accessions of Birain, Bora and Chokuwa groups of glutinous rice. It showed the existence of a matching relation between the increased values of respective apparent amylose content and the genetic similarity. It seems that apparent amylose content though cannot play a solid indicator for genetic variability in glutinous rice germplasm. However it may help to gauge biochemical bases towards genetic variability under same environmental condition.
PDF Abstract XML References Citation
How to cite this article
B. Shaptadvipa and R.N. Sarma, 2009. Study on Apparent Amylose Content in Context of Polymorphism Information Content along with Indices of Genetic Relationship Derived through SSR Markers in Birain, Bora and Chokuwa Groups of Traditional Glutinous Rice (Oryza sativa L.) of Assam. Asian Journal of Biochemistry, 4: 45-54.
DOI: 10.3923/ajb.2009.45.54
URL: https://scialert.net/abstract/?doi=ajb.2009.45.54
DOI: 10.3923/ajb.2009.45.54
URL: https://scialert.net/abstract/?doi=ajb.2009.45.54
INTRODUCTION
Glutinous rice is also called sticky rice, sweet rice, Waxy rice, Botan rice, Mochi rice and pearl rice. It is a type of short-grained rice that is especially sticky when cooked and is grown in Japan, Korea, China, Philippines, Thailand, Indonesia, India and Vietnam. This class of rice was introduced to Assam from Thailand or Burma a considerable time ago (Sharma et al., 1971). The Waxy rice of Assam has been classified in two groups, Bora (glutinous) and Chokuwa (semi-glutinous) based on apparent amylose content. Birain or Beruin rice is also another class of glutinous rice popular in Barak valley in the southern region of Assam, India and adjoining areas of Bangladesh. It is called glutinous in the sense of being glue-like or sticky and not in the sense of containing gluten. Its sticky properties should not be confused with the other varieties of rice that become sticky to one degree or another when cooked. Waxy rice of Assam has significance in social and religious ceremonies and forms a popular daily breakfast diet in rural Assam. Milled rice is also used in the preparation of snacks, flat rice, puffed rice, bamboo rice, sweet rice beer and other dishes. The parboiled polished Chokuwa rice swells on soaking and becomes soft and is used by the people of Assam as a fast food with curd. These multiplicities of uses make the glutinous rice very popular among farmers, in spite of the advent of modern high yielding rice varieties. What distinguishes glutinous rice from other types of rice is having no (or negligible amounts of) amylose content. Amylose, a linear polymer of glucose is one of two components of starch, the other being amylopectin. The apparent amylose content of Chokuwa varieties varies from 15 to 20%, while that of Bora varieties was found in trace quantities (Dutta and Barua, 1978).
To date more than 2000 SSR markers of cultivated rice are available, which provides a powerful tool for studying Oryza genus as a whole, as SSR markers have good cross-species amplification (McCouch et al., 2002). Waxy locus has been evaluated with SSR markers. Ayres et al. (1997) determined the relation between the Wx microsatellite polymorphism and the amylose content in the 92 US rice strains. They demonstrated that the Wx microsatellite is polymorphic enough to distinguish most rice strains in different amylose classes and the inheritance of the CT repeats can be traced through a cross-section of the US rice pedigree. Bergman et al. (2001) demonstrated that the CT repeats classes of Wx gene with n = 10 or 11, 14, 20 and 17 and 18 were categorized as high amylose, intermediate amylose and low-amylose types, respectively. Bao (2002) reported the usefulness of (CT)n microsatellitic markers of Wx gene in rice starch quality improvement. Prathepha (2003) characterized 68 rice strains belonging to two species of Oryza by using Wx microsatellite, (CT)n repeats, that are closely linked to the rice waxy gene. Prathepha (2003) observed the predominance of (CT)17 repeats in the Wx locus, in addition to identification of a unique microsatellite class,(CT)18 repeats, in the strains tested. While evaluating amylose content or identifying Waxy locus, no study has been reported to link amylose content with indices of genetic relationship among the glutinous germplasm for appropriate characterization. The present investigation was carried out to study variation for apparent amylose content in context of polymorphism information content along with indices of genetic relationship derived through SSR markers in Birain, Bora and Chokuwa groups of traditional glutinous rice of Assam.
MATERIALS AND METHODS
Seeds of 41 glutinous rice varieties (Table 1) were used in the present study during 2004-06. Among them, 15 (suffixing the word Birain) were collected from Regional Agricultural Research Station (RARS), Assam Agricultural University (AAU), Karimganj and the remaining 26 (suffixing the words Bora and Chakuwa) were from RARS, AAU, Titabor, representing distinct agro-climatic variation within the plains of the two river valleys, the former one from the Barak valley and rest two from the Brahmaputra valley of the northeast India. The traditional names as given by the farmers were retained in the germplasm accessions for field level identification. Karnel of Pusha Birain, Das Birain, Mow Birain, Kala Birain, Agirjal Birain, Akib Birain, Gela Bora, Memon Bora, Chokuwa Bora-2 and Parochokuwa Sali were red while rests were white. The accessions like Uba Birain, Gela Bora, Jaisungam Birain, Das Birain, Maju Chokuwa and Pakhiloga Bora could be classed as slender grain and rest were as either non-slender or bold.
Amylose content was determined by the method described by Sawbhagya and Bhattacharya (1979). Defatted moisture-free powdered (100 mg) sample was taken in a stoppered conical flask. One milliliter of distilled alcohol was added to wet the powder. Ten milliliter of 1 N NaOH was then gently added by the side and kept for overnight and heated next day in a boiling water bath for 2-3 min. After cooling, the volume was made up to 100 mL with distilled water. Five milliliter of above dispersion was transferred into a 100 mL volumetric flask. Fifty milliliter of water was added to it, followed by the addition of 1 mL of acetic acid and 2 mL of iodine (0.2%) solution.
| Table 1: | Apparent amylose content in (AACP) and corresponding pair-wise relative rate of increase (%) in AACP as well as corresponding values of pair-wise Jaccard’s co-efficient of similarity within Birain, Bora and Chokuwa groups of glutinous rice accessions of Assam |
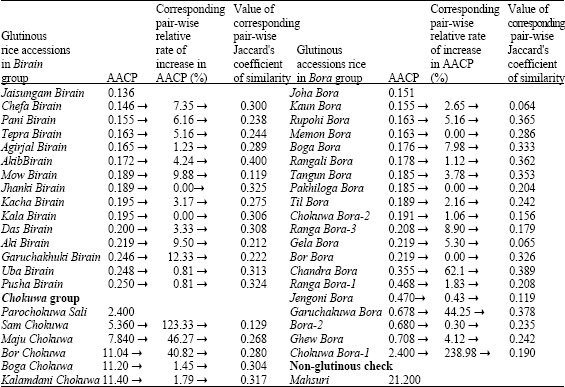 | |
| ! Value corresponding to glutinous rice accession in the previous row | |
The volume was made up to the mark with distilled water; pH of this solution was maintained at 4.5. One milliliter of standard amylose solution was taken and treated using the same procedure. A reagent blank was also prepared as such except for adding amylose extract or the standard amylose solution. After 30 min of interval, the intensity of colour developed was measured in spectrophotometer (Systronics UV-VIS, Model-118) at 630 nm against the reagent blank. Amylose content was calculated using the following relationship:
Where:
| R | = | 630 nm reading for sample dispersion |
| A | = | 630 nm reading for standard amylose solution |
| a | = | Amount of the standard amylose taken |
| r | = | Amount of sample taken |
The estimation was done in triplicate and their mean was recorded as g of amylose per 100 g in moisture free sample.
For studying genetic variation through SSR techniques, the total genomic DNA from each of the genotypes was extracted following the protocol of Plaschke et al. (1995) with slight modification. Genetic relatedness among the genotypes was then computed by using the Jaccard’s coefficient of similarity using SIMQUAL module of NISYS-pc. The pair-wise genetic similarity index was calculated as per Jaccard’s coefficient of similarity (Jaccard, 1908) is given below:
where, F is similarity index; NAB1 is number of bands present (Scored 1) in both accessions A and B; NAB0 is number of bands present in all test entries but not present in accession A or B; NT is total number of bands scored in the study.
RESULTS AND DISCUSSION
Apparent Amylose Content
The Birain genotypes had the lowest amylose content (0.191%) followed by bora genotypes (0.502%) and Chokuwa genotypes (9.368%), respectively. Among the glutinous rice accessions, the maximum and minimum amylose containing genotypes were Kalamdani Chokuwa (11.400%) and Jaisungam Birain (0.136%), respectively. Among Birain genotypes, the maximum and minimum amylose containing genotypes were Pusha Birain (0.250%) and Jaisungam Birain (0.136%), respectively. Among Chokuwa genotypes, the maximum amylose containing genotype was Kalamdani Chokuwa (11.400%) and the minimum amylose containing genotype was Sam Chokuwa (5.360%). Considering Bora genotypes alone, the maximum and minimum amylose containing genotypes were Ghew Bora (0.708%) and Joha Bora (0.151%), respectively. Present investigation revealed that the amylose content of indigenous glutinous rice ranged from 0.136 to 11.4%, indicating some of them as sticky or Waxy rice. In general, Birain group (0.191%) has lower amylose content than Bora genotypes (0.502%). Based on amylose content, Juliano et al. (1981) classified rice as Waxy (0-2% amylose), very low (5-12%), low (12-20%), intermediate ((20-25%) or high (25-30%). Dutta and Baruah (1978) reported amylose content in Bora genotypes as trace amount and that in Chokuwa from 17.14 to 19%, which were more than that observed in the present study. But Kandali et al. (1995) reported 0.95 to 1.25% of amylose in Bora accessions of Assam, which are in close correspondence with that observed in the present investigation. There is a possibility of duplication in Assam rice collection by having the similarity in names. It has been observed that the accessions with similar names (two Chokuwa Bora accessions and two Ranga Bora accessions) had different level of amylose content (Table 1). Moreover, amylose content is highly influenced by environmental conditions (Juliano and Pascual, 1980). So, based on amylose content alone, the duplicate nature of the four accessions could not be ascertained, warranting the use of better system for analyzing genetic variability and duplicate identification. However, for Birain group of Barak valley, no published report on amylose content is available. Glutinous rice lacks the starch amylose, which constitutes up to 30% of the total starch in non-glutinous rice endosperm (Oka, 1988). The glutinous phenotype arises through the disrupted expression of the amylose biosynthesis gene, (Wx), which encodes a granule-bound starch synthase (Sano, 1984). Glutinous rice contains a G to T mutation at the 5´splice site of Wx intron 1 that leads to incomplete post-transcriptional processing of Wx pre-mRNA (Wang et al., 1995; Bligh et al., 1998; Cai et al., 1998; Hirano et al., 1998; Isshiki et al., 1998). Glutinous rice does not have detectable levels of spliced mRNA as a result of this mutation (Wang et al., 1995; Bligh et al., 1998). But some degree of amylose synthesis is restored in varieties that carry the mutation due to display of cryptic splice site activation (Cai et al., 1998; Olsen and Puragganan, 2002). This might be the reason for detecting very low level of amylose in the present study and other similar studies in glutinous rice of Assam. Dipti et al. (2003) reported that the amylose content of Beruin rice of Bangladesh ranged from 7.9 to 10%, which was much higher than that obtained in Birain rice of Assam (0.136 to 0.250%) from the present study. Such differences in amylose content might be due to differences in genotypes under investigation along with environmental variation. However, methods to estimate amylose content might influence the results, which cannot be confirmed from the present investigation. Mahsuri, a non-glutinous rice variety contained intermediate amylose content (21.2%) remains soft and fluffy on cooking. Vanaja and Babu (2006) reported that Mahsuri contains 23.64% amylose, which is slightly more than the present study.
Therefore, it seems that the apparent amylose content in Chokuwa (9.368%) was higher than Bora (0.502%) and Birain (0.191%) genotypes.
Polymorphism Information Content (PIC)
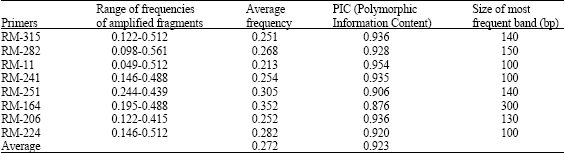
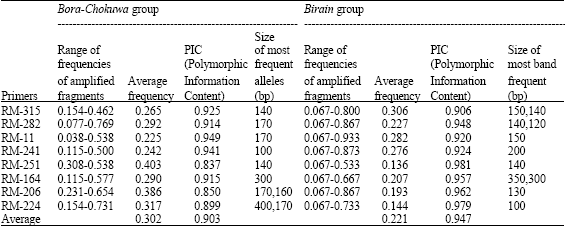
Eight SSR markers were used in the study to access genetic variability. The size of amplified fragments ranged from 100 to 500 bp. Average Polymorphism Information Content (PIC) was 0.923 in all the genotypes. The marker RM-11 showed a maximum PIC (0.954) and RM-164 showed a minimum PIC (0.876) across all the genotypes (Table 2). Within Bora-Chokuwa group, RM-11 generated maximum PIC (0.949) and RM-251 revealed a minimum PIC of 0.837. Within Birain accessions, RM-251 revealed a maximum PIC (0.981) and RM-315 showed a minimum PIC of 0.906 (Table 3). A lower PIC was observed for Bora-Chokuwa genotypes (0.903) than that for Birain genotypes (0.947).
Indices of Genetic Relationship
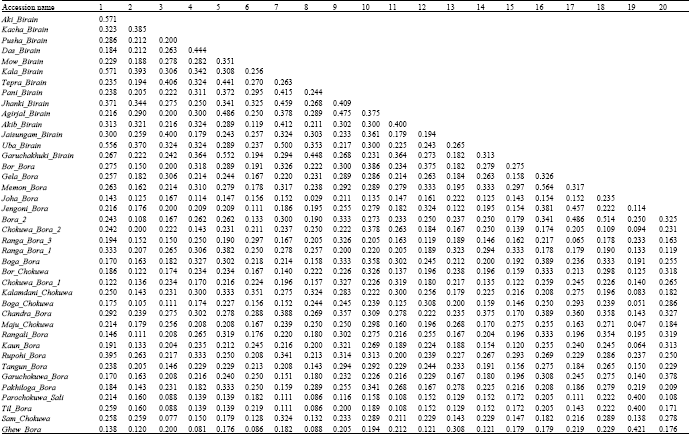
In all the genotypes, the genetic similarity ranged from 0.029 to 0.667 with an average similarity of 0.238. Maximum similarity (0.667) was observed between Parochokuwa Sali and Til Bora, while the minimum similarity (0.029) was exhibited by Tepra Birain and Joha Bora (Table 4).
Within the Birain accessions, similarity value ranged from 0.119 to 0.571 with an average of 0.306. The highest similarity (0.571) was exhibited by Chefa Birain with Aki Birain and Kala Birain, whereas, the lowest similarity value (0.119) was observed between Mow Birain and Akib Birain. Within Bora-Chokuwa accessions, similarity value ranged from 0.047 to 0.667. The accessions. Parochokuwa Sali and Til Bora exhibited maximum similarity (0.667) and Joha Bora and Maju Chokuwa showed minimum similarity (0.047). The average similarity was 0.228, which reflected that Bora group is more diverse than the Birain group.
| Table 2: | Level of polymorphism in few glutinous rice germplasm of Assam a s detected by SSR markers |
 | |
| Table 3: | Comparative analysis of level of polymorphism as detected by SSR markers in few glutinous rice germplasm of Assam |
 | |
| Table 4: | Pair-wise Jaccar’s coeffcient of smilarity based on SSR data in glutinous rise accessions under study |
 | |
| Table 4: | Continued |
-2k9-45-54.gif) | |
| 1: Chefa_Birain, 2: Aki_Birain, 3: Kacha_Birain, 4: Pusha_Birain, 5: Das_Birain, 6: Mow_Birain, 7: Kala_Birain, 8: Tepra_Birain, 9: Pani_Birain, 10: Jhanki_Birain, 11: Agirjal_Birain, 12: Akib_Birain, 13: Jaisungam_Birain1, 14: Uba_Birain, 15: Garuchakhuki_Birain, 16: Bor_Bora, 17: Gela_Bora, 18: Memon_Bora, 19: Joha_Bora, 20: Jengoni_Bora, 21: Bora_2, 22: Chokuwa_Bora_2, 23: Ranga_Bora_3, 24: Ranga_Bora_1, 25: Boga_Bora, 26: Bor_Chokuwa, 27: Chokuwa_Bora_1, 28: Kalamdani_Chokuwa, 29: Boga_Chokuwa, 30: Chandra_Bora, 31: Maju_Chokuwa, 32: Rangali_Bora, 33: Kaun_Bora, 34: Rupohi_Bora, 35: Tangun_Bora, 36: Garuchokuwa_Bora, 37: Pakhiloga_Bora, 38: Parochokuwa_Sali, 39: Til_Bora, 40: Sam_Chokuwa, Cophenetic correlation, r = 0.663 | |
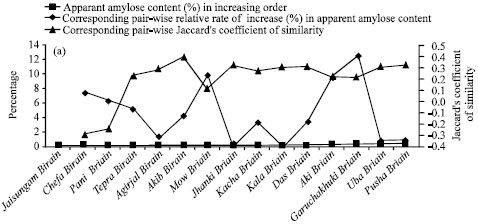 | |
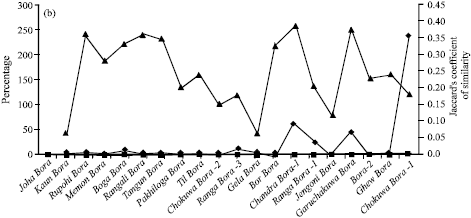
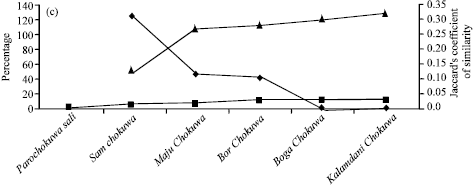
| Fig. 1: | Apparent amylose content (%) and corresponding pair-wise relative rate of increase (%) in apparent amylose contents as well as corresponding values of pair-wise Jaccard’s co-efficient of similarity in (a) Birain (b) Bora and © Chokuwa groups of glutinous rice accessions in Assam |
Apparent Amylose Content vis-à-vis Genetic Relationship
Amylose content is said to be highly influenced by environmental conditions. As respective accessions of Birain, Bora and Chokuwa groups were collected from the same environmental condition viz. the Barak sub-basin and the Brahmaputra valley agro-climatic conditions, an analysis was therefore made with corresponding pair-wise relative rate of increase (%) in AACP as well as corresponding values of pair-wise Jaccard’s co-efficient of similarity among the accessions of Birain, Bora and Chokuwa groups of glutinous rice (Table 1, Fig. 1a-c). It showed the existence of a matching relation between the increased values of respective AACP and the genetic similarity. For instance, in two Birain accessions viz., Akib Birain and Mow Birain, Das Birain and Garuchakuki Birain, the AACP was 0.172 and 0.189, 0.200 and 0.246 percent, respectively. Thus the AACP of Mow Birain, Garuchakuki Birain were about 10 and 23% higher than Akib Birain and Das Birain, which in turn, were about 4 and 2.6% higher in amylose content than that of Agirjal Birain (0.165) and Kala Birain (0.195).
The Jaccard’s coefficient (measures similarity) was decreased between Akib Birain and Mow Birain (0.119) by more than 200% than that of Akib Birain and Agirjal Birain (0.400) or between Aki Birain and Garuchakuki Birain (0.222) by more than 39% than that of Kala Birain and Das Birain (0.308). Similarly, AACP in Bora group of accessions, viz., Ghew Bora (0.708) was 4% higher than Bora-2 (0.680) against its nearest (0.678) Bora accession Goruchokuwa Bora. The Jaccard’s coefficient of similarity between Goruchokuwa Bora and Bora-2 was 0.378. The genetic similarity between Ghew Bora and Bora-2 was decreased by about 61 percent. Thus relative increase in AACP might be a reflection of direct or inverse proportionality to the genetic similarity in Birain and Bora accessions, which had lower amylose contents. The same reflection was not visualized in Chokuwa accessions which had comparatively higher amylose content. Here Maju Chokuwa (7.840%) was 46 and 227% higher amylose content than Sam Chokuwa (5.360%) and Parochokuwa (2.400%), respectively. Genetic similarity between Sam Chokuwa and Maju Chokuwa (0.268) was however increased by 108% over Sam Chokuwa and Parochokuwa (0.129). Thus it appeared that the AACP though could not play a concrete indicator for genetic variability in glutinous rice germplasm, however it might help to gauge genetic similarity, especially under lower amylose content. Customarily of course, apparent amylose content may help to characterize geographical indication of Waxy rice germplasm under the existing international patent regime.
ACKNOWLEDGMENTS
The authors are grateful to the Department of Plant Breeding and Genetics, Department of Biochemistry, Regional Agricultural Research Stations of Karimganj and Titabor under the Assam Agricultural University, Jorhat for rendering necessary help towards the study. The study was conducted on the funding of the NATP, Govemment of India.
REFERENCES
- Bligh, H.F., P.D. Larkin, P.S. Roach, C.A. Jones, H. Fu and W.D. Park, 1998. Use of alternate splice sites in granule-bound starch synthase mRNA from low-amylose rice varieties. Plant Mol. Biol., 38: 407-415.
CrossRefDirect Link - Cai, X.L., Z.Y. Wang, Y.Y. Xing, J.L. Zhang and M.M. Hong, 1998. Aberrant splicing of intron 1 leads to the heterogeneous 5' UTR and decreased expression of waxy gene in rice cultivars of intermediate amylose content. Plant J., 14: 459-465.
CrossRefDirect Link - Dipti, S.S., M.N. Bari and K.A. Kabir, 2003. Grain quality characteristics of some Beruin rice varieties of Bangladesh. Pak. J. Nutr., 2: 242-245.
CrossRefDirect Link - Hirano, H.Y., M. Eiguchi and Y. Sano, 1998. A single base change altered the regulation of the Waxy gene at the posttranscriptional level during the domestication of rice. Mol. Biol. Evol., 15: 978-987.
CrossRefDirect Link - Isshiki, M., K. Morino, M. Nakajima, R.J. Okagaki, S.R. Wessler, T. Izawa and K. Shimamoto, 1998. A naturally occurring functional allele of the rice waxy locus has a GT to TT mutation at the 5′splice site of the first intron. Plant J., 15: 133-138.
CrossRefDirect Link - Juliano, B.O., C.M. Perez, A.B. Blakeney, T. Castillo and N. Kongseree et al., 1981. International cooperative testing on the amylose content of milled rice. Starch/Staerke, 33: 157-162.
CrossRefDirect Link - Olsen, K.M. and M.D. Purugganan, 2002. Molecular evidence on the origin and evolution of glutinous rice. Genetics, 162: 941-950.
PubMedDirect Link - Sano, Y., 1984. Differential regulation of waxy gene expression in rice endosperm. Theor. Appl. Genet., 68: 467-473.
CrossRefDirect Link - Sawbhagya, C.M. and K.R. Bhattacharya, 1979. Simplified determination of amylose in milled rice. Starke, 31: 159-163.
CrossRefDirect Link - Sharma, S.D., J.M.R. Vellanki, K.I. Hakim and R.K. Singh, 1971. Primitive and current cultivars of rice in Assam: A rich source of valuable genes. Curr. Sci., 40: 126-128.
Direct Link - Vanaja, T. and L.C. Babu, 2006. Variation for grain and quality characteristics in rice (Oryza sativa L.). Indian J. Genet., 66: 13-15.
Direct Link - Wang, N.Y., F.Q. Zheng, G.Z. Shen, J.P. Gao and D.P. Snustad et al., 1995. The amylose content in rice endosperm is related to the post-transcriptional regulation of the waxy gene. Plant J., 7: 613-622.
CrossRefDirect Link - Ayres, N.M., A.M. McClung, P.D. Larkin, H.F.J. Bligh, C.A. Jones and W.D. Park, 1997. Microsatellites and a single-nucleotide polymorphism differentiate apparentamylose classes in an extended pedigree of US rice germ plasm. Theor. Applied Genet., 94: 773-781.
CrossRefDirect Link - Bergman, C.J., J.T. Delgado, A.M. McClung and R.G. Fjellstrom, 2001. An improved method for using a microsatellite in the rice waxy gene to determine amylose class. Cereal Chem., 78: 257-260.
CrossRef - Jaccard, P., 1908. Nouvelles recherches sur la distribution florale. Bull. Soc. Vaud. Sci. Nat., 44: 223-270.
CrossRefDirect Link - McCouch, S.R., L. Teytelman, Y. Xu, K.B. Lobos and K. Clare et al., 2002. Development and mapping of 2240 new SSR markers for rice (Oryza sativa L.). DNA Res., 9: 257-279.
CrossRefDirect Link - Plaschke, J., M.W. Ganal and M.S. Roder, 1995. Detection of genetic diversity in closely related bread wheat using microsatellite markers. Theor. Applied Genet., 91: 1001-1007.
CrossRefDirect Link
Dr. H. Singla Reply
The study contain very fine piece of information on indication of genetic relation between glutinous rice and the amylose contents- a good correlation between genomics and proteomics. The glutinous rice is an important constituent of all body toner, used in cosmetic industry. I congratulate the researchers for their innovative assessment of glutinous rices of India from genomics and proteomics points of view.