
P.D. Lokhande
Department of Organic Chemistry, The Center for Advanced Studies, University of Pune, Maharashtra, Pune, 411007, India
B.S. Kuchekar
Departrnent of Pharmaceutical Chemistry,
Government College of Pharmacy, Maharshtra, Karad 415124, India
A.R. Chabukswar
Department of Organic Chemistry, The Center for Advanced Studies, University of Pune, Maharashtra, Pune, 411007, India
S.C. Jagdale
Department of Organic Chemistry, The Center for Advanced Studies, University of Pune, Maharashtra, Pune, 411007, India
Asian Journal of Biochemistry
Year: 2006 | Volume: 1 | Issue: 1 | Page No.: 1-17







ABSTRACT
Nitric oxide is synthesized by the enzyme nitric oxide synthase. Nitric oxide synthase maintains the normal levels of nitric oxide in body. Discovery of nitric oxide synthase has lead to a new era in the drug research. Nitric oxide regulates the normal physiological balance and functions of the body. Three isoforms of nitric oxide synthase have been identified, an endothelial type, a neuronal type and a macrophage (inducible) type. Nitric oxide helps in regularizing normal blood pressure, relaxation of smooth muscle, release of growth hormone, insulin and pancreatic polypeptide. Nitric oxide plays very important role in host defense mechanism of body. Studies have shown that excess or deficiency of nitric oxideis associated with many diseases. In spite of the involvement of NO in several physiological functions, overproduction of NO can be harmful. An excess of NO produced by nNOS or iNOS is involved in pathological conditions, including post-ischemic stroke damage, septic shock, seizures, schizophrenia, migraine headaches, Alzheimer`s disease, development of colitis, tissue damage and inflammation, osteoporosis and rheumatoid arthritis, depression, neuroimmune diseases. In this study we have taken review of the important role of nitric oxide in biological system and the strategies to inhibit the overproduction of nitric oxide.
PDF Abstract XML References Citation
How to cite this article
P.D. Lokhande, B.S. Kuchekar, A.R. Chabukswar and S.C. Jagdale, 2006. Nitric Oxide: Role in Biological System. Asian Journal of Biochemistry, 1: 1-17.
DOI: 10.3923/ajb.2006.1.17
URL: https://scialert.net/abstract/?doi=ajb.2006.1.17
DOI: 10.3923/ajb.2006.1.17
URL: https://scialert.net/abstract/?doi=ajb.2006.1.17
Introduction
Nitric oxide is synthesised from the amino acid arginine (Fig. 1). Nitric oxide has played important role in the pathogenesis of diseases including hypertension, septic shock and dementia. (Moncada et al., 1991; Moncada and Higgs, 1993).
Nitric Oxide (NO), a molecular messenger synthesized by Nitric Oxide Synthase (NOS) from L-arginine and molecular oxygen, is involved in a number of physiological and pathological processes in mammalians (Ignarro et al., 1987). Three isoforms of nitric oxide synthase have been identified, an endothelial type, a neuronal type and a macrophage (inducible) type (Marsden et al., 1993). The genes for these enzymes have been localised to chromosome 7 (endothelial type), chromosome 12 (neuronal type) and chromosome 17 (macrophage type). The neuronal isoform is found in some central and peripheral neurons and the endothelial isoform is in vascular endothelium, platelets and the heart (endocardium and myocardium).
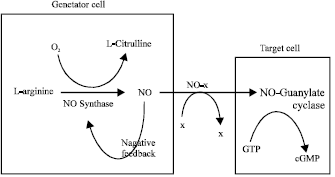 | |
| Fig. 1: | Synthesis of nitric oxide |
These two isoforms are normal constituents of the cells but the macrophage enzyme is not normally found in any healthy, quiescent cell type. Macrophage type nitric oxide synthase is expressed only after activation of cells with products of infection, including bacterial endotoxin or exotoxin, or certain inflammatory mediators, including the cytokines tumor necrosis factor or interleukin-1. It can be expressed in many types of cells including vascular smooth muscle, heart muscle, gut, immune cells and hepatocytes. This is an inducible enzyme and produces larger amounts of nitric oxide than the other two isoforms. Nitric oxide synthase regulate the production of nitric oxide in body. Excess or deficiency of the nitric oxide can lead to various diseases. Research have been focused on the strategies to control the production of nitric oxide in order to avoid various diseases, which are associated with nitric oxide balance. The discovery of nitric oxide synthase enzyme and the important role of nitric oxide in physiological system have lead to a new era in drug discovery programme.
Synthesis and Characteristics of Nitric Oxide
Nitric oxide is a free radical and is therefore highly reactive. It has a half-life of a few seconds and readily combines with other free radicals (Beckman and Crow, 1993). In biological systems it decomposes rapidly to yield nitrite and nitrate and this reaction is catalysed by transition metal like iron. Haemoglobin inactivates nitric oxide by binding it to form nitrosohaemoglobin and by catalysing the degradation of nitric oxide to nitrite and nitrate, resulting in the formation of methaemoglobin. Nitric oxide synthase has a haem moiety and negative feedback of nitric oxide production can occur (Fig. 1). Nitric oxide is most likely to act as a local messenger molecule transferring messages within and between individual cells.
Synthesis of Nitric Oxide
Nitric oxide synthase catalyses the synthesis of nitric oxide from L-arginine and molecular oxygen. L-citrulline is the byproduct. Nitric oxide itself might inhibit the activity of nitric oxide synthase by interacting with the haem moiety of this enzyme. Physiological effects are produced after nitric oxide binds to the haem moiety of guanylate cyclase and activates this enzyme to produce cyclic Guanosine Monophosphate (cGMP) from Guanosine Triphosphate (GTP) in target and generator cells. Carrier molecules (x) that stabilise nitric oxide have also been proposed.
Guanylate cyclase catalyses the formation of cyclic Guanosine Monophosphate (cGMP). Nitric oxide interacts with the haem moiety of this enzyme to activate it and the rise in cyclic guanosine monophosphate concentrations produces changes in cell function, often by affecting intracellular calcium concentrations. Nitric oxide synthesised by inducible nitric oxide synthase also activates guanylate cyclase, but the large quantities have additional toxic effects. High concentrations of nitric oxide inactivate enzymes containing transition metals, including certain mitochondrial enzymes. Inducible nitric oxide synthase seems to be part of the immune system (Hibbs, 1991).
Cardiovascular System
Nitric oxide is released continuously from arterial and arteriolar endothelium. Infusion of a nitric oxide synthase inhibitor (for example, N (super G)-monomethyl-L-arginine) into the brachial artery of healthy volunteers causes a substantial fall in resting forearm blood flow, indicating that continuous synthesis of nitric oxide is an important determinant of the basal tone of small arteries and arterioles. In contrast, veins do not seem to synthesis nitric oxide basally. Systemic injection of the inhibitor increases blood pressure in experimental animals and healthy volunteers but produces little change in venous pressure. Pulmonary vessels also synthesis nitric oxide continuously and this seems to be important for maintaining blood flow within the lungs and matching ventilation to perfusion. It is now clear that a rise in blood pressure or vasospasm in a vessel does not have to be due to an increase in vasoconstrictors; it could be due to a loss of the basal dilator tone mediated by nitric oxide (Calver et al., 1993).
Nitric oxide synthesised in the endothelium maintains the vasodilatation of the cardiovascular system and contributes to the thrombo resistance of vessels walls. It inhibits platelet aggregation and attenuates adhesion of platelets and white cells to the vessel wall. The antiaggregatory effects of nitric oxide are synergistic with those of another endothelium derived mediator, prostacyclin. Platelets also express nitric oxide synthase and the nitric oxide produced seems to prevent excessive platelet activation in response to aggregating stimuli. The endocardium is a continuation of endothelium and it is therefore unsurprising that it synthesises nitric oxide. Myocardial cells have also been shown to possess nitric oxide synthase. Nitric oxide affects cardiac relaxation during diastole and may be negatively inotropic (Henderson et al., 1992).
Stimulation of cardiac afferents either mechanically or chemically can induce reflex cardiovascular responses, including changes in Blood Pressure (BP), Heart rate (HR) and vasomotor tone (Tjen-A-Looi et al., 2001; Malliani, 1982; Reimann, 1980).
Myocardial ischemia is an important pathophysiological condition that causes both inhibitory and excitatory cardiac-cardiovascular reflexes. Stimulation of vagal afferents leads to hypotension, bradycardia and decreases in systemic vascular resistance (Oberg and Throen, 1973). Excitatory reflexes from the ventricular myocardium result from activation of cardiac sympathetic spinal afferents (Ignarro, 1989). Activation of sympathetic afferents increases BP, HR and myocardial contractility (Brown, 1967; Malliani et al., 1969). Metabolites such as prostaglandins, lactic acid, reactive oxygen species and Bradykinin (BK) released during myocardial ischemia can directly activate or sensitize cardiac afferent nerve endings (Kelm and Schrader 1990 and Pan et al., 1999). BK stimulates ventricular C-fiber sympathetic afferents during ischemia (Tjen ALooiS et al., 1998) leading to reflex activation of the cardiovascular system ( Rudbrook, 1990; Huang et al., 1995). In the periphery, endothelial NO acts as a potent vasodilator by stimulating guanylyl cyclase, which then generates cGMP, inducing smooth muscle relaxation (Just et al., 1994) and a change in myocardial contractility. Recent evidence also suggests that endogenous NO has BP-buffering capabilities comparable to that of the baroreceptors (Mohan et al., 1996; Goodson et al., 1994; Garthwaite, 1991).
There are other ways by which NO might modify cardiac-cardiovascular reflexes. For instance, it has been shown that NO acts as a neuromodulator and/or neurotransmitter in the central nervous system. A number of studies (Sakuma et al., 1992) have shown that inhibition of the synthesis of NO enhances vasoconstriction and release of norepinephrine during activation of the sympathetic nervous system. Also, there is growing evidence that a modulatory and facilitatory role for NO exists in medullary cardiovascular centers. However, it is still controversial whether NO inhibits or enhances sympathetic outflow, or causes both effects (Harada et al., 1993; Kagiyama et al., 1997). Endothelial NO plays an important role in the tonic and reflex regulation of arterial BP, whereas neuronal NO facilitates the sympatho excitatory cardiovascular reflex elicited by chemical stimulation of sympathetic cardiac afferents. Thus full expression of the cardiac-cardiovascular pressor reflex induced by epicardial application of BK is dependent on an intact central NO mechanism. Neuronal NOS facilitates, whereas endothelial NOS modulates, the excitatory cardiovascular reflex elicited by chemical stimulation of sympathetic cardiac afferents.
Peripheral and Central Nerves
Nerves staining for nitric oxide synthase have been found in the cardiovascular system, bronchial tree, urinary tract and gut and a "nitrergic" nervous system has been proposed. nitric oxide is released from many nerves previously classified as non-adrenergic, non-cholinergic nerves; however, it is probably only one of several transmitter substances released. Nitric oxide relaxes smooth muscle and nitrergic nerves are thought to play an important part in the dilatation of certain blood vessels (including the corpus cavernosum), adaptive relaxation of the stomach, relaxation of sphincters, the relaxant part of the peristaltic cycle, bronchodilatation and relaxation of smooth muscle in the upper and lower urinary tract.
Immunity and Inflammation
Nitric oxide synthesised by inducible nitric oxide synthase in activated murine macrophages is an important host defense mechanism. It kills pathogens including leishmania, Mycobacterium tuberculosis, malaria parasites and certain fungi; mediates "non-specific" immunity; and is toxic to tumor cells. Nitric oxide also regulates lymphocyte function and may have a role in inhibiting certain subsets of T helper cells (Liew et al.,1991). Nitric oxide also contributes to the inflammatory response. Vasodilatation may be mediated by inflammatory mediators stimulating endothelial nitric oxide synthase or by induction of macrophage isoform in endothelium, smooth muscle and inflammatory cells in the vessel wall. The nitric oxide produced might contribute to tissue leakage and damage (Vallance and Moncada, 1993).
Pregnancy
The vasodilatation and fall in blood pressure that occur in pregnancy may be partly due to nitric oxide and nitric oxide synthesised in the uterus may prevent uterine contraction (Kurtzman et al., 1993).
Endocrine Function
Arginine, the substrate for synthesis of nitric oxide, has been used by endocrinologists for many years to stimulate the release of growth hormone, insulin, pancreatic polypeptide and other hormones. It is unclear whether these effects are due to nitric oxide. Nitric oxide has also been proposed to have a role in regulating renin production and sodium homoeostasis in the kidney. Endothelial cell nitric oxide synthase is activated by increased shear stress across the cell surface and nitric oxide inhibits release of renin. This might provide a link between renal blood flow and the control of renin and sodium balance, with the endothelial cell acting as a signal transducer.
Diseases
Excess or deficiency of nitric oxide has been linked with many conditions. Nitric oxide role in physiological system have triggered a new area for drug discovery. Various diseases associated with nitric oxide can be avoided and treated with proper management of nitric oxide levels.
In the cardiovascular system loss of nitric oxide mediated effects has been found in every disease associated with increased vascular tone, vasospasm, or enhanced adhesion of platelets and white cells to the vessel wall. Abnormalities of the arginine- nitric oxide pathway have been shown in patients with hypertension, diabetes, hyperlipidaemia and overt atheroma (Barbul, 1986).
In the central nervous system, inhibition of nitric oxide synthesis impairs learning in rats, induces somnolence in sheep, enhances the action of certain anaesthetic agents, protects against epilepsy and reduces or increases damage caused by stroke depending on the experimental model. Vast overproduction of nitric oxide could contribute to programmed or pathological cell death in the central nervous system. Indeed, nitric oxide has been implicated in cerebral damage produced by stroke, Parkinson's disease and AIDS dementia (Snyder, 1993). Expression of inducible nitric oxide synthase in response to cytokines or endotoxin seems to be part of the inflammatory response and could contribute to vasodilatation, vascular leakage and tissue damage in some inflammatory conditions (Middleton et al., 1993; Boughton et al., 1993; De et al., 1993). Nitric oxide synthesis is enhanced in patients with septic shock and injection of an inhibitor reverses the hypotension, suggesting that overproduction of nitric oxide contributes to the pathophysiology (Petros et al., 1991). Nitric oxide has joined the list of mediators involved in local and systemic inflammation in humans. In patients with renal failure, abnormalities of the arginine-nitric oxide pathway might be due to accumulation of arginine analogues, which are known to accumulate as renal function deteriorates (Vallance et al., 1992).
Drugs based on nitric oxide have been used for over a century. It is now recognised that nitric oxide is the active moiety of glyceryl trinitrate and related nitro-vasodilators, which may be considered nitric oxide donors (Feelisch, 1991). Glyceryl trinitrate preferentially dilates veins and this accounts for a large part of its efficacy in angina and heart failure and for the unwanted effect of postural hypotension. The antiplatelet effects of nitric oxide released by these drugs may also have therapeutic importance. New nitrovasodilators are likely to emerge and there are already hints that it should be possible to deliver nitric oxide to specific tissues (De et al., 1994).
Recent studies showed the possibility of enhancing nitric oxide production by providing excess of the substrate, arginine. Although the mechanism by which this might work is far from clear (the amino acid is present in abundance and does not seem to be rate limiting for nitric oxide synthesis), there are reports that arginine prevents the onset of atheroma in experimental models and restores certain aspects of endothelial function in humans (Drexler et al., 1991). Arginine is present in relatively large amounts in nuts (particularly brazil nuts and almonds), shellfish and meats including beef, bacon and game.
Strategies for Regulation of Nitric Oxide Levels
In spite of the involvement of NO in several physiological functions, overproduction of NO can be harmful (Moore et al., 1993; Gross et al., 1992). An excess of NO produced by nNOS or iNOS is involved in pathological conditions, including post-ischemic stroke damage (Choi, 1988) septic shock (Crossin, 1991) seizures (Ferrendelli et al., 1980), schizophrenia (Das et al., 1995), migraine headaches (Thomsen et al., 1994), Alzheimer’s disease (Dorheim et al., 1994), tolerance to and dependence on morphine (Bhargava, 1995), development of colitis (Seo et al., 1995), tissue damage and inflammation (Kubes et al., 1991), overproduction of osteoclasts leading to osteoporosis, Paget’s disease and rheumatoid arthritis (MacIntyre et al., 1991), destruction of photoreceptors in the retina (Ross and Snyderm, 1990), long-term depression (Shibuki and Okada, 1991) and priaprism (Burnett et al., 1992). Increased formation of NO may also play an important role in neuroimmune diseases and in the pathogenesis of AIDS, dementia (Lipton et al., 1991). Because of the harmful effects of excess nitric oxide, inhibition of NO biosynthesis should be an important approach to the design of new drugs to treat these conditions. However, inhibition of its production also could be detrimental to the other essential functions of NO; it is very important that the inhibitory action does not affect the eNOS activity because of its important role in maintaining blood flow and pressure.
Potent inhibitors are more selective for neuronal NOS than endothelial NOS, could be useful in the treatment of cerebral ischemia (stroke) and other neurodegenerative diseases, whereas iNOS inhibition has been targeted for the treatment of septic shock (Wright et al., 1992) and arthritis (Hansen et al., 1998). NO overproduction can be attenuated by pharmacological agents that inhibit NOS activity through different mechanisms of action; in fact, the catalytic activity of NOS can be reduced by pharmacological attack on the three principal enzyme sites, the arginine, heme and BH4 sites.
Structure Activity Relationship (SAR) studies revealed that the distance between the heme cofactor and residues responsible for the recognition of L-amino acid, are quite similar (Lee et al., 1999a). In addition studies on the crystal structures of human eNOS and human iNOS catalytic domains revealed that active site residues of iNOS and eNOS are nearly identical and the size of the active site decreases in the order nNOS>iNOS>eNOS (Gerber et al., 1997). Because of the above-cited close similarity among the three NOS isoforms, the design of isoform-specific inhibitors is a challenging problem. However, despite an overall similarity, some subtle structural differences among the substrate binding sites of the NOS isoenzymes (Fan et al., 1997; Lowe et al., 1999) can be used to direct rationale design of second-generation isoform-specific NOS inhibitors with broad therapeutic potential.
Uptill now several NOS inhibitors acting with different mechanisms of action have been described (Marletta, 1994; Ogden and Morre 1995; Southan et al., 1996; Macdonald, 1996; Chabrier et al., 1999). Various degrees of potency and selectivity towards all the three isoforms together with others pharmacological actions have been reported (Mayer and Andrew 1998; Moore and Handy, 1997). Other compounds both synthetic and natural are listed in the recent literature (Honda et al., 1998; Quintela et al., 1999; Dirsch et al., 2000; Wang et al., 2000; Murakami et al., 2000; Matsuda et al., 2000) as inhibitors of the induction/expression of iNOS caused by Lipopolysaccharide (LPS) in macrophages and some of them show IC50 values in the nanomolar range.
Nitric oxide Inhibitor substances can be divided into two principal groups, aminoacid-based inhibitors and non-aminoacid-based inhibitors.
Aminoacid-Based Inhibitors
This class of derivatives can be further divided into three groups of compounds, L-arginine analogues, conformationally restricted arginine analogues and dipeptides.
L-Arginine Analogues
The well-known strategy to modify an enzyme’s substrate has been successful in the discovery of new NOS inhibitors. The first NOS inhibitors were mono- or di-substituted guanidino analogues of the L-arginine substrate.
Restricted Arginine Analogues
Second approach in the field of aminoacid-based inhibitors is the synthesis of compounds in which the free rotation of the methylene chain present in L-arginine analogues is hindered by the introduction of various rigid scaffolds. The underlying ideas in the design of these compounds are to increase selectivity and to better understand the preferred binding orientation of aminoacid-based inhibitors within the arginine binding site (Shearer et al., 1997; Lee et al., 1999b; Eustache et al.,1998). Conformational restriction appears to prevent the molecules from assuming the appropriate discriminatory binding orientations needed for high selectivity of the isoenzymes of NOS; thus, imposition of restricted conformations does not seem a good approach for the increase of inhibitor selectivity.
Dipeptides
Another group of aminoacid-based analogues include L-NNA-containing dipeptides. Because of the selectivity of L-NNA for nNOS vs iNOS, in order to improve selectivity in favor of nNOS, L-NNA (ArgNO2)- and phenylalanine (Phe)-containing dipeptides and dipeptide esters were developed (Silverman et al., 1997).To better investigate the SAR and to improve the potential bioavailability of the dipeptide inhibitors, a series of peptidomimetic analogues were evaluated (Huang et al., 2000). other dipeptides containing arginine-analogues have also designed by following dipeptide approach (Kobayashi et al., 1999).
Non-Aminoacid-Based Inhibitors
For the specific selectivity and the therapeutic profile of L-Arginine derivatives, non-aminoacid-based inhibitors of NOS have been studied. For example; guanidines (Misko et al., 1993), isothioureas (Garvey et al., 1994; Southan et al., 1995) amidines (Moore et al., 1996a,b) thiazines (Nakane et al., 1995), imidazoles (Wolff et al., 1993a; Chabin et al., 1996) indazoles (Bland-Ward et al., 1995), benzoxazoles (Shankaran et al., 1997), pyridines and pteridines (Fröhlich et al., 1999).
Amidinic Compounds
These include all those compounds containing a carbamidine carbon bound to either N (guanidines), S (isothioureas) or another carbon (simple amidines).
Guanidines
Guanidines are only weak inhibitors (Tilton et al., 1993), guanidine moiety can mimic L-Arg for recognition at the binding site so that it can be considered a potential pharmacophore for NOS inhibition. Aminoguanidine is the most representative guanidine derivative described as a selective inhibitor of mouse inducible NOS, acting as a mechanism-based inactivator (Wolff and Lubeskie, 1995). Numerous in vivo studies have demonstrated that this inhibition could be useful in the treatment of disease states characterized by the pathological overproduction of NO from iNOS such as adjuvant-induced arthritis (Connor et al., 1995), autoimmune-encephalomyelitis (Cross et al., 1994) or diabetes (Corbett et al., 1992).
Isothioureas
Isothiourea containing compounds emerged early as very potent and selective inhibitors of human inducible isoform vs endothelial NOS. S-Ethylisothiourea S, S’- (1,3-phenylenebis (1,2-ethanediyl)
bisisothiourea and 2-amino-5, 6-dihydro-6-methyl-4H-1, 3-thiazine were the most representative compounds for potency and selectivity (McMillan and Masters, 1993). The SAR studies of these compounds have revealed that the active sites and surrounding regions of the human isoenzymes could be different and are useful to propose a model of binding of NOS substrates and inhibitors, clinical use of these isothiourea-based compounds is limited by poor cellular penetration and by their systemic toxicity (Tracey et al., 1995). An improvement in cellular penetration was achieved by some aminoalkylisothioureas among which aminoethylisothiourea (Southan and Szabo 1996) and aminoethylisoselenourea (Southan et al., 1996b) are reported as more potent inhibitors of nitrite formation in immunostimulated J774 macrophages (a model for iNOS activity) than L-NMA, with IC50 in the low micromolar range.
Amidines
From a comparison of potency and selectivity between aminoacid-based and non-aminoacid-based inhibitors of the three human isoforms it emerged that aminoacid moiety is not essential for inhibition. Therefore, simple amidine derivatives, which represent a simplification of guanidino moiety of aminoacid analogue, were tested; 2-methylthioacetamidine and 2-thienylcarbamidine were the most potent compounds of the series. Although none of these compounds showed appreciable selectivity, these studies confirmed that the amidine group by itself could be considered a common pharmacophore for NOS inhibition and opened the way for the development of potent and selective NOS inhibitors.
Substitutions on the five membered ring, 2-iminopyrrolidine were investigated (Hagen et al., 1998) and gave the most potent and selective cyclic amidines known so far with compounds active against all forms of NOS in the submicromolar range. N- (3- (Aminomethyl) benzyl) acetamidine, derived from a progressive simplification of bisisothiourea through its corresponding bisamidine (Garvey et al., 1997), was identified as a selective inhibitor of iNOS vs eNOS and nNOS. Among numerous amidines and guanidines synthesized as NOS inhibitors, N-phenyl-2-thiophenecar-boxamidine, named, was a compound, which showed submicromolar potency against nNOS and low selectivity vs eNOS (Reif et al., 2000).
Heterocyclic Compounds
The second group of non-aminoacid-based NOS inhibitors includes a range of heterocyclic compounds among which the most represented are, indazoles, imidazoles and analogues of tetrahydrobiopterine.
Indazoles
Inspite of the absence of in vitro selectivity in enzymatic or functional assays (Moore et al., 1993; Babbedge et al., 1993), 7-nitroindazole, named 7-NI, may be considered as the first selective inhibitor of nNOS in vivo; in fact, several studies have demonstrated its protective effects in pain, experimental stroke and in a mouse model of Parkinson’s disease with minimal systemic pressure effects (Schulz et al., 1995). This discrepancy is probably due to a better uptake into neurons vs endothelial cells, or to an extensive catabolism of by xanthine oxidase in the vascular tissue rather than in the brain. The inhibition mechanism of has been extensively studied and is referred to as multiple because competition with both L-Arg and BH4 has been observed (Wolff et al., 1994; Mayer et al., 1994). Based on the available SAR on indazole compounds, essentially it can be said that inhibition of nNOS was a characteristic property of the indazole nucleus and nitration at the 5-, 6- or 7-position resulted in a graded increase in inhibitory potency, whereas the presence of an amino-group at the 5-or 6-position, gave totally inactive compounds. The effect of chlorination was not clear since 3-chloroindazole was more active than indazole, while 3-chloro-5-nitroindazole was less active than the corresponding 5-nitro derivative. 3-Bromo-7-nitroindazole were also reported as interesting inhibitors of this class.
Imidazoles
Since it has been demonstrated that NOS contains a very similar heme site to that of cytochrome-P450 (White and Marletta, 1992), imidazole, 1-phenyl, 2-phenyl and 4-phenylimidazole and antimycotic drugs (myconazole, ketoconazole and clotrimazole) were tested as inhibitors (Wolff et al., 1993). In fact, it is well known that imidazoles inhibit the activity of various heme-containing proteins by binding to the heme group (Rogerson et al., 1977). 1-phenylimidazole gave better selectivity for iNOS vs eNOS (Wolff et al., 1994). Substitution with a phenyl group at the 2-or 4-position generally gave a severe loss of inhibition against all isoforms tested.
SAR on a series of simple commercially available imidazoles as inhibitors of recombinant human iNOS, confirmed that the 2-and the 4-position of imidazole cannot be substituted whereas modification at the 1-position with alkyl or aromatic substituents were well tolerated. From pharmacological studies 1-(2-trifluoro-methyl-phenyl) imidazole named TRIM, emerged as the first imidazole derivative endowed with relatively potent inhibition against nNOS and iNOS. Imidazole analogues of L-Arg bearing various substituents at the imidazolic ring have shown NOS inhibitory activity (Ulhaq et al., 1998).
For improvement in potency, some related azoles were designed and tested. The unsubstituted derivative 2-amino-5- (imidazol-1-yl) pentanoic acid was identified as the most potent member against rat iNOS, rat nNOS and a human derived constitutive NOS. Imidazole-containing aminoacids and guanidine-substituted imidazoles (Atkinson et al., 1999), designed to bind simultaneously to the heme and to aminoacid binding sites, were prepared and tested as inhibitors of the three isoforms of NOS. In fact, X-ray crystallographic studies of the iNOS oxygenase domain/imidazole complex revealed two molecules of imidazole bound in the active site, one directly to the heme iron as a sixth ligand and the second hydrogen-bound to Glu, an amino acid residue responsible of the binding with the guanidino group of L-Arg.
In the case of imidazole-containing aminoacids, compounds with a chain with an odd number of methylene groups were found to be potent inhibitors of iNOS and nNOS with poor selectivity vs eNOS. In the case of guanidine substituted imidazoles, four types of compounds were designed and tested. In the previous years (McMillan et al., 2000), potent and selective inhibitors of inducible NOS were identified in an encoded combinatorial chemical library based on a pyrimidine-imidazole core, that was selected because it was structurally related to phenylimidazoles. Two series of N-substituted imidazoles showed better inhibition of rat recombinant nNOS than human platelet eNOS for example N-phenacyl-imidazoles (Salerno et al., 1999a) and N- (aryloxy) alkyl-imidazoles (Salerno et al., 1999b).
Analogues of Tetrahydrobiopterine
Due to the structural relation with the enzymatic cofactor tetrahydrobiopterine BH4, pteridine based compounds have been described as NOS inhibitors. This approach appears promising because of the much lower affinity and selectivity of BH4 binding to other pteridine dependent enzymes (Mayer and Werner, 1995). Among a series of tested compounds, the 4-amino analogue of BH4, 5, 6, 7, 8-tetrahydro-6- (D-threo-1,2-dihydroxypropyl)pterin, was a potent inhibitor of the recombinant rat brain NOS both in vitro and in vivo (Mayer and Hemmens, 1994; Pfeiffer et al., 1997; Schmidt et al., 1999).
Other pteridine-based compounds, named anti-pterines, structurally related to, were described as NOS inhibitors. Some of these compounds specifically interacted with the BH4 binding site of NOS without interference with any other known cofactors or substrate binding sites (Bömmel et al., 1998). Among heterocycles with NOS inhibitory activity, 1,2-diaminobenzimidazoles was the first compound. The parent compound was a selective inhibitor of human iNOS with a competitive action with L-Arg, whereas its 1-methylamino analogue resulted potent against nNOS and essentially inactive against the other isoforms. This selectivity was probably due to steric requirements of the active sites, as confirmed by the total inactivity of the 1-isopropylamino derivative; in addition, the role of the amino group in the 1-position was critical as confirmed by the total inactivity of the 1-ethyl derivative. This work demonstrated that it is possible to inhibit iNOS or nNOS by means of non-aminoacid-based compounds.
A series of benzoxazolones has been synthesized and tested as inhibitors of NOS, starting from a broad based screening from which the morpholino derivative, which showed some selectivity for human iNOS vs the other isoforms, was selected. Substitution at the side chain with other amines maintained the observed activity and selectivity only in pyrrolidine and N-methyl-piperazine derivatives, whereas it gave partial or total loss of activity for the other tested compounds, especially in characterized by a longer alkyl chain between the morpholino and benzoxazolone nitrogens, indicating that the distance between the basic and aromatic functionalities was critical for iNOS inhibition. Also the benzoxazolone ring, especially when substituted with a 5-CH3, was endowed with iNOS and nNOS activity and some selectivity vs eNOS, while pyridine bioisosters and the oxathiazolone were totally inactive (Boer et al., 2000).
A substituted 2-aminopyridine showed interesting rat and human nNOS inhibitory activity with some selectivity vs human eNOS. A simple pyridine derivative, 2-amino-4-methylpyridine, was previously reported as a potent inhibitor (competitive with respect to L-Arg) of the hiNOS. 2-amino-4-methylpyridine can be used as a radioligand for affinity and selectivity studies of a number of selected NOS inhibitors towards the three isoforms of NOS.
Conclusions
Nitric oxide is playing a very important role in maintaining the normal physiological balance of the human body. The discovery of nitric oxide synthase and its three isoforms open a new way for drug discovery strategies. Selective inhibition of nitric oxide synthase is helpful in fighting against various serious diseases. New chemical compounds are showing good results in the studies. L- Arginine has served as a best target for the inhibition of nitric oxide. Mono or Di substituted guanidine derivatives, Dipeptide series are the examples of aminoacid based inhibitors. Among the Non aminoacid based inhibitors Isothiourea, Amidines, Indazole, Imidazole, Tetrahydrobiopterine shows potent and selective inhibition. Further detailed structure activity relationship of all these compounds along with the stereochemical aspect will be useful for the development of more potent and selective molecules for nitric oxide inhibition. However very few data is available so far on phytochemical investigation of the compounds which can produce better results in specific and effective inhibition of Nitric oxide Synthase. Discovery of new synthetic and natural products for regulation of normal nitric oxide levels will be a milestone in the drug discovery programme in nearby future.
REFERENCES
- Atkinson, R.N. and S.B. King, 1999. Guanidine substituted imidazoles as inhibitors of nitric oxide. Bioorg. Med. Chem. Lett., 9: 2953-2958.
CrossRef - Babbedge, R.C., P.A.B. Ward, S.L. Hart and P.K. Moore, 1993. Inhibition of rat cerebellar nitric oxide synthase by 7 nitroindazole and related substituted indazoles. Br. J. Pharmacol., 110: 225-228.
Direct Link - Barbul, A., 1986. Arginine biochemistry physiology and therapeutic implications. J. Parenteral Enteral Nutr., 10: 227-238.
CrossRef - Beckman, J.S. and J.P. Crow, 1993. The chemistry of peroxynitrite a product from the reaction of nitric oxide and pathological implications of nitric oxide superoxide and peroxynitrite formation. Biochem. Soc. Trans., 21: 330-334.
PubMed - Bhargava, H.N., 1995. Attenuation of tolerance to and physical dependence on morphine in the rat by inhibition of nitric oxide synthase. Gen. Pharmacol. Vascular Syst., 26: 1049-1053.
CrossRefDirect Link - Ward, PA.B. and P.K. Moore, 1995. 7 Nitro indazole derivatives are potent inhibitors of brain endothelium and inducible isoforms of nitric oxide synthase. Life Sci., 57: 131-333.
CrossRef - Boer, R., U.W. Rudiger, T. Klein and B. Mirau, S. Haas and I. Baur, 2000. The inhibitory potency and selectivity of arginine substrate site nitric-oxide synthase inhibitors is solely determined by their affinity toward the different isoenzymes. Mol. Pharmacol., 58: 1026-1034.
CrossRefDirect Link - Bommel, H.M., A. Reif, L.G. Frohlich , A. Frey and H. Hofmann et al., 1998. Anti pterins as tools to characterize the function of tetrahydrobiopterin in no synthase. J. Biol. Chem., 273: 33142-33149.
PubMed - Boughton, S.N., S.M. Evans, C.J. Hawkey, A.T. Cole, M. Balsitis, B.J. Whittle and S. Moncada, 1993. Nitric oxidesynthase activity in ulcerative colitis and crohns disease. Lancet., 342: 338-340.
PubMed - Brown, A.M., 1967. Excitation of afferent cardiac sympathetic nerve fibres during myocardial ischaemia. J. Physiol., 190: 35-53.
Direct Link - Burnett, A.L., C.J. Lowenstein, D.S. Bredt, T.S. Chang and S.H. Synder, 1992. Nitric oxide a physiologic mediator of penile erection. Science, 257: 401-403.
CrossRef - Calver, A., J. Collier and P. Vallance, 1993. Nitric oxide and cardiovascular control. Exp. Physiol., 78: 303-326.
Direct Link - Chabin, R.M., E. McCauley, J.R. Calaycay, T.M. Kelly and K.L. MacNaul et al., 1996. Active site structure analysis of recombinant human inducible nitric oxide synthase using imidazole. Biochemistry, 35: 9567-9575.
CrossRef - Chabrier, P.E., C.D. Pallardy and M. Auguet, 1999. Nitric oxide synthases targets for therapeutic strategies in neurological diseases. Cell. Mol. Life. Sci., 55: 1029-1035.
Direct Link - Choi, D.W., 1988. Glutamate neurotoxicity and diseases of the nervous system. Neuron., 1: 623-634.
PubMed - Connor, J.R., P.T. Manning, S.L. Settle, W.M. Moore and G.M. Jerome et al., 1995. Suppression of adjuvant induced arthritis by selective inhibition of inducible nitric oxide synthase. Eur. J. Pharmacol., 273: 15-24.
PubMed - Corbett, A., R.G. Tilton, K. Chang, K.S. Hasan and M.A. Sweetland et al., 1992. Aminoguanidine a novel inhibitor of nitric oxide formation prevents diabetic vascular dysfunction. Diabities, 41: 552-556.
PubMed - Cross, A.H., T.P. Misko, R.F. Lin, W.F. Hickey, J.L. Trotter and R.G. Tilton, 1994. Aminoguanidine an inhibitor of inducible nitric oxide synthase ameliorates experimental autoimmune encephalomyelitis in SJL mice. J. Clin. Invest., 93: 2684-2690.
CrossRef - Crossin, K.L., 1991. Nitric oxide (NO) a versatile second messenger in brain. Trends. Biochem. Sci., 16: 81-82.
PubMed - Das, I., N.S. Khan, B.K. Puri, S.R. Sooranna, J. Debelleroche and S.R. Hirsch, 1995. Elevated platelet calcium mobilization and nitric oxide synthase activity may reflect abnormalities in schizophrenic brain. Biochem. Biophys. Res. Commun., 21: 375-380.
CrossRef - De, B.A.J., M.W. Radomski, H.J.F. Why, P.J. Richardson and C.A. Bucknall, 1993. Nitric oxide synthase activities in human myocardium. Lancet., 341: 84-85.
Direct Link - De Belder, A.J., R. Mac Allister, M.W. Radomski and S. Moncada, 1994. Effects of S-nitroso-glutathione in the human forearm circulation. Evidence for selective inhibition of platelet activation. Cardiovasc. Res., 28: 691-694.
CrossRefDirect Link - Dirsch, V.M., H. Stuppner, E.P. Ellmerer-Muller and A.M. Vollmar, 2000. Structural requirements of sesquiterpene lactones to inhibit LPS-induced nitric oxide synthesis in RAW 264.7 macrophages. Bioorg. Med. Chem., 8: 2747-2753.
CrossRefDirect Link - Dorheim, M.A., W.R. Tracey, J.S. Pollock and P. Grammas, 1994. Nitric oxide synthase activity is elevated in brain microvessels in alzheimers disease Biochem. Biophys. Res. Commun., 205: 659-665.
PubMed - Drexler, H., A.M. Zeiher, K. Meinzer and H. Just, 1991. Correction of endothelial dysfunction in coronary microcirculation of hypercholesterlaemic patients by L arginine. Lancet, 338: 1546-1550.
Direct Link - Eustache, J., A. Grob, C. Lam, O. Sellier and G. Schulz, 1998. Conformationally constrained NO synthase inhibitors rigid analogs of L N iminoethylornithine. Bioorg. Med. Chem. Lett., 8: 2961-2966.
PubMed - Fan, B., J. Wang, D.J. Stuehr and D.L. Rousseau, 1997. No synthase isozymes have distinct substrate binding sites. Biochemistry, 36: 12660-12665.
PubMed - Feelisch, M., 1991. Preparation and handling of aqueous no solutions. J. Cardiovasc. Pharmacol., 17: 25-33.
Direct Link - Ferrendelli, J.A., A.C. Blank and R.A. Gross, 1980. Relationships between seizure activity and cyclic nucleotide levels in brain. Brain. Res., 200: 93-103.
PubMed - Frohlich, L.G., P. Kotsonis, H. Traub and S.T. Moghadam, 1999. Inhibition of neuronal nitric oxide synthase by 4 amino pteridine derivatives structure activity relationship of antagonists of (6R) 5 6 7 8 tetrahydrobiopterin cofactor. J. Med. Chem., 42: 4108-4121.
PubMed - Garvey, E.P., J.A. Oplinger, G.J. Tanoury, P.A. Sherman and M. Fowler et al., 1994. Potent and selective inhibition of human nitric oxide synthases. Inhibition by non-amino acid isothioureas. J. Biol. Chem., 269: 26669-26676.
PubMed - Goodson, A.R., J.M. Leibold and D.D. Gutterman, 1994. Inhibition of nitric oxide synthesis augments centrally induced sympathetic coronary vasoconstriction in cats. Am. J. Physiol. Heart Circ. Physiol., 267: 1272-1278.
Direct Link - Gross, S.S. and R. Levi, 1992. Nitric oxide synthesis an absolute requirement for cytokine induced nitric oxide generation by smooth muscle. J. Biol. Chem., 267: 25722-25729.
Direct Link - Hagen, T.J., A.A. Bergmanis, S.W. Kramer, K.F. Fok and E.A. Schmelzer et al., 1998. Iminopyrrolidines as potent and selective inhibitors of human inducible nitric oxide synthase. J. Med. Chem., 41: 3675-3683.
CrossRef - Hansen, D.W., K.B. Peterson, M. Trivedi, S.W. Kramer and R.K. Webber et al. 1998. Iminohomopiperidinium salts as selective inhibitors of inducible nitric oxide synthase (iNOS). J. Med. Chem., 41: 1361-1366.
CrossRef - Harada, S., S. Tokunaga, M. Momohara, H. Masaki, T. Tagawa, T. Imaizumi and A. Takeshita, 1993. Inhibition of nitric oxide formation in the nucleus tractus solitarius increases renal sympathetic nerve activity in rabbits. Circ. Res., 72: 511-516.
Direct Link - Henderson, A.H., M.J. Lewis, A.M. Shah and J.A. Smith, 1992. Endothelium endocardium and cardiac contraction. Cardiovasc. Res., 26: 305-308.
CrossRef - Hibbs, J.B.J., 1991. Synthesis of nitric oxide from L arginine a recently discovered pathway induced by cytokines with antitumour and antimicrobial activity. Res. Immunol., 142: 565-569.
PubMed - Huang, H., P. Martasek, L.J. Roman and R.B. Silverman, 2000. Synthesis and evaluation of peptidomimetics as selective inhibitors and active site probes of nitric oxide synthases. J. Med. Chem., 43: 2938-2945.
CrossRef - Huang, H.S., H.L. Pan, G.L. Stahl and J.C. Longhurst, 1995. Ischemia and reperfusion sensitive cardiac sympathetic afferents influence of H2O2 and hydroxyl radicals. Am. J. Physiol. Heart Circ. Physiol., 269: 888-901.
Direct Link - Ignarro, L.J., G.M. Buga, G.M. Wood, R.E. Byrns and G. Chaudhuri, 1987. Endothelium derived relaxing factor produced and released from artery and vein is nitric oxide. Proc. Natl. Acad. Sci., USA., 84: 9265-9269.
PubMed - Ignarro, L.J., 1989. Biological actions and properties of endothelium derived nitric oxide formed and released from artery and vein. Circ. Res., 65: 1-21.
PubMedDirect Link - Just, A., U. Wittmann, B. Nafz, C.D. Wagner, H. Ehmke, H.R. Kirchheim and P.B. Persson, 1994. The blood pressure buffering capacity of nitric oxide by comparison to the baroreceptor reflex. Am. J. Physiol. Heart Circ. Physiol., 267: 521-527.
Direct Link - Kagiyama, S., T. Tsuchihashi, I. Abe and M. Fujishima, 1997. Cardiovascular effects of nitric oxide in the rostral ventrolateral medulla of rats. Brain. Res., 757: 155-158.
CrossRef - Kelm, M. and J. Schrader, 1990. Control of coronary vascular tone by nitric oxide. Circ. Res., 6: 1561-1575.
PubMed - Kobayashi, N., T. Higuchi, Y. Urano and K. KiKuchi, 1999. Dipeptides containing L arginine analogs new isozyme selective inhibitors of nitric oxide synthase. Biol. Pharm. Bull., 22: 936-940.
Direct Link - Kubes, P., M. Suzuki and D.N. Granger, 1991. Nitric oxide: An endogenous modulator of leukocyte adhesion. Proc. Natl. Acad. Sci., USA., 88: 4651-4655.
Direct Link - Lee, Y., M.A. Marletta, P. Martasek, L.J. Roman, B.S.S. Masters and R.B. Silverman, 1999. Conformationally restricted arginine analogues as alternative substrates and inhibitors of nitric oxide synthases. Bioorg. Med. Chem., 7: 1097-1104.
PubMed - Lee, Y., P. Martasek, L.J. Roman, B.S.S. Masters and R.B. Silverman, 1999. Imidazole containing amino acids as selective inhibitors of nitric oxide synthases. Bioorg. Med. Chem., 7: 1941-1951.
CrossRef - Lipton, S.A., N.J. Sucher, P.K. Kaiser and E.B. Dreyer, 1991. Synergistic effects of HIV coat protein and NMDA receptor mediated neurotoxicity. Neuron., 7: 111-118.
PubMed - Lowe III, J.A., W. Qian, R.A. Volkmann, S. Heck and J. Nqwakowski, 1999. A new class of selective and potent inhibitors of neuronal nitric oxide synthase. Bioorg. Med. Chem. Lett., 9: 2569-2572.
PubMed - MacIntyre, I., M. Zaidi, A. Towhidul, H.K. Datta and B.S. Moonga et al, 1991. Osteoclastic inhibition an action of nitric oxide not mediated by cyclic gmp. Proc. Natl. Acad. Sci., 88: 2936-2940.
Direct Link - Malliani, A., P.J. Schwartz and A. Zanchetti, 1969. A sympathetic reflex elicited by experimental coronary occlusion. Am. J. Physiol., 217: 703-709.
PubMed - Malliani, A., 1982. Cardiovascular sympathetic afferent fibers. Rev. Physiol. Biochem. Pharmacol., 94: 11-74.
CrossRef - Marletta, M.A., 1994. Approaches toward selective inhibition of Nitric oxide synthase. J. Med. Chem., 37: 1899-1907.
CrossRef - Marsden, P.A., H.H. Heng, S.W. Scheree and A.V. Hall et al., 1993. Structure and chromosomal localization of the human constitutive endothelial nitric oxide synthase gene. J. Biol. Chem., 268: 17478-17488.
Direct Link - Matsuda, H., T. Kageura, T. Morikawa, I. Toguchida, S. Harima and M. Yoshikawa, 2000. Effects of stilbene constituents from rhubarb on Nitric oxide production in lipopolysaccharide activated macrophages. Bioorg. Med. Chem. Lett., 10: 323-327.
PubMed - Mayer, B., P. Klatt, E.R. Werner and k. Schmidt, 1994. Molecular mechanisms of inhibition of porcine brain nitric oxide synthase by the antinociceptive drug 7 nitro indazole. Neuropharmacol., 33: 1253-1259.
PubMed - Mayer, B. and E.R. Werner, 1995. In search of a function for tetrahydrobiopterin in the biosynthesis of nitric oxide. Naunyn Schmiedebergs Arch. Pharmacol., 351: 453-463.
CrossRef - McMillan, K. and B.S.S. Masters, 1993. Optical difference spectrophotometry as a probe of rat brain nitric oxide synthase heme substrate interaction. Biochemistry, 32: 9875-9880.
PubMed - Middleton, S.T., M. Shjorthouse and J.O. Hunter, 1993. Increased Nitric oxide synthesis in ulcerative colitis. Lancet, 341: 465-466.
Direct Link - Misko, T.P., W.M. Moore, T.P. Kasten, G.A. Nickols and J.A. Corbett et al., 1993. Selective inhibition of the inducible Nitric oxide synthase by aminoguanidine. Eur. J. Pharmacol., 233: 119-125.
Direct Link - Mohan, P., D.L. Brutsaert, W. Paulus and S.U. Sys, 1996. Myocardial response to nitric oxide and cGMP. Circulation, 93: 1223-1229.
PubMed - Moncada, S., R.M.J. Palmer and E.A. Higgs, 1991. Nitric oxide physiology pathophysiology and pharmacology. Pharmacol. Rev., 43: 109-142.
PubMed - Moncada, S. and A. Higgs, 1993. The L-arginine-nitric oxide pathway. N. Engl. J. Med., 329: 2002-2012.
CrossRefPubMedDirect Link - Moore, P.K., P. Wallace, Z. Gaffen, S.L. Hart and R.C. Babbedge, 1993. Characterization of the novel nitric oxide synthase inhibitor 7 nitroindazole and related indazoles antinociceptive and cardiovascular effects. Br. J. Pharmacol., 110: 219-224.
Direct Link - Moore, P.K. and R.L.C. Handy, 1997. Selective inhibitors of neuronal nitric oxide synthase is no NOS really good NOS for the nervous system. Trends. Pharmacol. Sci., 18: 204-211.
PubMed - Moore, W.M., R.K. Webber, K.F. Fok, G.M. Jerome and J.R. Connor et al., 1996. Iminopiperidine and other 2 iminoazaheterocycles as potent inhibitors of human Nitric oxide synthase isoforms. J. Med. Chem., 39: 669-672.
CrossRef - Moore, W.M., R.K. Webber, K.F. Fok, G.M. Jerome, C.M. Kornmeier, F.S. Tjoeng and M.G. Currie. 1996. Inhibitors of human nitric oxide synthase isoforms with the carbamidine moiety as a common structural element. Bioorg. Med. Chem., 4: 1559-1564.
PubMed - Murakami, A., G. Gao, M. Omura, M. Yano and C. Ito et al., 2000. Dimethylallylcoumarins potently suppress both lipopolysaccharide and interferon g induced Nitric oxide generation in mouse macrophage RAW 264.7 cells. Bioorg. Med. Chem. Lett., 10: 59-62.
PubMed - Nakane, M., V. Klinghofer, J.E. Kuk, J.L. Donnelly and G.P. Budzik et al., 1995. Novel potent and selective inhibitors of inducible nitric oxide synthase. Mol. Pharmacol., 47: 831-834.
PubMedDirect Link - Oberg, B. and P.N. Thoren, 1973. Circulatory responses to stimulation of medullated and non medullated afferents in the cardiac nerve in the cat. Acta. Physiol. Scand., 87: 121-132.
PubMed - Ogden, J.E. and P.K. Moore, 1995. Inhibition of nitric oxide synthase potential for a novel class of therapeutic agent. Trends. Biotech., 13: 70-78.
PubMed - Pan, H.L., J.C. Longhurst, J.C. Eisenach and S.R. Chen, 1999. Role of protons in activation of cardiac sympathetic C fibre afferents during ischaemia in cats. J. Physiol., 518: 857-866.
CrossRef - Petros, A., D. Bennett and P. Vallance, 1991. Effect of nitric oxide synthase inhibitors on hypotension in patients with septic shock. Lancet, 338: 1557-1558.
PubMed - Pfeiffer, S., A.C.F. Gorren, E. Pitters, K. Schmidt, E.R. Werner and B. Mayer, 1997. Allosteric modulation of rat brain nitric oxide synthase by the pterin site enzyme inhibitor 4 aminotetrahydrobiopterin. Biochem. J., 328: 349-352.
Direct Link - Reif, D.W., D.J. McCarthy, E. Gregan and J.E. Macdonald, 2000. Discovery and development of neuronal nitric oxide synthase inhibitors. Free Radical Biol.Med., 28: 1470-1477.
CrossRef - Reimann, KA. and L.C. Weaver, 1980. Contrasting reflexes evoked by chemical activation of cardiac afferent nerves. Am. J. Physiol. Heart Circ. Physiol., 239: 316-325.
Direct Link - Sakuma, I., H. Togashi, M. Yoshioka, H. Saito, M. Yanagida and M. Tamura et al., 1992. NG methyl L arginine an inhibitor of L arginine derived nitric oxide synthesis stimulates renal sympathetic nerve activity in vivo a role for Nitric oxide in the central regulation of sympathetic tone. Circ. Res., 70: 607-611.
PubMed - Salerno, L., V. Sorrenti, F. Guerrera, M.C. Sarva, M.A. Siracusa, D.C. Giacomo and A. Vanella, 1999. N substituted imidazoles as inhibitors of nitric oxide synthase a preliminary screening. Pharmazie, 54: 685-690.
Direct Link - Schmidt, K., G.W. Felmayer, B. Mayer and E.R. Werner, 1999. Preferential inhibition of inducible nitric oxide synthase in intact cells by the 4 amino analogue of tetrahydrobiopterin. Eur. J. Biochem., 259: 25-31.
PubMed - Seo, H.G., I. Takata, M. Nakamura, H. Tatsumi, K. Suzuki, J. Fujii and N. Taniguchi, 1995. Induction of Nitric oxide synthase and concomitant suppression of superoxide dismutases in experimental colitis in rats. Arch. Biochem. Biophys., 324: 41-47.
CrossRef - Shearer, B.G., S. Lee, K.W. Franzmann, H.A.R. White and D.C.J. Sanders et al., 1997. Conformationally restricted arginine analogues as inhibitors of human Nitric oxide synthase. Bioorg. Chem. Lett., 7: 1763-1768.
CrossRef - Shibuki, K. and D. Okada, 1991. Endogenous Nitric oxide release required for long term synaptic depression in the cerebellum. Nature, 349: 326-328.
CrossRef - Southan, G.J., C. Szabo and C. Thiemermann, 1995. Isothioureas, potent inhibitors of Nitric oxide synthases with variable isoform selectivity. Br. J. Pharmacol., 114: 510-516.
Direct Link - Southan, G.J., A.L. Salzman and C. Szabo, 1996. Potent inhibition of the inducible isoform of Nitric oxide synthase by aminoethylselenourea and related compounds. Life Sci., 58: 1139-1148.
CrossRef - Southan, G.J. and C. Szabo, 1996. Selective pharmacological inhibition of distinct NOS isoforms. Biochem. Pharmacol., 51: 383-394.
CrossRef - Southan, G.J., B. Zingarelli, M.O. Connor, A.L. Salzman and C. Szabo, 1996. Spontaneous rearrangement of aminoalkylisothioureas into mercaptoalkylguanidines a novel class of Nitric oxide synthase inhibitors with selectivity towards the inducible isoform. Br. J. Pharmacol., 117: 619-632.
Direct Link - Tilton, R.G., K. Chang, K.S. Hasan, S.R. Smith and J.M. Petrash et al., 1993. Prevention of diabetic vascular dysfunction by guanidines inhibition of Nitric oxide synthase versus advanced glycation end product formation. Diabetes, 42: 221-232.
Direct Link - Tjen, A., S. Looi, H.L. Pan and J.C. Longhurst, 1998. Endogenous bradykinin activates ischaemically sensitive cardiac visceral afferents through kinin B2 receptors in cats. J. Physiol., 510: 633-641.
CrossRef - Looi, S.C.T.A., N.T. Phan and J.C. Longhurst, 2001. Nitric oxide modulates sympathoexcitatory cardiac cardiovascular reflexes elicited by bradykinin. Am. J. Physiol. Heart Circ. Physiol., 281: 2010-2017.
Direct Link - Tracey, W.R., M. Nakane, F. Basha and G. Carter, 1995. In vivo pharmacological evaluation of two novel type II (inducible) Nitric oxide synthase inhibitors. Can. J. Physiol. Pharmacol., 73: 665-669.
PubMed - Ulhaq, S., E.C. Chinje, M.A. Naylor, M. Jaffar, I.J. Stratford and M.D. Threadgill, 1998. S 2 Amino 5 azolylpentanoic acids related to L ornithine as inhibitors of the isoforms of Nitric oxide synthase (NOS). Bioorg. Med. Chem., 6: 2139-2149.
Direct Link - Vallance, P., A. Leone, A. Calver, J. Collier and S. Moncada, 1992. Accumulation of an endogenous inhibitor of Nitric oxide synthesis in chronic renal failure. Lancet, 339: 572-575.
PubMed - Vallance, P. and S. Moncada, 1993. Role of endogenous Nitric oxide in septic shock. New. Horizons., 1: 77-86.
PubMedDirect Link - Wang, C.C., J.E. Lai, L.G. Chen, K.Y. Yen and L.L. Yang, 2000. Inducible nitric oxide synthase inhibitors of Chinese herbs. Part 2: Naturally occurring furanocoumarins. Bioorg. Med. Chem., 8: 2701-2707.
CrossRef - White, K.A. and M.A. Marletta, 1992. Nitric oxide sythase is a cytochrome P 450 type hemoprotein. Biochemistry, 31: 6627-6631.
CrossRef - Wolff, D.J., G.A. Datto, R.A. Samatovicz and R.A. Tempsick, 1993. Calmodulin dependent Nitric oxide synthase mechanism of inhibition by imidazole and phenylimidazoles. J. Biol. Chem., 268: 9425-9429.
Direct Link - Wolff, D.J., G.A. Datto and R.A. Samatovicz, 1993. The dual mode of inhibition of calmodulin dependent Nitric oxide synthase by antifungal imidazole agents. J. Biol. Chem., 268: 9430-9436.
Direct Link - Wolff, D.J. and B.J. Gribin, 1994. Interferon gamma inducible murine macrophage Nitric oxide synthase studies on the mechanism of inhibition by imidazole agents. Arch. Biochem. Biophys., 311: 293-299.
PubMed - Wolff, D.J., A. Lubeskie and S. Umansky, 1994. The inhibition of the constitutive bovine endothelial Nitric oxide synthase by imidazole and indazole agents. Arch. Biochem. Biophys., 314: 360-366.
PubMed - Wolff, D.J. and A. Lubeskie, 1995. Aminoguanidine is an isoform selective mechanism based inactivator of Nitric oxide synthase. Arch. Biochem. Biophys., 316: 290-301.
PubMed - Wright, C.W., D.D. Rees and S. Moncada, 1992. Protective and pathological roles of Nitric oxide in endotoxin shock. Cardiovasc. Res., 26: 48-57.
PubMed