
Ehsan Oskoueian
Institute of Tropical Agriculture, Universiti Putra Malaysia, 43400, Serdang, Selangor, Malaysia
Norhani Abdullah
Institute of Tropical Agriculture, Universiti Putra Malaysia, 43400, Serdang, Selangor, Malaysia
Wan Zuhainis Saad
Department of Microbiology, Faculty of Biotechnology and Biomolecular Sciences, Serdang, Selangor, Malaysia
Abdul Rahman Omar
Institute of Bioscience, Universiti Putra Malaysia, 43400, Serdang, Selangor, Malaysia
Yin Wan Ho
Institute of Bioscience, Universiti Putra Malaysia, 43400, Serdang, Selangor, Malaysia
Asian Journal of Animal and Veterinary Advances
Year: 2014 | Volume: 9 | Issue: 1 | Page No.: 37-46







ABSTRACT
Phorbol Esters (PEs) from Jatropha curcas seeds are known to have toxicity effects on various animal tissues but little is known about their effects on microorganisms residing in the gut systems of animals when the compounds are consumed. Hence, in the present study, the effects of PEs isolated from Jatropha meal on rumen microbial activities were determined. The rumen microbial fermentation, enzyme activity, microbial protein synthesis, rumen microbial population and the morphology of rumen microbes were examined in vitro in the presence of PEs at 0, 1.5, 3, 4.5 and 6% with 200 mg of Panicum maximum as substrate. The results showed that PEs at 1.5 and 3% did not affect the rumen microbial fermentation activities including, dry matter digestibility, in vitro gas production at 24 h, total VFA, acetic: propionic ratio and cellulolytic enzyme activity (carboxymethylcellulase, filter paperase). However, microbial activities were significantly (p<0.05) suppressed at 4.5 and 6% PEs. On the other hand, microbial protein synthesis and microbial population (general bacteria, fungi, protozoa, methanogens, archaea and major cellulolytic bacteria) values were significantly (p<0.05) reduced at all levels of PEs. Morphological examination using scanning electron microscopy revealed the aggregation and membrane disruption of the rumen microbes. The results of this study showed that, although PEs affected the rumen microbial activities in a dose-dependent manner with no significant adverse effects up to 3% but the microbial population was significantly suppressed (p<0.05) at all levels of PEs.
PDF Abstract XML References Citation
Received: March 25, 2013;
Accepted: October 28, 2013;
Published: March 11, 2014
How to cite this article
Ehsan Oskoueian, Norhani Abdullah, Wan Zuhainis Saad, Abdul Rahman Omar and Yin Wan Ho, 2014. Dose-response Effects of Phorbol Esters Isolated from Jatropha Meal on Rumen
Microbial Activities. Asian Journal of Animal and Veterinary Advances, 9: 37-46.
DOI: 10.3923/ajava.2014.37.46
URL: https://scialert.net/abstract/?doi=ajava.2014.37.46
DOI: 10.3923/ajava.2014.37.46
URL: https://scialert.net/abstract/?doi=ajava.2014.37.46
INTRODUCTION
Recently, there has been a growing interest in Jatropha curcas Linn. (J. curcas) seeds for biodiesel production. Therefore J. curcas plantation in a large scale has been established in many countries including Malaysia. In the oil extraction process, the leftover residue, called Jatropha meal which contained about 60% protein (Oskoueian et al., 2011), is a potential protein supplement in animal production. However, the presence of Phorbol Esters (PEs) (3 mg g-1 DM) in the local genotype renders the meal unsuitable as an animal feed (Makkar and Becker, 2009). Phorbol esters are diterpenes and thought to be the toxic metabolites present in the Jatropha meal. Li et al. (2010) and Rakshit et al. (2008) observed the adverse effects of PEs when mice and rats were fed with diets containing PEs. Numerous attempts have been made using physical, chemical and biological treatments to remove the PEs from Jatropha meal with removal efficiency in the range of 75-94% (Aregheore et al., 2003; Martinez-Herrera et al., 2006; Rakshit et al., 2008; Oskoueian et al., 2011). The presence of residual PEs in the Jatropha meal needs serious consideration on their effects of rumen microbes before the meal can be utilised as an animal feed.
The rumen ecosystem is the first part of the digestive tract expose to the plant materials consumed. Some plant metabolites could be toxic to the rumen microbes when their presence is above the tolerance level of the rumen microbes (Laparra and Sanz, 2010). Due to the lack of agronomically improved varieties of J. curcas devoid of PEs and the incomplete removal of PEs from the meal by various treatments it is necessary to evaluate the tolerance levels of PEs by the rumen microbes. Hence, in the current study, the effects of PEs isolated from Jatropha meal on rumen microbial activities were evaluated. This information would determine the inclusion rate of Jatropha meal in the ruminant diet which show no adverse effects on the rumen microbial population and their activities.
MATERIALS AND METHODS
Plant materials: The seeds of J. curcas were obtained from Faculty of Agriculture, Universiti Putra Malaysia. The seeds were dehulled and the kernels were ground using a mechanical grinder. Then, the ground kernel was subjected to oil extraction with the Soxhlet apparatus as described by Martinez-Herrera et al. (2006). Jatropha meal was air dried and kept at -20oC for PEs isolation.
Phorbol esters isolation: The isolation of PEs form Jatropha meal was conducted as described by Li et al. (2010). Methanol was used to extract the PEs and the crude methanolic extract was injected to the High-Performance Liquid Chromatography (HPLC) (Waters, Milford, MA, USA). The column was a reverse-phase, C18 LiChrospher 100, 250x4 mm I.D with 5 μm pore size (Agilent Technologies, Germany). The solvents consisted of deionized water and acetonitrile and the separation was carried out using a gradient elution. The peaks appeared at 24.4, 25.5, 26.5 and 26.9 min measured 280 nm. Each of the peaks was collected individually using a fraction collector (Waters, Milford, MA, USA). The fractions were pooled and dried using a freeze dryer. The isolated PEs were dissolved in ethanol and analysed again to confirm the concentration and purity. The concentration of the isolated PEs used in this study was expressed as equivalent to a standard, phorbol-12-myristate 13-acetate (PMA).
In vitro rumen fermentation: Rumen liquor was collected before the morning feed from two rumen fistulated Kedah-Kelantan male cattle (averaged 209 kg BW) fed with 0.025 of BW DM day-1 of forage (Panicum maximum) and commercial cattle concentrate at 60:40 ratio. The incubation medium was prepared as described by Menke and Steingass (1988) and 30 mL was dispensed anaerobically into each 100 mL syringe (Häberle Labortechnik, Lonsee-Ettlenschieß, Germany). As substrate, 200 mg of milled freeze-dried P. maximum at 1 mm size were used. Standard hay (University of Hohenheim, Stuttgart, Germany) with an estimated gas production of 49.61 mL g-1 DM was used as a standard to check the quality of inoculum and to calibrate the in vitro gas production system. Four levels of PEs at 1.5, 3, 4.5 and 6% (w/w) of the substrate on dry matter basis were tested. Ethanol was used as the carrier with the concentration of 0.3% (v/v) of buffered rumen fluid. The treatments were as I= grass, II = grass+ethanol (as a carrier), III = grass+1.5% PEs, IV = grass+3% PEs, V = grass+4.5% PEs, VI = grass+6% PEs. A total of nine syringes for each treatment were used. The content of three syringes were used for DM Degradability (DMD), pH, fermentation parameters and another three for microbial protein synthesis and the remaining three syringes were used for the rumen microbial population quantification and enzyme activity assays. The above procedures were conducted in three individual runs. In vitro gas production was measured at 2, 4, 8, 12 and 24 h. Cumulative gas production data were fitted to the model described by Orskov and McDonald (1979) using NEWAY Excel version 5.0 package (Chen, 1997).
Rumen microbial enzyme activity: The content of the syringe after 24 h of incubation was transferred into glass beakers and mixed with 2.5 mL of carbon tetrachloride and 2.5 mL of lysozyme solution (4 g L-1 of phosphate buffer, 0.1 mol, pH 6.8) and the mixture was incubated at 40°C for 3 h followed by sonication at 4°C using a sonicator (LabSonic, B. Braun, Germany). The sonicated samples were centrifuged at 20,000xg for 25 min at 4°C and supernatant was collected and used for enzyme assays (Patra et al., 2010). The carboxymethylcellulase (CMCase), Filter Paperase (FPase), β-glucosidase and xylanase activities were determined and the specific activity of each enzyme was expressed as μmol of product (glucose/xylose) released min-1 mg-1 protein under the assay conditions (Zuhainis Saad et al., 2008). The protein content was determined based on Bradford assay using the Bio-Rad Protein Assay Kit II (Bio-Rad, USA).
Rumen microbial protein synthesis: The purines contents in apparent undegraded residue left after fermentation was determined as described by Makkar and Becker (1999). Briefly, the whole content of syringe was lyophilized and 50 mg of undigested residue was digested using perchloric acid (0.6 M). The digested sample was filtered through 0.45 μm filter and inject to the high-performance liquid chromatography (HPLC Agilent-1200) series instrument equipped with an analytical column Agilent Reverse-phase C18 LiChrospher 100 (250x4 mm I.D and 5 μm pore size). The separation was performed using a gradient elution with solvents comprising of solvent A (10 mM NH4H2PO4, pH 6) and solvent B (150 mL of acetonitrile added to 600 mL of 12.5 mM NH4H2PO4, pH 6) with the flow rate of 0.8 mL min-1. The absorbance was read at 254 nm. Guanine and adenine peaks appeared at about 8.3 and 11.1 min, respectively. Allopurinol was used as internal standards and appeared at about 16.6 min.
Rumen microbial population analysis using real time PCR: At the end of incubation, 1 mL of rumen fluid containing digesta was used for DNA extraction using QIAamp DNA Stool Mini Kit (QIAGEN). The 16S rRNA of bacteria and archaea and 18S rRNA of protozoa and fungi were amplified by PCR using primers for general bacteria, general fungi, total protozoa (Sylvester et al., 2004), Ruminococcus flavefaciens, Fibrobacter succinogenes and Butyrivibrio fibrisolvens (Denman and McSweeney, 2006), Ruminococcus albus (Koike and Kobayashi, 2001), Butyrivibrio fibrisolvens (Stevenson and Weimer, 2007) total methanogens and total archaea (Yu et al., 2005). The PCR products were cloned in pCR®2.1-TOPO® TA cloning vector (Invitrogen, Carlsbad, CA, USA) and transformed into chemically competent E. coli TOP10 cells (Invitrogen). The plasmid was extracted (iNtRON Biotechnology, Sungnam, Kyungki-Do, Korea) and sequenced using capillary electrophoresis on an Applied Biosystems 3730xl DNA Analyzer (Applied Biosystems, Foster City, CA, USA). The sequences were checked for chimeric rDNA (Huber et al., 2004) and were compared to those available in the GenBank using the Basic Local Alignment Search Tool (Altschul et al., 1997). The plasmid carrying the sequences that were = 99% similar to the previously published sequence were used for real time PCR to construct the standard curve. Real-time PCR assays were conducted on a BioRad CFX 96 real-time PCR thermocycler (Bio-Rad, Hercules, United States of America) using iQ SYBR Green Supermix (Bio-Rad Laboratories, Inc., Hercules, CA). Data from the real-time PCR reactions were analyzed using CFX manager software version 2 (Bio-Rad Laboratories). All real-time PCR amplifications were performed in triplicate.
Morphological characteristic of rumen microbes: The morphology of bacteria, fungi and protozoa in the rumen fluid obtained from treatment II (grass+ethanol as a carrier) and VI (grass+6% PEs) upon 24 h incubation were examined using a Scanning Electron Microscopy (SEM). The rumen microbes were fixed using a standard procedure (Chong et al., 2009). Then, the samples were transferred to a specimen basket and dried in a critical dryer for 35 min. The dried samples were mounted on SEM stabs, sputter-coated with gold and observed using a JEOL-6-400 microscope at various magnifications.
Statistical analyses: The GLM procedure of SAS (2003) was used for the statistical analysis using a complete randomized design following the model:
Yi = μ i+ei
where, μ is the value of the mean, Ti is the effect of treatment and ei is the error of experiment, respectively and means were compared with Duncan’s Multiple Range test (p<0.05).
RESULTS
Isolation of phorbol esters: The result of high performance liquid chromatography (HPLC) analysis showed the presence of four PEs present in Jatropha meal. They were named as PE1, PE2, PE3 and PE4 (Fig. 1). The retention times were similar to that of PEs reported by Makkar et al. (2007) and Li et al. (2010).
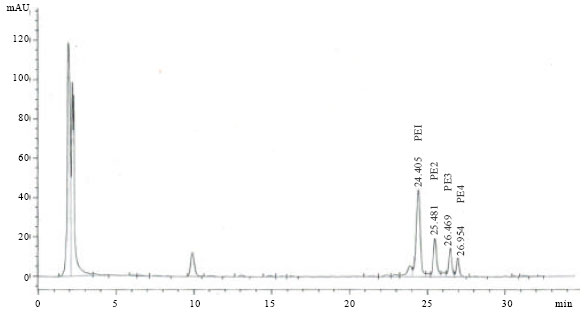 | |
| Fig. 1: | HPLC chromatogram of the PEs present in Jatropha meal. Four peaks (PE1, PE2, PE3 and PE4) correspond to four PEs compounds |
Hass et al. (2002) have also characterised the PEs of J. curcas seed and confirmed the presence of six PEs. In the present analysis, only four peaks were observed, possibly due to the similar molecular weight of some of the PEs present in Jatropha meal. The concentrations of the isolated PEs used in this study were expressed as equivalents to the standard phorbol-12-myristate 13-acetate (PMA). The concentration of PEs in Jatropha meal was 3 mg PMA equivalent/g dry matter of Jatropha meal. The PEs were pooled, thus the response of rumen microbes observed in this experiment corresponded to the total PEs present in Jatropha meal.
Effects of isolated PEs on rumen fermentation: In vitro rumen fermentation parameters are presented in Table 1. The PEs carrier (ethanol) did not have any effect on gas production parameters which indicated that the level of carrier used in the experiment (0.3% v/v) was not detrimental to rumen microbes. The Dry Matter Digestibility (DMD), Metabolisable Energy (ME) and total volume of gas produced after 24 h of incubation (IVGP24) decreased significantly (p≤0.05) when the inclusion of the PEs reached to 4.5 and 6% of the substrate on dry matter basis. Fraction (a) and (c) in gas production parameters were not affected by PEs significantly while the fractions (a+b) and (b) were depressed significantly (p≤0. 05) at 6% PEs.
Fermentation parameters are shown in Table 2. The pH values and ammonia nitrogen were not significantly different among treatments. However, the total VFA produced by rumen microbes and the ratio of acetic to propionic acid were significantly (p≥0.05) suppressed at 4.5 and 6% addition rates as compared to the controls (I and II).
| Table 1: | Effects of PEs on rumen microbial fermentation parameters |
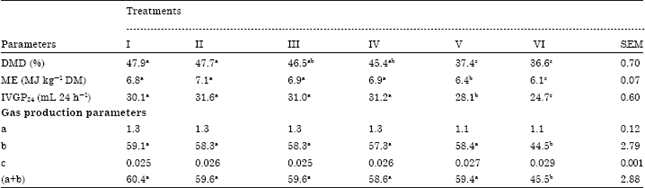 | |
| a: Gas production from the immediately soluble fraction, b: Gas production from the insoluble fraction, c: Gas production rate constant for the insoluble fraction (b), (a+b): Potential extent of gas production, I: Grass, II: Grass+ethanol (as a carrier), III: Grass+1.5% PEs, IV: Grass + 3% PEs, V: Grass+4.5% PEs, VI: Grass+6% PEs. Means in each row with different superscripts are significantly different (p≤0.05) | |
| Table 2: | Effects of PEs on ruminal pH, ammonia and volatile fatty acids |
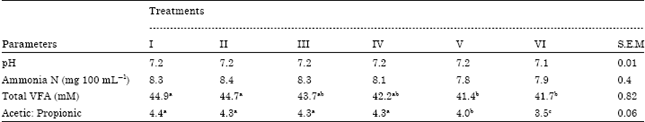 | |
| I: Grass, II: Grass+ethanol (as a carrier), III: Grass+1.5% PEs, IV: Grass+3% PEs, V: Grass+4.5% PEs, VI: Grass+6% PEs, Means in each row with different superscripts are significantly different (p≤0.05) | |
Rumen microbial enzyme activity: Significant (p≤0.05) reductions in CMCase, FPase, xylanase and β-glucosidase enzymes activity in the presence of PEs were observed (Table 3). The activities of microbial CMCase and FPase enzymes were found to be significantly suppressed at high concentrations of PEs (4.5 and 6%) while the activities of microbial xylanse and β-glucosidase enzymes appeared to be significantly (p≤0.05) affected even in the lower concentration of PEs.
Rumen microbial protein synthesis: The result showed that addition of PEs at the concentration of 1.5, 3, 4.5 and 6% of substrate led to significant (p≤0.05) decrease in purine content as an index for rumen microbial protein (Table 4). The total gas and total VFA productions values were incorporated to determine the efficiency of rumen microbial protein synthesis and the values indicated significant reduction in the presence of 4.5 and 6% inclusion of PEs.
Rumen microbial population: The rumen microbial population analysis results are shown in Table 5. These results were consistent with gas production, enzyme activity and microbial protein synthesis data, where all concentrations of PEs were detrimental to rumen microbes with significant (p≤0.05) reduction in the population of general bacteria, protozoa, fungi, methanogens, archaea and major cellulolytic bacteria.
| Table 3: | Effects of PEs on the specific activity of enzymes in buffered rumen fluid |
 | |
| *: μmol min-1 mg-1 protein, I: Grass, II: Grass+ethanol (as a carrier), III: Grass+1.5% PEs, IV: Grass+3% PEs, V: Grass+4.5% PEs, VI: Grass+6% PEs, Means within the same row with different superscripts are significantly different (p≤0.05) | |
| Table 4: | Effects of PEs on purine content (index of microbial protein) and efficiency of rumen microbial protein synthesis |
 | |
| I: Grass, II: Grass+ethanol (as a carrier), III: Grass+1.5% PEs, IV: Grass+3% PEs, V: Grass+4.5% PEs, VI: Grass+6% PEs, Means in the same row with different superscripts are significantly different (p≤0.05) | |
| Table 5: | Effect of isolated PEs on rumen microbial population |
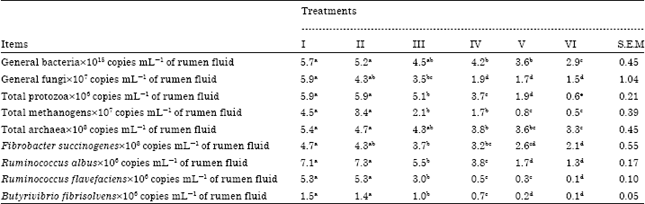 | |
| I: Grass, II: Grass+ethanol (as a carrier), III: Grass+1.5% PEs, IV: Grass+3% PEs, V: Grass+4.5% PEs, VI: Grass+6% PEs. Means within the same row with different superscripts are significantly different (p≤0.05) | |
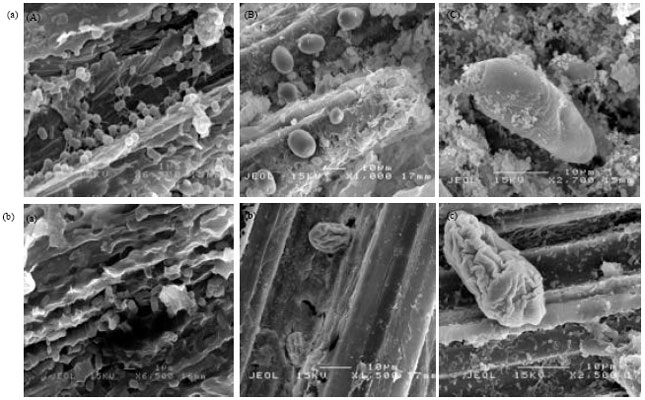 | |
| Fig. 2(a-b): | Scanning electron micrographs of rumen microbes showing the morphology of rumen bacteria, fungi and protozoan, respectively (a) in the absence of PEs (A, B and C) and (b) in the presence of PEs (a, b and c) at the concentration of 6% of the substrate upon 24 h fermentation |
Morphological characteristic of rumen microbes: The morphology of ruminal bacteria (A), fungi (B) and protozoa (C) in the rumen fluid upon 24 h incubation without PEs are shown in Fig. 2. The rumen microbes appeared to be normal and in the good condition as indicated by their attachment, uniformity and intact cell walls. It seems that, the presence of PEs reduced the attachment of microbes to the plant material, affect the population and distribution of rumen bacteria (a) fungi (b) and protozoa (c). In addition, aggregation and membrane disruption were also observed in these microorganisms.
DISCUSSION
The results showed that PEs level of 3% and above reduced the purine content and rumen microbial population but the effects of PEs on dry matter degradability, metabolisable energy, total VFA, gas production and enzyme activities were only apparent with higher levels of PEs (4.5 and 6%). This difference might be attributed to the variation in the susceptibility of rumen microorganism to the presence of PEs. The result obtained in this study was consistent with the results reported by Makkar and Becker (2010) who observed no alteration in rumen fermentation with the incorporation of 1.5% PEs.
A wide range of antimicrobial properties of the PEs has been reported (Chumkaew et al., 2003; Saetae and Suntornsuk, 2010), where responses are suggested to be triggered by the attachment of PEs functional groups to the surface receptors of microorganisms. Most probably, the hydrophobic nature of PEs and their affinity toward lipids could possibly the reasons to disrupt the cytoplasmic membrane either directly or by damaging the membrane proteins, causing increased membrane permeability and finally leakage of cytoplasmic constituents (Gomperts et al., 2009). Phorbol esters may also cause cytoplasmic coagulation and finally lead to cell lysis similar to other terpenoids (Abdel-Hafez et al., 2002; Chumkaew et al., 2003). This reported findings augur well with the result observed in the present study including disruption and aggregation of bacteria, fungi and protozoa cell walls upon incubation with PEs. Its likely that the fungi and protozoa were more susceptible to the presence of PEs as compared to the bacteria. At this stage it is not known that the detrimental effects of PEs at high inclusion rates (4.5 and 6%) on rumen microbial activities were due to the direct effects of PEs or by metabolites produced upon biodegradation or biotransformation of the PEs.
CONCLUSION
The results of this study suggested that, PEs of Jatropha meal affected rumen microbial activities in a dose-dependent manner with no significant adverse effects below 3% on dry matter basis of substrate used. However, the rumen microbial population was significantly suppressed (p<0.05) at all levels of PEs.
ACKNOWLEDGMENTS
The research grant provided by the Ministry of Science, Technology and Innovation, Malaysia (Project No. 02-01-04-SF1132) is gratefully acknowledged.
REFERENCES
- Abdel-Hafez, A., N. Nakamura and M. Hattori, 2002. Biotransformation of phorbol by human intestinal bacteria. Chem. Pharm. Bull., 50: 160-164.
PubMedDirect Link - Altschul, S.F., T.L. Madden, A.A. Schaffer, J. Zhang, Z. Zhang, W. Miller and D.J. Lipman, 1997. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res., 25: 3389-3402.
CrossRefPubMedDirect Link - Aregheore, E.M., K. Becker and H.P.S. Makkar, 2003. Detoxification of a toxic variety of Jatropha curcas using heat and chemical treatments and preliminary nutritional evaluation with rats. S. Pac. J. Nat. Sci., 21: 51-56.
CrossRefDirect Link - Chong, M.L., N.A.A. Rahmana, R.A. Rahim, S.A. Aziz, Y. Shirai and M.A. Hassan, 2009. Optimization of biohydrogen production by Clostridium butyricum EB6 from palm oil mill effluent using response surface methodology. Int. J. Hydrogen Energy, 34: 7475-7482.
CrossRefDirect Link - Denman, S.E. and C.S. McSweeney, 2006. Development of a real-time PCR assay for monitoring anaerobic fungal and cellulolytic bacterial populations with in the rumen. FEMS Microbiol. Ecol., 58: 572-582.
Direct Link - Haas, W., H. Sterk and M. Mittelbach, 2002. Novel 12-deoxy-16-hydroxyphorbol diesters isolated from the seed oil of Jatropha curcas. J. Nat. Prod., 65: 1434-1440.
CrossRefDirect Link - Huber, T., G. Faulkner and P. Hugenholtz, 2004. Bellerophon: A program to detect chimeric sequences in multiple sequence alignments. Bioinformatics, 20: 2317-2319.
PubMed - Koike, S. and Y. Kobayashi, 2001. Development and use of competitive PCR assays for the rumen cellulolytic bacteria: Fibrobacter succinogenes, Ruminococcus albus and Ruminococcus flavefaciens. FEMS Microbiol. Lett., 204: 361-366.
CrossRefPubMedDirect Link - Laparra, J.M. and Y. Sanz, 2010. Interactions of gut microbiota with functional food components and nutraceuticals. Pharmacol. Res., 61: 219-225.
CrossRef - Li, C.Y., R.K. Devappa, J.X. Liu, J.M. Lv, H.P.S. Makkar and K. Becker, 2010. Toxicity of Jatropha curcas phorbol esters in mice. Food Chem. Toxicol., 48: 620-625.
CrossRefDirect Link - Makkar, H.P. and K. Becker, 1999. Purine quantification in digesta from ruminants by spectrophotometric and HPLC methods. Br. J. Nutr., 81: 107-112.
PubMed - Makkar, H.P.S. and K. Becker, 2009. Jatropha curcas, a promising crop for the generation of biodiesel and value-added coproducts. Eur. J. Lipid Sci. Technol., 111: 773-787.
CrossRefDirect Link - Makkar, H.P.S. and K. Becker, 2010. Are Jatropha curcas phorbol esters degraded by rumen microbes? J. Sci. Food Agric., 90: 1562-1565.
CrossRefDirect Link - Martinez-Herrera, J., P. Siddhuraju, G. Francis, G. Davila-Ortiz and K. Becker, 2006. Chemical composition, toxic/ antimetabolic constituents and effects of different treatments on their levels, in four provenances of Jatropha curcas L. from Mexico. Food Chem., 96: 80-89.
Direct Link - Menke, K.H. and H. Steingass, 1988. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev., 28: 7-55.
Direct Link - Orskov, E.R. and I. McDonald, 1970. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agric. Sci., 92: 499-503.
CrossRefDirect Link - Oskoueian, E., N. Abdullah, W.Z. Saad, A.R. Omar, M.B. Puteh and Y.W. Ho, 2011. Anti-nutritional metabolites and effect of treated Jatropha curcas kernel meal on rumen fermentation in vitro. J. Anim. Vet. Adv., 10: 214-220.
CrossRefDirect Link - Patra, A.K., D.N. Kamra and N. Agarwal, 2010. Effects of extracts of spices on rumen methanogenesis, enzyme activities and fermentation of feeds in vitro. J. Sci. Food Agric., 90: 511-520.
Direct Link - Rakshit, K.D., J. Darukeshwara, K.R. Raj, K. Narasimhamurthy, P. Saibaba and S. Bhagya, 2008. Toxicity studies of detoxified Jatropha meal (Jatropha curcas) in rats. Food Chem. Toxicol., 46: 3621-3625.
CrossRefDirect Link - Saetae, D. and W. Suntornsuk, 2010. Antifungal activities of ethanolic extract from Jatropha curcas seed cake. J. Microbiol Biotechn., 20: 319-324.
PubMedDirect Link - Sylvester, J.T., S.K.R. Karnati, Z. Yu, M. Morrison and J.L. Firkins, 2004. Development of an assay to quantify rumen ciliate protozoal biomass in cows using real-time PCR. J. Nutr., 134: 3378-3384.
Direct Link - Stevenson, D.M. and P.J. Weimer, 2007. Dominance of prevotella and low abundance of classical ruminal bacterial species in the bovine rumen revealed by relative quantification real-time PCR. Appli. Microbiol Biot., 75: 165-174.
PubMed - Yu, Y., C. Lee, J. Kim and S. Hwang, 2005. Group-specific primer and probe sets to detect methanogenic communities using quantitative real-time polymerase chain reaction. Biotechnol. Bioeng., 89: 670-679.
PubMed