
S. Sutharshiny
Department of Fisheries, University of Jaffna, Jaffna, Sri Lanka
K. Sivashanthini
Department of Zoology, University of Jaffna, Jaffna, Sri Lanka
W.S. Thulasitha
Department of Zoology, University of Jaffna, Jaffna, Sri Lanka
Asian Journal of Animal and Veterinary Advances
Year: 2013 | Volume: 8 | Issue: 4 | Page No.: 555-570







ABSTRACT
The present study was undertaken to understand the lipid changes in gonad, muscle and liver tissues of tropical double spotted queen fish, Scomberoides lysan (Family: Carangidae) in relation to sexual maturity and spawning. Cholesterol (CS), phospholipid (PL) and triacylgycerol (TAG) were determined in gonad, muscle and liver tissues with respect to maturity stages of both sexes as well as months. Fish were periodically caught from waters around Sri Lanka throughout the year 2010 to 2011. Fish length, weight, sex and maturation status were recorded. Content of CS, PL and TAG in gonad, muscle and liver tissues were determined at the laboratory. The values of CS, PL and TAG in the ovary increased to 2-5 fold throughout the ovarian maturation and decreased to 2-8 fold after spawning, whereas in liver and muscle tissue, increased up to maturation and decreased during spawning. Similar dynamics was recorded in males. The main lipid constituents in the liver and muscle of mature fish were TAG and PL, respectively. Lipid constituents in gonads showed higher value, whereas muscle and liver showed lower value in June and September, which represent the spawning time of S. lysan. It has been concluded that the values of lipid in tissues of S. lysan influence the cycle of maturation and time of spawning. This new information can be used for the determination of the fishing season for S. lysan, when it is not reproductively active and has high nutritional value in terms of lipid. The knowledge gained on CS, PL and TAG in different maturity stages of S. lysan can also be utilized in broodstock diet formulation in the future culture trials of S. lysan.
PDF Abstract XML References Citation
Received: March 19, 2013;
Accepted: May 03, 2013;
Published: June 27, 2013
How to cite this article
S. Sutharshiny, K. Sivashanthini and W.S. Thulasitha, 2013. Lipid Changes in Relation to Maturation and Spawning of Tropical Double Spotted Queenfish, Scomberoides lysan (Forsskål, 1775). Asian Journal of Animal and Veterinary Advances, 8: 555-570.
DOI: 10.3923/ajava.2013.555.570
URL: https://scialert.net/abstract/?doi=ajava.2013.555.570
DOI: 10.3923/ajava.2013.555.570
URL: https://scialert.net/abstract/?doi=ajava.2013.555.570
INTRODUCTION
In fish, lipids are known to be an important energy source for reproduction, since large amounts of lipids are required both for female egg production and for male breeding activities, such as enhanced swimming activity, competition, courtship, parental care and nesting (Goda et al., 2007; Ebrahimnezhadarabi et al., 2011).
When a spawning migration is involved, the adult fish generally deplete all their reserves and die after spawning as in the case of Sockeye Salmon (Oncorhynchus nerka) (Hinch et al., 2006) and eel (Anguillia anguillia) (Fricke and Kaese, 1995). If no migration is involved, fish are capable of building their energy reserves completely after spawning.
The lipid storage tissue varies depending on the fish species; some species deposit in liver (most gadoids; Alonso-Fernandez and Saborido-Rey, 2012) while some in muscle (Antarctic fish; Clarke et al., 1984) and some in both liver and muscle (Jezierska et al., 1982; Hedayatifard and Yousefian, 2010).
Fluctuation of lipid classes in gonad, muscle and liver of adult fish is directly associated with sexual maturity and spawning of fish (Mourente et al., 2002; Huynh et al., 2007). Knowledge gained from lipid changes in different tissues of species would be helpful to understand the physiology and ecology of that species. Due to higher investment of resources for reproduction, lipid reserves in liver and muscle are mobilized and transferred to the gonad during maturation and spawning (Zaboukas et al., 2006; Sutharshiny and Sivashanthini, 2011a; Singh et al., 2012). Further, variations in lipid composition in fish tissues depend on environmental conditions (Lund et al., 2000) and seasonal variations (Kandemir, 2010).
The length of the spawning season and spawning frequencies varies greatly between species. Some species for example Salmoniform, Atheriniform and Tetraodontiform have a marked seasonal periodicity in gonadal maturation (Taylor, 1984), while species of Blennius pholis has ripe ovaries throughout the year (Qasim, 1957). Some fish spawn only once a year or once in their lifetime (e.g., most Oncorhynchus spp. and Anguilla spp., De Vlaming, 1983) while others spawn several times a year for example Black and White bream (Jacques and Patrick, 2003), Latris lineate (Bransden et al., 2007) and Scombroides lysan (Thulasitha and Sivashanthini, 2013).
Lipids are complex classes of compounds, can be broadly divided into two groups, one is polar lipids composed principally of phospholipids and the other one is non polar lipids composed principally of triacylglycerols and cholesterol (Tocher, 2003). These components affect the biochemical processes of organism at different level. Phospholipid is the main lipid of cellular membranes and important constituents of egg yolk in fish (Johnson, 2009). It can also be an important source of energy (fatty acids) in fish, particularly during embryonic and early larval development in species that produce phospholipid rich eggs (Tocher, 1995). Triacylglycerol is the major energy storage form in fish (Shulman, 1974) and stored in liver, muscle and mesenteric fat (Sheridan, 1994). Cholesterol is a precursor for the steroid hormones and bile acids (Scott, 1987) and used for additional cellular functions in the testis (Sharpe et al., 2006).
The Double spotted queenfish (Scomberoides lysan) is a tropical fish and broadly distributed throughout the Indo-Pacific region (Froese and Pauly, 2010; Varghese et al., 2011). It is an economically important food fish in Sri Lanka. The species is popular for dry fish production with export demands and especially consumed by mothers during pregnancy and immediately after delivery. Thus, it is highly prized, continues to maintain a high market demand and marketed preserved, dried or salted (Sutharshiny and Sivashanthini, 2011b; c) and hold an important position towards the economy of the fisheries of Sri Lanka.
Though there are several studies on lipid dynamics in different tissues related to reproduction were recorded for temperate fish species (Fiorin et al., 2007; Lloret et al., 2008) only few studies are available for tropical fish species (Arrington et al., 2006; Hiroaki, 2012; Ovie et al., 2007; Talat et al., 2006) especially for carangids (Ramadan, 2002; Assem et al., 2005). Few research works related to lipid composition of fish were carried out by different authors in Sri Lanka (Anas et al., 2009; Thilakarathne and Attygalle, 2009; Ubhayasekera et al., 2012). However no studies were performed on lipid changes in S. lysan and therefore the present study on variation in lipid classes of S. lysan is the first pilot study in Sri Lanka.
The knowledge gained from the present study on lipid changes of S. lysan based on lipid class constituents-cholesterol, triacylglycerol and phospholipids can be considered and applied in the future studies, contributing to economic and health development and sustainable management of S. lysan in Sri Lanka.
MATERIALS AND METHODS
Sample collection: Regular field visits were made once a month to the landing centers at Jaffna, Trincomalee, Mannar and Puttalam (Fig. 1). From the landed marketed fish, size selective samples were collected monthly from January 2010 to December 2011 with the assistance of fishermen co-operative society’s Union of each landing site. The fish samples collected were actually caught mainly by 17.78 cm 21 ply mesh size, drift nets used particularly for queen fish (Katta valai). Fish samples were also collected from the by catch species caught using 6.35 and 8.89 cm mesh size drift net and seine net. Immediately after collection, fish were chilled before freezing (Graham et al., 1992) and brought to the laboratory in an ice box (Giostyle, Ole 25; Italy).
Morphometric analysis: Fish were allowed to thaw slowly and Standard Length (SL) was determined using measuring tape to the nearest 0.1 mm and Body Weight (BW) was measured using top loading balance to the nearest 0.01 g before conducting lipid analysis.
Sex determination: Sex and gonad maturity stages were determined for each specimen using macroscopic examination of gonad and recorded.
 | |
| Fig. 1: | Sampling stations of S. lysan. A: Jaffna, B: Mannar, C: Trincomalee and D: Puttalum |
Tissues analysis: Fish were dissected and whole gonad and liver were removed and weighed using an electronic balance (OHAUS; USA) to the nearest 0.01 g. The muscle tissue from dorsal side that is directly under the dorsal fin and well above the lateral line was removed. Tissues were dried in an Oven (YCO-010; Germany) at 60°C for 24 h. The tissues were covered with filter paper to prevent accidental weight loss of lipid from tissues and to stop droplets erupting out of the container. The dried tissues were reweighed and ground twice in an electric grinder (Preett XT- 97; India).
Total lipid extraction: Total lipid in each tissue was analysed according to the Bligh and Dyer (1959) method. All chemicals were purchased from standard sources Sigma chemical company, USA. A weight of 10 g dried tissue powder was homogenized with 200 mL of chloroform/methanol mixture that prepared as in the ratio 2:1 (v/v). After dispersion, the whole mixture was agitated for 20 min at 2000 rpm in vortex mix (Karl Hecht KG; Germany) at room temperature. The whole mixture was filtered (funnel with a folded filter paper; Diameter-11 cm). The solids on filter paper were washed with 40 mL of distilled water, then the fluid mixture was vortexed for 1 min and centrifuged (Sigma; Germany) at low speed (2000 rpm) for 10 min to separate the two phases and allowed to stand. A biphasic system was obtained. The upper phase was siphoned, the lower chloroform phase containing lipids was filtered off and the water was removed from the extract by passing it through a folded filter paper containing anhydrous sodium sulphate. The interface was rinsed twice with methanol/chloroform (1:1 v/v). The lower phase containing individual lipids were recovered after evaporating under vacuum in a rotary evaporator (1 KA HB 10 basic; Germany). The dried lipid extracts with a small volume of chloroform-methanol mixture in Kjldhal flask were left to evaporate in the fume chamber. The resulting extract of total lipid was stored in a sealed vial at -20°C for further analysis.
Lipid class analysis: Cholesterol (CS) (Zlatkis et al., 1953), phospholipid (PL) (Zilversmit and Davis, 1950) and triacylglycerol (TAG) (Foster and Dunn, 1973) in different tissues were estimated. Standard curve for each lipid class constituents was plotted using the attached LABOMED, INC software in UV Visible spectrophotometer (LABOMED, UVD-3000). The concentrations of lipid classes in tissues were quantified.
Cholesterol analysis: A weight of 0.1 g extracted lipid was taken. Five milliliter of ferric chloride (in acetic acid) reagent was pipetted into lipid and mixed. Then 3 mL of concentrated sulphuric acid was pippetted into it, mixed again and allowed to stand for 20 min. 0.1 mL of glacial acetic acid was used for blank. The concentration of CS in tissues was read against the blank at 560 nm.
Phospholipid analysis: A weight of 0.1 g extracted lipid was taken into a 150 mL of kjeldhal flask and 1.0 mL of 5 N sulphuric acid was added to digest the lipid in a digestion rack (Sigma; Germany) till the appearance of light brown colour. Three drops of concentrated nitric acid were added to flask and continued the digestion till the brown colour changed into colourless. The Kjeldhal flask was cooled. 0.1 mL of distilled water was added to it and heated in a boiling water bath for 5 min. 1.0 mL of ammonium molybdate and 0.1 mL of amino-2-naphthol-4-sulphonic acid were added and it was transferred to 5 mL volumetric flask and total volume was made upto 5 mL with distilled water. Distilled water was used for blank. The concentration of PL in tissues was read against the blank at 660 nm within 10 min.
Triacylglycerol analysis: A weight of 0.1 g extracted lipid was taken. Four milliliter of isopropanol was added and mixed well. 400 mg of washed alumina was added. The mixture was placed in a mechanical rotator for 15 min and centrifuged. Two milliliter of supernatant was transferred into 15x100 mm of screw-capped tubes. A volume of 0.6 mL of potassium hydroxide was added into supernatant solutions, stoppered and incubated at 70°C for 15 min. Tubes were allowed to cool. 1 mL of metaperiodate solution and 0.5 mL of acetone reagent were added and mixed well; stoppered and incubated at 50°C for 30 min. Tubes were allowed to cool. One milliliter of distilled water was used for blank. The concentration of TAG in tissues was read against the blank at 405 nm.
Data analysis: All data were statistically analyzed by Micro soft Excel (Version 2007) and STATISTICA Soft ware (Version 6; Statsoft Inc.,Tulsa, USA). The data were checked for normal distribution by one-sample kolmogorov-smirno test and the variances were tested by the Levene’s test for homogeneity. Lipid class concentrations in tissues were compared among gonad maturity stages as well as months. Lipid class contents in tissues were first analyzed by one way Analysis of Variance (ANOVA). When the results of the one way ANOVA show the mean values of the samples are significantly different, the ANOVA was followed by Post hoc comparison of means and Duncan’s Multiple Range Test (DMRT) using STATISTICA 6.0 software. The level of statistical significance was set at p<0.05. Monthly analysis of lipid class content in different tissues was conducted only for adult fish (maturing stage to spent stage), which was collected throughout the year except January, November and December for female whereas March, November and December for male. Monthly lipid class data in different tissues for both years were pooled together and the average values for each month were computed.
RESULTS
One thousand four hundred and nineteen fish samples ranging from 10.7 to 67.8 cm in SL were examined and BW of individuals ranged from 21.10 to 2925.00 g. Reproductive status of individual fish was examined and the gonad developmental stages were classified as immature unsex (stage I), immature (stage II), maturing (stage III), mature (stage IV), spawning (stage V) and spent (stage VI) (Table 1).
| Table 1: | Macroscopic features of different gonad maturity stages (GMS) in S. lysan |
 | |
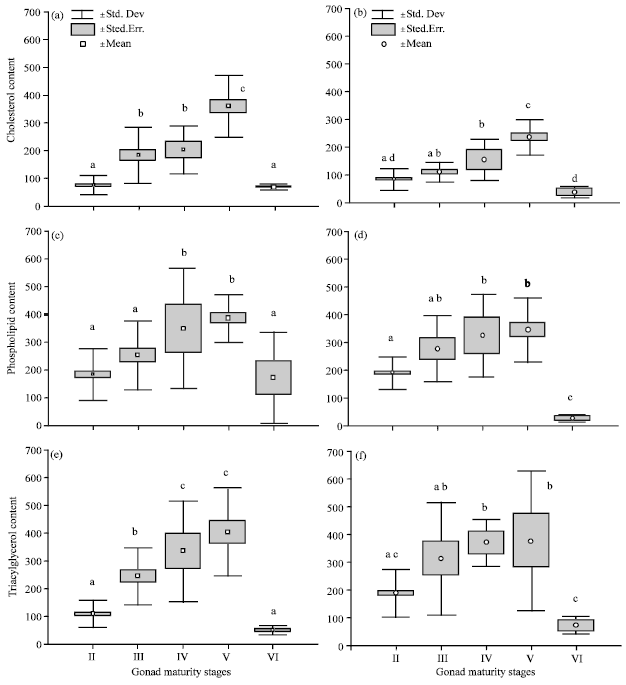 | |
| Fig. 2(a-f): | Box and Whisker plot showing changes of lipid class content (mg.100g-1) in gonad tissue of S. lysan in different gonad maturity stages, II: Immature; III: Maturing; IV: Mature; V: Spawning; VI: Spent. a, c and e-Gonad in female; b, d and f-Gonad in Male. Mean values for each gonad maturity stages with the common letters indicate not significantly difference (p>0.05) |
Lipid changes in tissues with gonad maturity stages
Lipid class content in gonad tissues: The changes in lipid content of tissues for gonad maturity stages were analyzed for the entire data set (2010/2011). The amounts of CS, PL and TAG in the ovary increased throughout the ovarian maturation (stage I-V).
CS content in ovary showed 5 fold increase (p = 4.57E - 05) from stage II to stage V while 5 fold decrease (p = 2.86E-05) from stage V to VI (Fig. 2a). CS content in testis showed approximately 3 fold increase from stage II to V whereas a 6 fold decrease thereafter (Fig. 2b). The mean PL levels of both sexes showed 2 fold increases up to stage V and decreased thereafter (Fig. 2c, d). Content of TAG in ovary showed 4 fold increase (p = 4.68E-05) up to stage V whereas 8 fold decrease (p = 2.94E -05) from stage V to VI (Fig. 2e). Content of TAG in testis showed approximately 2 fold increase at stage V when compared to stage II while a 5 fold decrease from stage V to VI (Fig. 2f).
Lipid class content in muscle tissues: Lipid class concentration in immature unsex (stage I) of S. lysan was higher in muscle tissue compared with liver. PL content of muscle and liver tissues in stage I was higher than that of other lipid classes. The content of CS in muscle of female significantly (p = 0.004) increased from stage I to stage III and significantly (p = 4.05E-06) decreased beyond that (Fig. 3a). In the case of male, CS content in stage II is significantly (p = 0.01) increased from stage I and decreased therafter (Fig. 3b). The PL levels in the muscle of female were (p = 0.008) increased from stage I to IV (Fig. 3c). The content of PL was higher in females than males at stage IV. The highest contents of PL in male muscle tissue was observed at stage III and it was decreased (p = 3.65E-05) beyond stage IV (Fig. 3d). Content of TAG in muscle of females significantly (p = 4.29E-06) increased from stage I to stage IV and decreased (p = 4.05E-06) upto stage VI (Fig. 2e). In male, the content of TAG significantly (p = 4.05E-06) increased from stage I to III and decreased thereafter (Fig. 3f).
Lipid class content in liver: In female liver tissue, content of CS significantly (p = 4.50E-06) increased from stage I to III and significantly (p = 4.3E-06) decreased thereafter (Fig. 4a). Although the increase in CS content in male was moderately increased (p = 4.05E-06) from stage I to IV, it sharply decreased from stage IV to VI (Fig. 4b). PL content in female showed a slight fluctuation among maturity stages (Fig. 4c), significant (p = 0.09) difference was not observed from stage I to IV but a considerable (p = 0.01) drop was recorded at stage V. However, PL content in male increased from stage I to II and significantly (p = 4.05E-06) decreased from stage II to stage VI (Fig. 4d). Changes in the content of TAG was higher when compared to the moderate changes in PL and CS in the liver tissues of female (Fig. 4e). The mean TAG content of female liver tissues showed a 2 fold increase (p = 1.09E-05) from stage II to IV and then a 4 fold decrease (p = 4.29E-06) from stage IV to VI. A similar trend was also observed for TAG in male liver tissues (Fig. 4f) .
Monthly lipid changes in tissues: All three constituents of lipid class, CS, PL and TAG, showed higher and lower values corresponding to the maturation stage and annual spawning events (Table 2, 3). CS content of testis was significantly (p = 0.003) different from CS of ovary, whereas PL and TAG were not significantly different between male and female. Mean CS, PL and TAG content in ovary of females fluctuated throughout study and attained higher value in June and September (Table 2). Male gonads also followed a more or less similar pattern for CS, PL and TAG as that of female adult fish (Table 3).
The highest mean CS content in ovary of females collected in June and September months were 226.4 mg.100 g-1±78.21 and 292.00 mg.100 g-1±156.08, respectively (Table 2). Similar trend of CS content in male testis was observed (Table 3). CS content of testis significantly (p = 0.046) increased from July to September and attained a peak at September (226.3 mg.100 g-1±69.92). The mean PL content in ovary of female reached the highest amount in June as 393.54 mg 100 g-1±73.00 (Table 2).
 | |
| Fig. 3(a-f): | Box and Whisker plot showing changes of lipid class content (mg.100g-1) in muscle tissue of S. lysan in different gonad maturity stages, I: Immature unsex; II: Immature; III: Maturing; IV: Mature; V: Spawning; VI: Spent. a, c and e-in female; b, d and f-in Male. Mean values for each gonad maturity stages with the common letters indicate not significantly difference (p>0.05) |
Similarly, mean PL content in testis of male significantly (p = 0.040) increased from April to June, attained the highest amount as 475.8 mg.100g-1±153.6 and decreased in July (Table 3). Significantly (p = 0.034) highest TAG content in ovary of female was recorded in September (Table 2). The highest amount of TAG in testis of adult male fish was recorded in June as 453.1 mg.100g-1±77.64 (Table 3).
Muscle tissue of both sexes contained low values of CS than the PL and TAG in all months (Table 2, 3). Fluctuations of CS content in muscle tissues of female were significantly lower in June (p = 3.21E -05) and September (p = 1.8E-05) when compared to May.
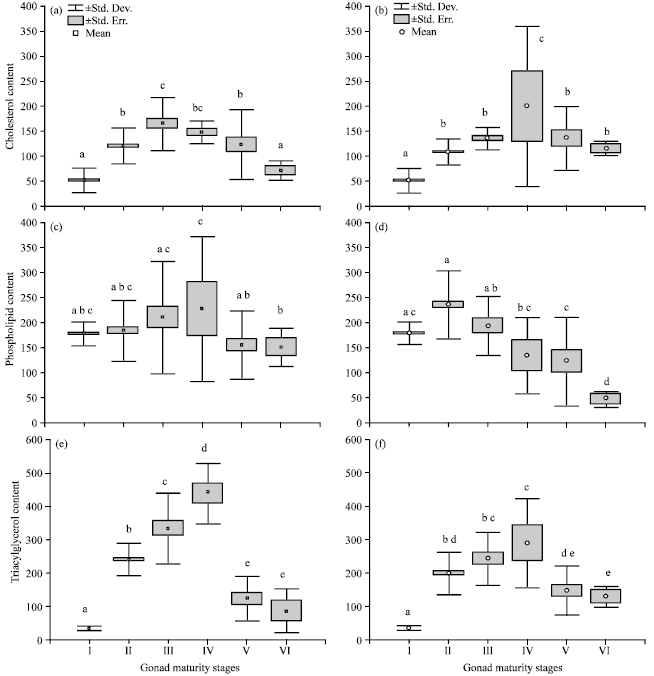 | |
| Fig. 4(a-f): | Box and Whisker plot showing changes of lipid class content (mg.100g-1) in liver tissue of S. lysan in different gonad maturity stages, I: Immature unsex; II: Immature; III: Maturing; IV: Mature; V: Spawning; VI: Spent, a, c and e -in female; b, d and f -in Male. Mean values for each gonad maturity stages with the common letters indicate not significantly difference (p>0.05) |
PL content in female muscle tissue significantly (p = 0.004) decreased from April to June. TAG content in muscle tissues of female was significantly (p = 0.001) decreased from August to September (Table 2). But, monthly changes of CS, PL and TAG in male muscle tissue was not significantly fluctuated (Table 3).
Changes in liver CS and PL in both sexes was not significantly predictable (Table 2, 3). But Fluctuation of TAG content in liver was evident in both sexes. Female liver TAG significantly (p = 0.0017) declined from May to June and significantly (p = 0.0012) increased from June to August and again significantly (p = 0.0002) decreased in September.
| Table 2: | Lipid class content of gonad, muscle and liver tissues in adult female S. lysan throughout the year 2010/2011 |
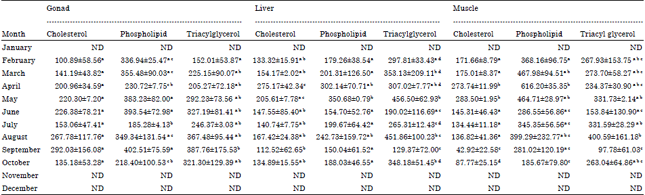 | |
| Mean values for each month with the common letters indicate not significantly difference (p>0.05). ND: Not detected, Values are Means±SD | |
| Table 3: | Lipid class content of Gonad, Muscle and Liver tissues in adult male S. lysan throughout the year 2010/2011 |
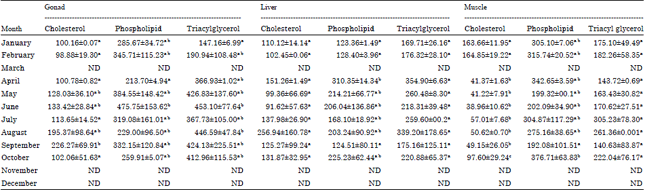 | |
| Mean values for each month with the common letters indicate not significantly difference (p>0.05). ND: Not detected, Values are Means±SD | |
Liver lipid in both sexes attained maximum value in August, whereas minimum values were obtained in June and September.
DISCUSSION
Analysis of lipid classes in gonad, muscle and liver tissues are a widely applied methodology in the study of reproduction (Shearer and Swanson, 2000; Das and Sahu, 2001).The results of the present study suggest that the S. lysan showed an important relationship between lipid classes and gonad maturity stages as well as with different months. It further shows that S. lysan undergoes major changes in lipid contents in gonad, muscle and liver tissue.
Lipid changes in tissues with gonad maturity stages: The findings of lipid classes in immature stages of S. lysan are in agreement with the observations made by Litvin et al. (2011), who reported that PL was the predominant lipid class in muscle of juvenile weakfish Cynoscion regalis, while TAG was in low concentrations. Yet, liver of juvenile fish contained low amount of TAG than that of muscle.
Total lipid content in ovary of S. lysan was higher in spawning stage while lowest in spent stage. Similar observation was shown in Trachinotus ovatus, where total lipid content of ovaries attained the highest value at spawning stage and lowest value at spent stage (Assem et al., 2005).
The results of the present investigation reveal that the mean value of lipid class composition in ovary has relatively higher amount of TAG and PL than the CS. Ovary of red drum (Vetter et al., 1983) and gilthead sea bream (Mourente and Odriozola, 1990) also contain highest amount of TAG and PL than the CS (proportions of total lipid content). Hilton et al. (2008) also noticed that the PL in the brood stock egg of yellowtail kingfish (Seriola lalandi) was higher whereas the triacylglycerol value was lower in egg of yellowtail kingfish. In contrast, sand eel has higher value of TAG than PL (proportions of total lipid content) (Tocher and Sargent, 1984).
Muscle lipid content of S. lysan attained a maximum value during mature stages and minimum value during spawning stage. Bransden et al. (2007) also identified a similar pattern, they specified that the fat content in muscle of male and female striped trumpeter Latris lineate were decreased by 25 and 40%, respectively during the spawning period. Contents of PL in muscle of S. lysan increased from immature stage to mature and decreased thereafter. Similar trend was demonstrated by Rao (1965), who recorded that concentration of inorganic phosphorous compounds in muscles of Caranx sexfasciatus increased with maturity. Yagana (1982) also reported that the value of phospholipid in muscle of catfish Clarias bairachus declined during spawning period and the low phosphorous content was observed in post-spawning period. In contrast, Thakur et al. (2009) reported that the polar lipid in muscle of yellowtail (Seriola quinqueradiata) was minor constituents throughout the maturation. Muscle tissues of mature S. lysan fish shows low CS content in the present study. Further the present study describes that S. lysan can be included under ‘low fat fish’ category (Sutharshiny and Sivashanthini, 2011a, c) Hence, consumption of S. lysan fish posses no risk to human health.
TAG content in liver tissues of mature S. lysan exhibited higher values. Seiichi et al. (1993) also identified that the major lipid component of the liver in amberjack and striped jack were triglyceride. Content of TAG in liver tissues of S. lysan was decreased after spawning. The observation is consistent with the findings by Phleger (1971), who found that the total liver lipid content of pink salmon Onchorhychus gorbuscha decreases in the spent fish. Phleger (1971) further explained that the liver of fish do not to synthesize the triglyceride after spawning. The cholesterol content of the liver of S. lysan exhibited a low variation during the maturation cycle and highest value was recorded at mature stage, while the minimal value was recorded at spent stage. Findings of the present investigation on S. lysan is in agreement with the work of Idler and Bitners, 1960), who reported that the total cholesterol content in liver declined and deposited in ovary of migratory salmon, Oncorhynchus nerka, during the spawning phases. In contrast, Phleger (1987) identified that the CS content in liver of pink salmon (Onchorhychus gorbuscha) remain constant after spawning.
Lipid changes in tissues through out the year: From the present study, it is obvious that the lipid class constituents CS, PL and TAG of gonad, muscle and liver tissues of tropical S. lysan show a variation throughout the year, corresponding to the maturation stage and annual spawning events. Peak spawning of adult female S. lysan was reported in June and September months (Thulasitha and Sivashanthini, 2013).
The lipid class content in ovary of S. lysan fluctuated throughout the year and attained a noticeable peak value during the spawning period while, muscle and liver lipid content of S. lysan attained the lower amount. Arrington et al. (2006) also proposed similar pattern of seasonal changes in lipid content of muscle, liver and gonad of three neo tropical fish. Similarly, Bustamante (1989) recorded that the body fat accumulated before the spawning of bar jack (Caranx ruber) and decreased during the spawning period. At the same time, the lipid content in ovary increased during the spawning time.
Major fluctuations of PL and TAG content in muscle of S. lysan were noticed during the period of spawning. Likewise, Thakur et al. (2009) showed TAG content in muscle of amberjack (Seriola dumerili) varied considerably with season. Although, Polar Lipid (PL) content in muscle remained almost constant over the study period.
Fluctuation of liver TAG content was evident in both sexes of S. lysan. Similarly, liver lipid composition of red drum (Sciaenops ocellatus) varied throughout the year (Craig et al., 2000). The mean TAG content in liver tissues of both sexes of S. lysan was higher during the maturation and declined thereafter. This is in confirmation with the findings of Craig et al. (2000).
CONCLUSION
From the present study it has been concluded that the variation in cholesterol (CS), phospholipid (PL) and triacylglycerol (TAG) in gonad, muscle and liver tissues in different gonad maturity stages, confirm a strong link between lipid profile and reproductive strategies of tropical Scomberoides lysan. Knowledge of the lipid dynamics of S. lysan throughout the year assist to determine the non spawning period and therefore it is the fishing season of S. lysan. Range of muscle CS, PL and TAG value obtained in the present study signifies that S. lysan recommended as one of the healthiest food fish for human consumption. Determination of CS, PL and TAG content in ovary, muscle and liver tissues of different maturity stages of S. lysan further provide information on nutrition of lipid in terms of diet formulation in future culture trails of S. lysan. The present study provides fundamental information to successful formulation and implementation of policies, strategies and plans in fisheries management and future aquaculture trials.
ACKNOWLEDGMENTS
The authors are grateful to the authorities of National Research Council, Sri Lanka for the financial assistance in the form of research grant (Grant No: 07-19) and university of Jaffna, Sri Lanka for the facilities provided. We also express our deep sense of gratitude for two anonymous reviewers for their valuable comments to improve the manuscript.
REFERENCES
- Alonso-Fernandez, A. and F. Saborido-Rey, 2012. Relationship between energy allocation and reproductive strategy in Trisopterus luscus. J. Exp. Mar. Biol. Ecol., 416-417: 8-16.
CrossRef - Anas, M.U.M., E.M.R.K.B. Edirisinghe and J.M.P.K. Jayasinghe, 2009. Lipid composition and fatty acid profiles of wild caught and fattened mud crab, Scylla serrata, in Sri Lanka. Sri Lanka J. Aquat. Sci., 14: 75-85.
CrossRef - Arrington, D.A., B.K. Daviavidson, K.O. Winemiller and C.A. Layman, 2006. Influence of life history and seasonal hydrology on lipid storage in three neotropical fish species. J. Fish Biol., 68: 1347-1361.
CrossRefDirect Link - Goda, A.M.A.S., O.M. El-Husseiny, G.M. Abdul-Aziz, A. Suloma and Y.H. Ogata, 2007. Fatty acid and free amino acid composition of muscles and gonads from wild and captive tilapia Oreochromis niloticus (L.) (Teleostei: Perciformes): An approach to development broodstock diets. J. Fish. Aquatic Sci., 2: 86-99.
CrossRefDirect Link - Assem, S.S., S.S. El-Serafy, M.M. El-Garabawy, M.E.G. Elabsawy and S.K. Kaldus, 2005. Some biochemical aspects of reproduction in female Trachinotus ovatus (Carangidae). Egypt. J. Aquat. Res., 31: 315-327.
Direct Link - Bligh, E.G. and W.J. Dyer, 1959. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol., 37: 911-917.
CrossRefPubMedDirect Link - Bransden, M.P., S.C. Battaglene, R.M. Goldsmid, G.A. Dunstan and P.D. Nichols, 2007. Broodstock condition, egg morphology and lipid content and composition during the spawning season of captive striped trumpeter, Latris lineate. Aquaculture, 268: 2-12.
Direct Link - Clarke, A., N. Doherty, A.L. DeVries and J.T. Eastman, 1984. Lipid content and composition of Three species of Antarctic Fish in Relation to buoyancy. Polar. Biol., 3: 77-83.
CrossRefDirect Link - Craig, S.R., D.S. MacKenzie, G. Jones and D.M. Gatlin, 2000. Seasonal changes in the reproductive condition and body composition of free-ranging red drum, Sciaenops ocellatus. Aquaculture, 190: 89-102.
CrossRef - Das, S. and B.K. Sahu, 2001. Biochemical composition and calorific content of fishes and shellfishes from Rushikulya Estuary, South Orissa coast of India. Indian J. Fish., 48: 297-302.
Direct Link - Ebrahimnezhadarabi, M., C.R. Saad, S.A. Harmin, M.K. Abdul Satar and A.A. Kenari, 2011. Effects of phospholipids in diet on growth of sturgeon fish (Huso-huso) juveniles. J. Fish. Aquat. Sci., 6: 247-255.
CrossRefDirect Link - Fiorin, R., S. Malavasi, A. Franco and P. Franzoi, 2007. Comparative energy allocation in two sympatric, closely related gobies: The black goby Gobius niger and the grass goby Zosterisessor ophiocephalus. J. Fish Biol., 70: 483-496.
CrossRefDirect Link - Foster, L.B. and R.T. Dunn, 1973. Stable reagents for determination of serum triglycerides by a colorimetric Hantzsch condensation method. Clin. Chim. Acta, 19: 338-340.
PubMedDirect Link - Hedayatifard, M. and M. Yousefian, 2010. The fatty acid composition of golden mullet fillet Liza aurata As affected by dry-salting. J. Fish. Aquat. Sci., 5: 208-215.
CrossRef - Hilton, Z., C.W. Poortenaar and M.A. Sewell, 2008. Lipid and protein utilisation during early development of yellowtail kingfish (Seriola lalandi). Mar, Biol., 154: 855-865.
Direct Link - Hiroaki, S., 2012. Lipid characteristics of two subtropical Seriola fishes, Seriola dumerili and Seriola rivoliana, with differences between cultured and wild varieties. Food. Chem., 135: 1718-1729.
PubMed - Huynh, M.D., D.D. Kitts, C. Hu and A.W. Trites, 2007. Comparison of fatty acid profiles of spawning and non-spawning Pacific herring, Clupea harengus pallasi. Comp. Biochem. Physiol. B., 146: 504-511.
CrossRefDirect Link - Jacques, R. and K. Patrick, 2003. Liver changes related to oocyte growth in roach, a single spawner fish and in bleak and white bream, two multiple spawner fish. Int. Rev. Hydrobiol., 88: 68-76.
CrossRef - Jezierska, B., J.R. Hazel and S.D. Gerking, 1982. Lipid mobilization during starvation in the rainbow trout. Salmo gairdneri Ricardson, with attention to fatty acids. J. Fish Biol., 21: 681-692.
CrossRef - Johnson, R.B., 2009. Lipid deposition in oocytes of teleost fish during secondary oocyte growth. Res. Fish. Sci., 17: 78-89.
CrossRef - Kandemir, S., 2010. The fatty acid composition and cholesterol and vitamin contents of different muscles of Esox lucius (Linnaeus, 1758) living in Lake Ladik. J. Anim. Vet. Adv., 9: 1179-1190.
CrossRefDirect Link - Ovie, K.S., A. Kabir and O. Jennifer, 2007. Sublethal effects of paraquat on some plasma organic constituents (metabolic parameters) of African Catfish: Clarias gariepinus (Osteichthys-Clariidae). Int. J. Zool. Res., 3: 213-217.
CrossRefDirect Link - Litvin, S.Y., V.G. Guida and M.P. Weinstein, 2011. Lipid class dynamics and storage depots in juvenile weakfish Cynoscion regalis and their application to condition assessment. J. Fish. Biol., 78: 1294-1310.
CrossRef - Lloret, J., M. Demestre and J. Sanchez-Pardo, 2008. Lipid (energy) reserves of European hake (Merluccius merluccius) in the North-Western Mediterranean. Vie. Et. Milieu., 58: 75-85.
Direct Link - Lund, E.D., C.V. Sullivan and A.R. Place, 2000. Annual cycle of plasma lipids in captive reared striped bass: Effects of environmental conditions and reproductive cycle. Fish. Physiol. Biochem., 22: 263-275.
CrossRef - Mourente, G. and J.M. Odriozola, 1990. Effects of broodstock diets on lipid classes and their fatty acid composition in eggs of gilthead sea bream (Sparus aurata L.). Fish. Physiol. Biochem., 8: 93-101.
Direct Link - Mourente, G., C. Megina and E. Diaz-Salvago, 2002. Lipids in female northern bluefin tuna (Thunnus thynnus thynnus L.) during sexual maturation. Fish. Physiol. Biochem., 24: 351-363.
Direct Link - Phleger, C.F., 1971. Liver triglyceride synthesis failure in post-spawning salmon. Lipids, 6: 347-349.
CrossRef - Phleger, C.F., 1987. Bone lipids of tropical reef fishes. Comp. Biochem. Physiol., B86: 509-512.
Direct Link - Qasim, S.Z., 1957. The biology of Blennius pholis L.(Teleostei). Proc. Zool. Soc. Lond., 128: 161-208.
CrossRefDirect Link - Rao, B.K.S.P., 1965. Biochemical studies on red and white muscles of Caranx sexfasciatus quoy and gaimard, I. Acid-soluble and acid-insoluble phosphorus compounds. Proc. Plant. Sci., 62: 87-91.
Direct Link - Talat, R., R. Azmat, S. Talata and Y. Akhter, 2006. Nutritive evaluation of edible trash fish. III: medicinal and commercial use of lipids of trash fish. Int. J. Zool. Res., 2: 123-128.
CrossRefDirect Link - Seiichi, A., Y. Mori, K. Nakamura and A. Sugawara, 1993. Characteristics of lipid accumulation types in five species of fish. Nippon. Suisan. Gakk., 59: 1559-1564.
Direct Link - Sharpe, R.L., M. Drolet and D.L. MacLatchy, 2006. Investigation of de novo cholesterol synthetic capacity in the gonads of gold fish (Carassius auratus) exposed to the phytosterol beta-sitosterol. Reprod. Biol. Endocrin.
Direct Link - Shearer, K.D. and P. Swanson, 2000. The effect of whole body lipid on early sexual maturation of 1+ age male chinook salmon (Oncorhynchus tshawytsch). Aquaculture, 190: 343-367.
CrossRef - Sheridan, M.A., 1994. Regulation of lipid metabolism in poikilothermic vertebrates. Comp. Biochem. Physiol. Part B, 107: 495-508.
Direct Link - Singh, R., A.K. Singh and M. Tripathi, 2012. Melatonin induced changes in specific growth rate, gonadal maturity, lipid and protein production in Nile Tilapia Oreochromis niloticus (Linnaeus 1758). Asian-Austral. J. Anim. Sci., 25: 37-43.
CrossRefDirect Link - Sutharshiny, S. and K. Sivashanthini, 2011. Lipid reserves of Scomberoides lysan (Pisces: Carangidae) from the Sri Lankan waters. Int. J. Biol. Chem., 5: 170-183.
CrossRefDirect Link - Sutharshiny, S. and K. Sivashanthini, 2011. Proximate composition of three species of Scomberoides fish from Sri Lankan waters. Asian J. Clin. Nutr., 3: 103-111.
CrossRefDirect Link - Sutharshiny, S. and K. Sivashanthini, 2011. Total lipid and cholesterol content in the flesh of the five important commercial fishes from waters around Jaffna Peninsula, Sri Lanka. Int. J. Biol. Chem., 6: 161-169.
CrossRefDirect Link - Taylor, M.H., 1984. Lunar synchronization of fish reproduction. Trans. Am. Fish. Soc., 113: 484-493.
CrossRef - Thakur, D.P., K. Morioka, N. Itoh, M. Wada and Y. Itoh, 2009. Muscle biochemical constituents of cultured amberjack Seriola dumerili and their influence on raw meat texture. Fish. Sci., 75: 1489-1498.
Direct Link - Thilakarathne, L.A.D.S. and M.V.E. Attygalle, 2009. Lipid composition of skin and muscle of the Indo-Pacific sailfish, Istiophorus platypterus. Vidyodaya J. Sci., 14: 161-166.
Direct Link - Thulasitha, W.S. and K. Sivashanthini, 2013. Microscopic staging system used in the identification of gonad developmental stages of Scomberoides lysan. J. Fish. Aqua. Sci., 8: 355-366.
CrossRef - Tocher, D.R. and J.R. Sargent, 1984. Analysis of lipids and fatty acids in ripe roes of some northwest European marine fish. LIPIDS., 19: 492-499.
CrossRefDirect Link - Tocher, D.R., 2003. Metabolism and functions of lipid and fatty acids in teleost fish. Revs. Fish. Sci., 11: 107-184.
CrossRef - Ubhayasekera, S.J.K.A., P. Jayasinghe, S. Ekanayake and P.C. Dutta, 2012. High cholesterol oxidation in pickled mackerel (Rastrelliger kanagurta) from Sri Lanka. Eur. J. Lipid. Sci. Tech., 114: 695-700.
CrossRefDirect Link - Varghese, M., M.K. Manisseri, N. Ramamurthy, P.M. Geetha, V.J. Thomas and A. Gandhi, 2011. Coral reef fishes of Gulf of Mannar, S.E of India. Fishing Chimes CMFRI., 31: 38-40.
Direct Link - Vetter, R.D., R.E. Hodson and C. Arnold, 1983. Energy metabolism in a rapidly developing marine fish egg, the red drum (Sciaenops ocellata). Can. J. Fish. Aquat. Sci., 40: 627-634.
CrossRefDirect Link - Yagana, B., 1982. Seasonal variations in the phosphorus contents of the muscle of catfish Clarias bairachus L. Proc. Indian Acad. Sci. Anim. Sci., 91: 423-426.
CrossRef - Zaboukas, N., H. Miliou, P. Megalofonou and M. Moraitou-Apostolopoulou, 2006. Biochemical composition of the Atlantic bonito Sarda sarda from the Aegean Sea (eastern Mediterranean Sea) in different stages of sexual maturity. J. Fish Biol., 69: 347-362.
CrossRef - Zilversmit, B.B. and A.K. Davis, 1950. Microdetermination of plasma phospholipids by trichloroacetic acid precipitation. J. Lab. Clin. Med., 35: 155-160.
PubMed - Zlatkis, A., B. Zak and A.J. Boyle, 1953. A new method for the direct determination of serum cholesterol. J. Lab. Clin. Med., 41: 486-492.
PubMedDirect Link - Fricke, H. and R. Kaese, 1995. Tracking of artificially matured eels (Anguilla anguilla) in the Sargasso Sea and the problem of the eel's spawning site. Naturwisswnschaften, 82: 32-36.
CrossRefDirect Link - Idler, D.R. and I. Bitners, 1960. Biochemical studies on sockeye salmon during spawning migration: IX. Fat, protein and water in the major internal organs and cholesterol in the liver and gonads of the standard fish. J. Fish. Res. Board. Can., 17: 113-122.
CrossRef