
Jiapeng Lin
The Key Lab of Animal Biotechnology of Xinjiang, Urumqi, People�s Republic of China
Liqin Wang
The Key Lab of Animal Biotechnology of Xinjiang, Urumqi, People�s Republic of China
Yuncheng Zhao
The Key Lab of Animal Biotechnology of Xinjiang, Urumqi, People�s Republic of China
Min Hou
The Key Lab of Animal Biotechnology of Xinjiang, Urumqi, People�s Republic of China
Jing Wang
The Key Lab of Animal Biotechnology of Xinjiang, Urumqi, People�s Republic of China
Juncheng Huang
The Key Lab of Animal Biotechnology of Xinjiang, Urumqi, People�s Republic of China
Asian Journal of Animal and Veterinary Advances
Year: 2012 | Volume: 7 | Issue: 9 | Page No.: 868-875







ABSTRACT
Oocyte quality directly affects production efficiency of modern assisted reproductive technologies such as the in vitro maturation, fertilization, cultivation of cattle oocytes and sperm injection, embryo transfer and animal cloning. Thus, maternal genes (GDF9, Zar1, Mater and DNMT1) are selected to analyze the relationship between mRNA expressions of these genes and quality of ovine oocyte. This study collects ovine oocytes at GV stage and 12-30 h of in vitro maturation (IVM) respectively, as well as ovine oocytes with the first polar body emitted and not emitted. With the application of quantitative RT-PCR technique, mRNA expression levels of GDF9, Zar1, Mater and DNMT1 of oocytes in the above test groups are tested. It is found that: mRNA relative expression levels of GDF9, Zar1 and Mater at GV stage are the highest (p<0.05), while expression of DNMT1 reaches the highest level at 12 h (p<0.05). With the maturity of oocytes, all gene expression levels gradually decline significantly between 18 and 24 h (p<0.05), while the expression levels of four genes present an upward trend at 30 h (p<0.05). Expression of 4 genes in non-polar body oocytes are all higher than those in polar body oocytes (p<0.05). In the natural bare oocytes, expression of each gene is lower than cumulus-oocytes complex. Thus, the test indicates that GDF9, Zar1, Mater and DNMT1 can be the molecular indicators to determine the quality of ovine oocytes.
PDF Abstract XML References Citation
Received: December 29, 2011;
Accepted: February 16, 2012;
Published: May 10, 2012
How to cite this article
Jiapeng Lin, Liqin Wang, Yuncheng Zhao, Min Hou, Jing Wang and Juncheng Huang, 2012. Relations Between GDF9, Zar1, Mater and DNMT1 Gene Relative mRNA Abundance and Oocytes Quality of Ovine Oocytes. Asian Journal of Animal and Veterinary Advances, 7: 868-875.
DOI: 10.3923/ajava.2012.868.875
URL: https://scialert.net/abstract/?doi=ajava.2012.868.875
DOI: 10.3923/ajava.2012.868.875
URL: https://scialert.net/abstract/?doi=ajava.2012.868.875
INTRODUCTION
Oocyte developmental competence is influenced by many factors, such as seasons (De Rensis and Scaramuzzi, 2003), follicle size (Saito et al., 2001) and health of donors and donor age (Marchal et al., 2002). Oocyte developmental competence can directly reflect the intrinsic quality of oocytes (Hendriksen et al., 2000), thus deciding whether the in vitro embryo production will be a success. A host of experimental data demonstrates that the in vitro culture conditions impose impact on the early embryonic developmental potential to a certain extent, yet the oocyte quality is the key factor that decides the developmental efficiency of oocytes blastocyst (Dieleman et al., 2002). In addition, in the fertilization process, the sperm only contributes DNA and a small amount of cytoplasm. Thus, for fertilization through intracytoplasmic sperm injection and nuclear transfer, to select oocyte with good quality will be a prerequisite of deriving high quality embryos. Moreover, the maturation of oocytes is one of the rate-limiting steps of in vitro embryo production (Downs and Polo, 1997).
Oocyte maturation is essential for the fertilization and early embryonic development. For years, researchers have been trying to find markers that can serve as effective indicators for mature oocytes and have found a number of morphological indicators relevant to the oocyte developmental competence: the germinal vesicle breakdown and emission of first polar body indicates the nuclear maturation. Besides, the greater the oocyte gap is, the severer the cytoplasm will contract, leading to insufficient oocyte quality. As the spindle is very sensitive to the changes of the surrounding environment and the cytoplasm, spindle rotation is conducive to the emission of polar body (Zhu et al., 2003; Tang et al., 2011). Moreover, a fully functional spindle can ensure accurate separation of chromosomes as the feature of mature oocytes at MII stage (Miyara et al., 2003; Al-Amin et al., 2007). In addition, with the detection of maturation efficiency of oocyte and developmental efficiency of blastocyst, maturation quality of oocyte can be reflected (Rahman, 2010).
After the fertilization, the speed of early embryo cell division and the location of all blastomeres are controlled by mRNA and protein which is stored in the maternal oocytes (Telford et al., 1990). The fertilization media is collected by maternal mRNA in oocytes. After the fertilization, the protein synthesis is increased rapidly and the stored mRNA is transcribed and translated. The developmental competence of these genes include regulation of transcription and translation, regulation of the cell cycle, stress responses, histone synthesis, transfer signals of plasmodesma, growth factor and intracellular signaling and metabolism and so on (Wrenzycki et al., 2005). A host of experimental data demonstrates that the in vitro culture conditions impose impact on the early embryonic developmental potential to a certain extent, yet the oocyte quality is the key factor that decides the developmental efficiency of oocytes blastocyst (Zhu et al., 2003). Yet, evaluation reports of oocyte quality are relatively less from the molecular perspective.
Oocytes are divided into three groups: Different in vitro maturation time, the emission of first polar body 24 h after the in vitro maturation, no emission of first polar body, natural naked oocytes and COCs at GV stage. With the application of quantitative RT-PCR technique, mRNA expression levels of GDF9, Zar1, Mater and DNMT1 of ovine oocytes in the above test groups are tested which provides experimental basis and the foundation for the research of oocytes quality.
MATERIALS AND METHODS
Chemicals: Where not specified, the products were purchased from Sigma Chemical Co., St. Louis, Mo.
Oocyte collection: Ovaries were collected from ovine ovaries slaughtered at a local slaughterhouse and transported to the laboratory within 2 to 3 h in 0.9% NaCI solution at 28 to 30°C. Cumulus-oocyte complexes (COCs) were aspirated from follicles 2 to 6 mm in diameter with a 20 g needle fitted to a 5 mL syringe. After aspiration, COCs were washed in HEPES-buffered tissue culture medium 199 with 0.1% (w/v) polyvinyl alcohol.
Eighty oocytes of all experimental group were collected after blown repeatedly so as to remove cumulus cells as many as possible, centrifuged, lysed with RLT buffer (RNeasy Lyser Tissue, Qiagen, Europe) and stored at -70°C.
| Table 1: | The sequence of primers for quantitative Real-time PCR |
 | |
In vitro maturation: After washing, the COCs selected were placed in a dish containing maturation media for 20-22 h at 39°C in a humidified atmosphere of 5% CO2 in air. The maturation medium was composed of: TCM 199 supplemented with 10% heat treated oestrus ovine serum, 10 mL mL-1 of FSH/LH and 100 mM cysteamine (Rahim and Saki, 2010).
RNA extraction and RT-PCR: Total RNA was isolated using an Rneasy Micro Kit (Qiagen, Europe) according to the manufacturer’s instructions.
RT-PCR condition: To mix 1 μg total RNA and 1 μL random six polymer primer, RNase free water is make up to 14.5 μL at 70°C for 10 min and experiences ice bath for 10 min. Upon the transient centrifugation, 0.5 μL RNA Inhibiter, 5 x buffer 2 μL, 10 mmol L-1 dNTP 2 μL and AMV 1 μL (TaKaRa) are added in for 42°C for 40 min.
Standard curve: Mater, Zar1, DNMT 1 and GDF9 genes are respectively connected with 18S rRNA and pMD20-T carrier. The restricted enzyme EcoR V is applied to linearize the plasmids of each gene and linearized plasmid of each gene is undertaking dilute concentration so as to make standard curve.
According to the gene sequence in GeneBank, Oligo 6.0 software is used to design quantitative primers of ovine Mater, Zar1, DNMT 1 and GDF9 gene and the internal reference gene 18S-rRNA (Table 1).
Quantitative RT-PCR: Quantitative RT-PCR (Roche, 1.5 LightCycle) reaction condition: for 2 μL standard plasmid and the sample cDNA, each 5 pM upstream and downstream primers are used, also including 2 x FastStart DNA Master SYBR Green I 10 μL (QIAGEN), plus RNase free water to 20 μL.
Reaction conditions: 95°C pre-denaturation for 10 min; 55 cycles include 95°C 10 see, 60°C 20 see and 72°C 20 see. From 40~99°C, the temperature for the melting curve is increased by 2°C per second and fluorescence signals are collected. In order to reduce the errors, the quantitative RT-PCR is repeated once again. The average value is applied in the analysis. All samples were operated in duplicate and the mean value of each duplicate was used for all further calculations.
Statistical analysis: Development data were analysed by ANOVA using the PROC GLM of the software Statistical Analysis System, version 9.1.3 and the Tukey test was used when statistical differences were detected. Data in percentage were arcsine transformed prior to ANOVA.
RESULTS
Expressions of genes in different maturation time: The 18S rRNA is taken as an internal reference gene. With standard curve method, specific primers are applied to cDNA samples for qRT-PCR which can test changes of gene expression of ovine oocytes at GV stage and different IVM culture times (Fig. 1).
From the results, it can be found that at the GV stage, the four genes show certain accumulation tendency. In relatively different maturation time, relatively higher gene expression (p<0.05) is shown. With the extension of IVM time, all the genes have shown a downward trend in mRNA expression which reaches the bottom at 18 and 24 h, respectively. It bounces significantly at 30 h (p<0.05) (Fig. 2).
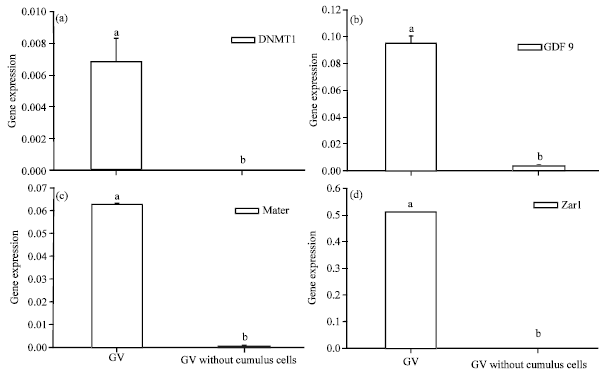
Expression of gene mRNA in naked oocytes, COCs and oocytes with or without the first polar body (Fig. 3).
Eighty pieces of ovine oocytes with the first polar body emitted or not emitted at COCs of GV stage, naked oocytes and IVM stage are collected respectively. The expression variation conditions of four genes are analyzed.
The results show that expressions of all the genes with the polar body emitted are significantly higher than those with polar body not emitted (p<0.05), while oocytes expression at GV stage is significantly higher than the natural naked oocytes (p<0.05).
 | |
| Fig. 1: | Indicates mRNA genes expression, namely, (A) DNMT1,(B) GDF9, (C) Mater and (D) Zar1 in ovine oocytes in GV stage and different IVM periods detected by quantitative RT-PCR. Differences of a, b indicate p<0.0, GV: Oocytes at GV stage, 12 h, IVM 12 h, 18 h IVM 18 h, 24 h IVM 24 h, 30 h IVM 30 h |
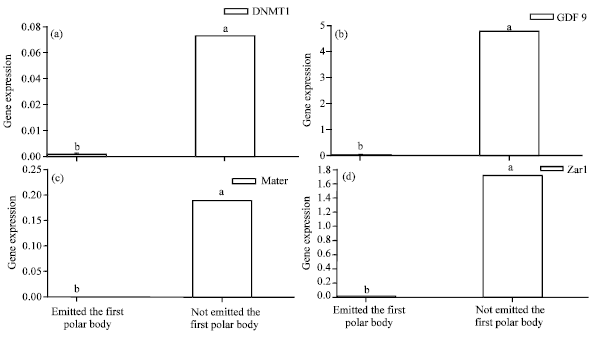 | |
| Fig. 2: | Indicates mRNA gene expression of (A) DNMT1,(B) (C) GDF9, Mater and (D) Zar1 in ovine oocytes with or without the first polar body which are detected by quantitative RT-PCR. Differences in a and b indicate p<0.05 |
 | |
| Fig. 3: | Indicates mRNA gene expression of (A) DNMT1,(B) GDF9, (C) Mater and (D) Zar1 in COCs at GV stage and naked oocytes at GV stage which are detected by quantitative RT-PCR. Differences in a and b indicate p<0.05, GV: COCs at GV stage, GV without cumulus cells: Natural naked oocytes at GV stage |
DISCUSSION
Maturation of the oocyte cytoplasm not only experiences ultrastructural changes of organelle, but also undergoes a series of complex molecular physiological and biochemical processes (De Sousa et al., 1998). A great number of genes are active in the transcription and the persistent protein translation has accumulated many transcript and micro-elements that support subtrahend division and embryonic development after fertilization (De Sousa et al., 1998). In the growth and maturation process, the mammalian oocyte has prepared and accumulated a set of maternal RNA and proteins which are needed in the embryonic development and constitute the maternal materials for the early embryonic development. Among them, the frequently studied genes include GDF9, Zar1, Mater and DNMT1 and so on (Tong et al., 2000; Howell et al., 2001; Prolla et al., 1998; Christians et al., 2000).
The experiment results indicate that mRNA expression of GDF9, Zar1, Mater, DNMT1 genes is highest in oocytes at GV stage which can be illustrated that at GV stage, the cytoplasm has accumulated sufficient maternal RNA and proteins needed in embryonic development. This has established certain material foundation for IVM. Along the development of oocyte maturation process, its mRNA expression declines. Thus, it can be inferred that these mRNA have been depleted in the IVM of oocytes to join the in vitro maturation process of oocytes.
With the process of meiosis, germinal vesicle breakdown occurs and oocytes emit the first polar body at the animal pole, indicating the nuclear maturation. Secondary oocyte will stay at the second meiotic metaphase (MII) before ovulation, waiting for the fertilization (Rahman et al., 2008). It is generally proved that oocyte maturation is determined by whether the vesicle breakdown occurs and the first polar body is emitted. This is the major morphological identification method of current oocyte quality. Yet, in the process of the in vitro maturation, nuclear and cytoplasmic maturation of oocytes are often not synchronized. The cytoplasmic maturation not only embodies changes in ultrastructure of organelle but also experiences a series of physiological and biochemical changes in the cytoplasm. Thus, its maturation identification cannot be observed solely from the morphological shape. So GDF9, DNMT1, Zar1 and Mater gene mRNA expression with or without the first polar body are detected in this paper. Results show that the above gene expression levels with are polar body significantly lower than that without polar body (p<0.05) and it can be inferred that this is because these genes cannot be normally transcribed in oocytes with polar body emitted, leading to the extensive accumulation. This generates its disability of gaining necessary proteins for the later development so that it is ultimately degraded or died. This may be one of the reasons that oocytes fail or delay the emission of the first polar body and indicate that GDF9, DNMT1, Zar1 and Mater genes may be related with the discharge condition of first polar body.
Oocytes and the surrounding cumulus cells achieve the signaling through paracrine and gap connections (Sutton et al., 2003) and the oocyte growth in ovarian follicles is regulated by the proteins generated by cumulus cells (Sterneck et al., 1997) which fully demonstrates the importance of secretion effects of cumulus cells to the development of oocytes. Thus, COCs at normal GV stage and natural naked oocytes are respectively collected for the diction of the expression of the above four genes. It is found that the expression of each gene in GV stage is significantly higher than that in natural naked oocytes (p<0.05). The result demonstrates that content of maternal RNA in natural naked oocytes is relatively smaller and may not provide adequate maternal RNA and proteins to maintain post-embryonic development and a series of physiological and biochemical changes (Ozluk, 2010). Thus, it is inferred that this could be one of the important reasons that affect maturation efficiency, post in vitro fertilization merogenesis efficiency and blastocyst efficiency of similar oocytes without the surrounding of granular cells.
Morphology regards oocytes without the first polar body as nuclear maturation which can provide better quality than that with first polar body. Based on this, it is assumed that the two hold differences on the molecular aspect. Thus, expression of maternal gene of the above two kinds of oocytes is detected. When the IVM is between 22-24 h, that is, oocytes rest at MII stage, mRNA expression of each gene in oocytes without the first polar body is lower than that with the first polar body(p<0.05). Thus, it is inferred that in the IVM developmental process of oocytes, the maternal RNA and proteins accumulated in GV stage are persistently consumed. In other words, these maternal genes or/and proteins promote or drive the developmental process from GV stage to M stage of oocytes which is possibly the preparation of secondary oocyte for the fertilization. Moreover, the consistent expression tendency of genes in oocyte without or with polar body further supports the inference.
To sum up, quantitative RT-PCR method is adopted, maternal mRNA expressions of genes in ovine oocytes at different IVM times, with the first polar body emitted or not emitted and with or without cumulus cells are studied. It is convinced that four genes, GDF9, DNMT1, Mater and Zar1, are eligible to serve as the molecular indicators of evaluation the quality of ovine oocyte.
ACKNOWLEDGMENTS
This study was supported by National transgenic major special of China (No. 2011ZX08008-003), the Hi-Tech Research and Development Program of China’s Xinjiang Uygur Autonomous Region (No. 200711104) and Development Program of China’s Xinjiang Uygur Autonomous Region (No. 201111113).
REFERENCES
- Rahman, A.N.M.A., 2010. Intracytoplasmic sperm injection-revolution in human and animal assisted reproduction: A review. Biotechnology, 9: 392-410.
CrossRef - Rahman, A.N.M.A., R.B. Abdullah and W.E. Wan-Khadijah, 2008. Gametogenesis, fertilization and early embryogenesis in mammals with special reference to goat: A review. J. Boil. Sci., 8: 1115-1128.
CrossRefDirect Link - Ozluk, A., 2010. The variation of the amount of the neurosecretory materials in the corpus allatum of Pimpla turionellae L. (hymenoptera: Ichneumonidae) during oocyte maturation. Pak. J. Biol. Sci., 13: 748-752.
CrossRef - Christians, E., A.A. Davis, S.D. Thomas and I.J. Benjamin, 2000. Maternal effect of Hsf1 on reproductive success. Nature, 407: 693-694.
CrossRef - Tang, D.W., S. Liu, Z.X. Liu, Y. Wu and X.L. Wang et al., 2011. Relationships of Ca2+ oscillations and ER cluster distribution to porcine oocyte fragmentation. Asian J. Anim. Vet. Adv., 6: 322-332.
CrossRefDirect Link - De Sousa, P.A., A. Caveney, M.E. Westhusin and A.J. Watson, 1998. Temporal patterns of embryonic gene expression and their dependence on oogenic factors. Theriogenol., 49: 115-128.
PubMed - Miyara, F., F.X. Aubriot, A. Glissant, C. Nathan and S. Douard et al., 2003. Multiparameter analysis of human oocytes at metaphaseⅡstage after IVF failure in non-male infertility. Hum Reprod, 18: 1449-1503.
CrossRef - Rahim, F. and G. Saki, 2010. Use of open pulled straw as a carrier in vitrification of metaphase ii oocyte in mice. Asian J. Biol. Sciences, 3: 85-91.
CrossRefDirect Link - Dieleman, S.J., P.J.M. Hendriksen, D. Viuff, P.D. Thomsen and P. Hyttel et al., 2002. Effects of in vivo prematuration and in vivo final maturation on developmental capacity and quality of pre-implantation embryos. Theriogenology, 57: 5-20.
CrossRefDirect Link - Howell, C.Y., T.H. Bestor, F. Ding, K.E. Latham, C. Mertineit, J.M. Trasler, J.R. Chaillet, 2001. Genomic imprinting disrupted by a maternal-effect mutation in the DNMT1 gene. Cell, 104: 829-838.
CrossRef - Marchal, R., C. Vigneron, C. Perreau, A. Bali-Papp and P. Mermillod, 2002. Effect of follicular size on meiotic and developmental competence of porcine oocytes. Theriogenology, 57: 1523-1532.
Direct Link - Al-Amin, M., A. Nahar and M.S. Ali, 2007. Luteinizing hormone (LH) effects on In vitro nuclear maturation of bovine oocytes. Asian J. Cell Biol., 2: 50-53.
CrossRefDirect Link - Prolla, T.A., S.M. Baker, A.C. Harris, J.L. Tsao and X. Yao et al., 1998. Tumour susceptibility and spontaneous mutation in mice deficient in Mlh1, Pms1 and Pms2 DNA mismatch repair. Nat. Genet, 18: 276-279.
CrossRef - De Rensis, F. and R.J. Scaramuzzi, 2003. Heat stress and seasonal effects on reproduction in the dairy cow-A review. Theriogenology, 60: 1139-1151.
CrossRefDirect Link - Sterneck, E., L. Tessarollo and P.F. Johnson, 1997. An essential role for C/EBPβ in female reproduction. Genes Dev., 11: 2153-2162.
CrossRef - Sutton, M.L., R.B. Gilchrist and J.G. Thompson, 2003. Effects of in vivo and in vitro environments on the metabolism of the cumulus-oocyte complex and its influence on the oocyte developmental competence. Hum. Reprod. Update, 9: 35-48.
Direct Link - Telford, N.A., A.J. Watson and G.A. Schultz, 1990. Transition from maternal to embryonic control in early mammalian development: A comparison of several species. Mol. Reprod. Dev., 26: 90-100.
CrossRef - Tong, Z.B., L. Gold, K.E. Pfeifer, H. Dorward and E. Lee et al., 2000. Mater, a maternal effect gene required for early embryonic development in mice. Nat. Genet, 26: 267-268.
PubMed - Wrenzycki, C., D. Herrmann, A. Lucas-Hahn, K. Korsawe, E. Lemme and H. Niemann, 2005. Messenger RNA expression patterns in bovine embryos derived from in vitro procedures and their implications for development. Reprod Fertil Dev, 17: 23-35.
PubMed - Zhu, Z.Y., D.Y. Chen, J.S. Li, L. Lian, L. Lei, Z.M. Han and Q.Y. Sun, 2003. Rotation of meitotic spindle is controlled by microfilaments in mouse oocytes. Biol. Reprod, 68: 943-946.
PubMed