
Hai Jin
School of Animal Science and Technology, Anhui Agricultural University, Hefei, 230036, The People`s Republic of China
Hong-Quan Chen
School of Animal Science and Technology, Anhui Agricultural University, Hefei, 230036, The People`s Republic of China
Jie Qin
School of Animal Science and Technology, Anhui Agricultural University, Hefei, 230036, The People`s Republic of China
Yin-Jian Zhu
School of Animal Science and Technology, Anhui Agricultural University, Hefei, 230036, The People`s Republic of China
Hua Chen
School of Animal Science and Technology, Anhui Agricultural University, Hefei, 230036, The People`s Republic of China
Gong-Wei Chen
School of Animal Science and Technology, Anhui Agricultural University, Hefei, 230036, The People`s Republic of China
Ya-Nan Xie
School of Animal Science and Technology, Anhui Agricultural University, Hefei, 230036, The People`s Republic of China
Zhong-Ting Pan
School of Animal Science and Technology, Anhui Agricultural University, Hefei, 230036, The People`s Republic of China
Ming-Hui Jiao
School of Animal Science and Technology, Anhui Agricultural University, Hefei, 230036, The People`s Republic of China
Sheng-Qiang Huang
School of Animal Science and Technology, Hunan Agricultural University, Changsha, 410128, The People`s Republic of China
Ming-Xing Chu
Key Laboratory of Farm Animal Genetic Resources and Utilization of Ministry of Agriculture, Institute of Animal Science, Chinese Academy of Agricultural Sciences, Beijing 100193, The People`s Republic of China
Asian Journal of Animal and Veterinary Advances
Year: 2012 | Volume: 7 | Issue: 7 | Page No.: 568-577







ABSTRACT
Adenosine Monophosphate activated Protein Kinase (AMPK) plays a key role in cellular metabolism and regulating energy and its γ3 subunit was encoded by the PRKAG3 gene which was regarded as the candidate gene for carcass quality trait in domestic animals and fowls. To understand PRKAG3 gene variation and its association with carcass quality of goat breeds, the polymorphism of goat PRKAG3 gene and its distribution patterns in different goat breeds were investigated. Five goat breeds selected were Anhui white goat, Matou, Boer, Xiangdong and Sanen dairy goat. DNA samples of all goat breeds were extracted from goat ear tissue. The polymorphisms in the 5’ regulatory region and exon 13 of PRKAG3 gene were examined using ApaI, Hpy188I, AciI and ClaI enzyme digestion, respectively and their distribution patterns were analyzed in different goat breeds. The results showed that two mutation loci in the 5’ regulatory region, C-525A and C-225T, located at -525 and -225 bp upstream of the start codon of the PRKAG3 gene and two mutation loci in the exon 13, T90C and C102T, located at 90 bp and 102 bp of the exon 13. The 525CC genotype frequencies of five goat breeds listed above were 0.8000, 0.9681, 0.6042, 0.7557 and 0.8889, respectively, while 225CC genotype frequencies corresponding to 0.7391, 0.9750, 0.8684, 0.9785 and 0.9344. The mutations at T90C and C102T didn’t cause the substitution of corresponding amino acids in the AMPK protein. The muscle fat grade and carcass fat grade are lower in goat breeds with higher 525CC225CC genotype frequency while higher in the breeds with higher 525CA225CT genotype frequency (p<0.05). The results suggest that the lipoidosis ability of goat breeds may be associated with C-525A and C-225T loci of PRKAG3 gene.
PDF Abstract XML References Citation
Received: October 26, 2011;
Accepted: December 26, 2011;
Published: February 25, 2012
How to cite this article
Hai Jin, Hong-Quan Chen, Jie Qin, Yin-Jian Zhu, Hua Chen, Gong-Wei Chen, Ya-Nan Xie, Zhong-Ting Pan, Ming-Hui Jiao, Sheng-Qiang Huang and Ming-Xing Chu, 2012. The Polymorphism in 5 'Regulatory Region and Exon 13 of PRKAG3 Gene and its Distribution Pattern in Different Goat Breeds. Asian Journal of Animal and Veterinary Advances, 7: 568-577.
DOI: 10.3923/ajava.2012.568.577
URL: https://scialert.net/abstract/?doi=ajava.2012.568.577
DOI: 10.3923/ajava.2012.568.577
URL: https://scialert.net/abstract/?doi=ajava.2012.568.577
INTRODUCTION
China has many goat breeds, native goat breeds nearly 50 and its rearing number reached 152 million in 2010, which has been playing an important role in meat supply and agricultural improvement. Due to diverse natural and geographical conditions, cultivated goat breeds in the particular habitat differ in the goat meat quality, commercial grade, meat taste, flavor and juiciness for consumer (Arain et al., 2010a, b; Das et al., 2007). Some reports showed that meat quality related to muscle fat content (Parizadian et al., 2011). However, in addition to the effect of nutrition level (Vahdatpour et al., 2008), the fat deposition ability of goat depends on the characters of goat breeds (Zhou et al., 2011) and associated with many fat metabolic enzymes. The molecular mechanism of fat deposition ability in goat yet remains unclear, although many molecular markers correlated with carcass fat content were found of which very few are utilized in breeding practice (Mahmoudi, 2010; Qiong et al., 2011; Yadav and Yadav, 2007). Compared with studies in the pig, cattle and other animal (Chen, 2010; Han et al., 2011; Chen et al., 2011a), there are few studies on the fat deposition ability in goat.
AMPK is highly conserved sensor of cellular energy and it plays a key role in cellular metabolism and regulating energy (Carling, 2004). Activation and phosphorylation of AMPK can promote glucose uptake, enhance fatty acid oxidation and inhibition of glycogen synthesis and consumption of energy in the body (Witters et al., 2006). AMPK is a heterotrimeric enzyme complex, consisting of one catalytic subunit (α) and two regulation subunits (β and γ) (Stapleton et al., 1997; Shen et al., 2006). γ subunit is divided into three isoforms that are γ1, γ2 and γ3 subunits encoded, respectively by the PRKAG1, PRKAG2 and PRKAG3 gene (Hardie, 2004; Long and Zierath, 2006).
Human PRKAG3 gene contains 13 exons and 12 introns (Park et al., 2003). PRKAG3 is specifically expressed in skeletal muscle (Yu et al., 2003). Milan et al. (2000) and Ciobanu et al. (2001) found that Arg200/Gln200 mutation of PRKAG3 protein in Hampshire pig reduced AMPK activity and increased skeletal muscle glycogen content, while the Val199/Ile199 mutations decreased skeletal muscle glycogen content (Mallick et al., 2006), which thereby influenced the meat quality. PRKAG3 gene mutations in human and mouse resulted in an increase of lipid peroxidation which affected the metabolism of skeletal muscle (Barnes and Zierath, 2005). Some reports found that the polymorphisms of cattle PRKAG3 gene related to the protein activity, meat quality traits and disease resistance (Roux et al., 2006; Yu et al., 2005). At present, the polymorphism of goat PRKAG3 gene and its association with meat quality has not been reported. In this study, the polymorphisms in 5’ regulatory region and exon 13 of goat PRKAG3 gene was detected and the polymorphic distribution in five goat breeds and its association with meat quality were analyzed.
MATERIALS AND METHODS
Samples: Genomic DNA samples were extracted form the ear tissues of five goat breeds using DNA extraction kit, stored at 4°C. The five goat breeds were Sanen dairy goat (Sanen) from Northwest Sci-Tech University of Agriculture and Forestry (Shanxi, China), Xiangdong from Xiangdong black goat sheepstud of Changsha (Hunan, China), Matou from Matou goat breed conservation field of Shimen County (Hunan, China) and Anhui white goat (Anbai) and Boer from Boda Feidong sheepstud (Anhui, China).
Assessment of goat meat grade: The Muscle Fat Grade (MFG) of five goat breeds were estimated according to the fat of goat longissimus muscle and less than 1.0% in the muscle as level 1, then each additional 0.5% to improve a grade, respectively. The Carcass Fat Grade (CFG) were assessed based on the fat content of goat carcass and less than 10.0% in the carcass as level 1, then each additional 1.0% to improve a grade, respectively.
Primers: Based on the sequences of bovine PRKAG3 gene (GenBank DQ082736), four pairs of primers were designed using the primer premier 5.0 program (http://www.premierbiosoft.com) in which primer P5-900 and PE-13 were employed for the amplification of the fragment containing 5’ partial regulatory region and exon 13 of goat PRKAG3 gene and primer P5-600 and P5-350 for PCR-RFLP analysis of the 5’ regulatory region. The primers were synthesized by Shanghai Bioengineering Inc. (Shanghai, China).
| P5-900: | Upstream primer 5’GTTGGCTCTAGGAGACGCAG3’ Downstream primer 5’ ATACACAGAGAAAATCGTGGTCA3’ |
| P5-600: | Upstream primer 5’ CGTCAAGGTGATTCTCAGGACT3’ Downstream primer 5’CGAGTGCGCAACACTGTATCT3’ |
| P5-350: | Upstream primer 5’TTTCCTCCTTTGGCACCTGAC3’ Downstream primer 5’CGAGTGCGGCACACTGTATCT3’ |
| PE-13: | Upstream primer 5’ ATTCTTAGTATCAACCTCATCAGC3’ Downstream primer 5’GAGCCTACCTGAACAAGAGC3’ |
PCR conditions and procedures: PCR was carried out using 25 μL PCR amplification kits: 1.0 μL (10 μM) of each primer, 2.5 μL of 10xPCR buffer with Mg2+, 2.0 μL dNTP (2.5 mM), 0.5 μL (5 units μL-1) Taq DNA polymerase (Beijing Trans Gen Biotech Co., Ltd. China), 1.0 μL genomic DNA (50 ng μL-1) and 17 μL double-distilled H2O. Amplification conditions were: initial denaturation of 5 min at 94°C; 35 cycles of 94°C for 30 sec, 30 sec at 57°C for primer P5-900 (35 sec at 59°C for primer P5-600, 58°C for primer P5-350 and 55°C for primer PE-13, respectively), 72°C for 1 min, followed by 72°C extension for 10 min. The PCR products were detected using 1.5% agarose gel electrophoresis.
Polymorphism analysis: PCR products digested with ApaI (Fermentas China Co., Ltd.) and Hpy188I (New England BioLabs), ClaI (Fermentas China Co., Ltd.) and AciI (Fermentas China Co., Ltd.) endonucleases and then electrophoresed on 3% agarose gel for genotyping polymorphisms. The gene frequencies of the different goat breeds were analyzed statistically. Allele gene frequencies (Chen et al., 2011b), p and q, was calculated by p = D+0.5H and q = 1-p, respectively where D and H represent homozygote and heterozygote genotype frequencies, respectively. The gene frequencies of two linkage SNP loci were calculated by Zhou et al. (2011).
Statistical analysis: The 5’ regulatory region was subjected to promoter and transcription factors binding loci prediction in Signal scan (http://www-bimas.cit.nih.gov/molbio/signal/). The correlation coefficients between the genotypes and the levels of MFG and CFG were calculated and evaluated for p = 0.05 significant relationship using student’s t-test.
RESULTS
Sequence analysis of the promoter region and exon 13: PCR amplification fragment lengths of primer P5-900, P5-600, P5-350 and PE-13 were 889, 596, 351 and 314 bp in Fig. 1, respectively. Based on the sequencing results of mixed DNA samples (Fig. 2), two mutation sites, C-525A and C-225T located at -525 and -225 bp upstream of the start codon of the PRKAG3 gene, respectively, were detected. The promoter region of PRKAG3 gene was assessed for potentially functional sequence elements by comparison with paralogous promoter sequences.
 | |
| Fig. 1: | Sequence characters of promoter region and exon 13 of goat PRKAG3 gene. The primers with same names in the brackets are a pair of primers and the region between upstream and downstream primers is the DNA sequence. The capital letters are the sequences of exon 1 and exon 13 of PRKAG3 gene, respectlvely. The atTgg sequence was binding motif for NF-Y transcription factors and the ccCctggg sequence for AP-2 transcription factors, in which capital letters, T and C, are mutation sites. While replacing the nucleotide at both sites, T→C and C→A, the binding sites for NF-Y and AP-2 disappeared |
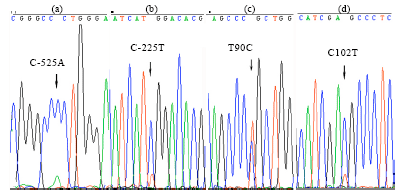 | |
| Fig. 2: | The mutation sites in the promoter region and exon 13 of goat PRKAG3 gene. C-525A and C-225T locate at -525 and -225 bp upstream of the start codon of the PRKAG3 gene, respectively. T90C and C102T locate at 90 bp and 102 bp in the exon13 of the PRKAG3 gene, respectively |
ATTGG sequence at C-225T was the binding motif of NF-Y transcription factor and CCCCTGGG sequence at C-525A was that of AP-2 transcription factor, while the binding site disappeared after T→C and C→A at C-225T and C-525A, respectively.
In exon 13, T90C and C102T mutant sites located at 90 bp and 102 bp of the exon 13 of the PRKAG3 gene were detected, respectively, which were only synonymous mutations for the protein.
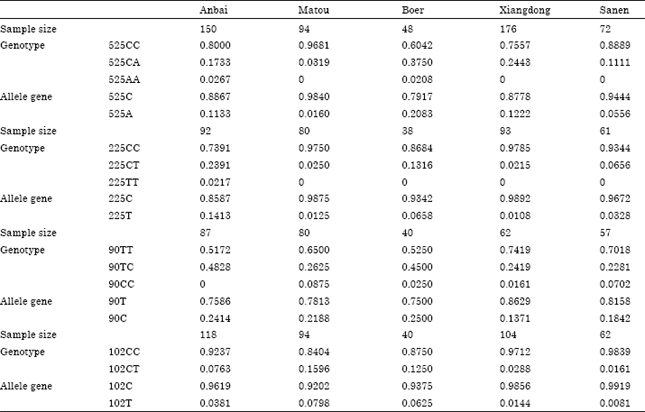
| Table 1: | The genotype and allele frequencies of PRKAG3 gene at C-525A, C-225T, T90C and C102T sites in different goat populations |
 | |
Polymorphisms of the promoter region and exon 13 in different goat breeds: Incision of the PCR product by ApaI, Hpy188I, AciI and ClaI, respectively, revealed that four polymorphism sites consisted of three genotypes, repectively.
Genotype and allele frequencies of the four sites in different goat populations were shown in Table 1. 525C gene frequencies of the Matou, Sanen, Xiangdong, Anbei and Boer were 0.9840, 0.9444, 0.8778, 0.8867 and 0.7917, respectively. 525AA genotype was not found in the Matou, Sanen and Xiangdong. From Table 1, the frequencies of 225CC genotype were higher than 0.93 in the Matou, Sanen and Xiangdong. 225TT genotype was only detected in the Anbai and lower. 90T was advantage alleles in the five goat populations at T90C site. The frequency of 90TT genotype in Xiangdong was higher (0.7419) and that in Boer lower (0.5250). 102C was advantage allele at C102T site. The frequencies of allele 102C in all goat populations were higher than 0.92. All goat populations of the four sites were in Hardy-weinberg equilibrium (p>0.05).
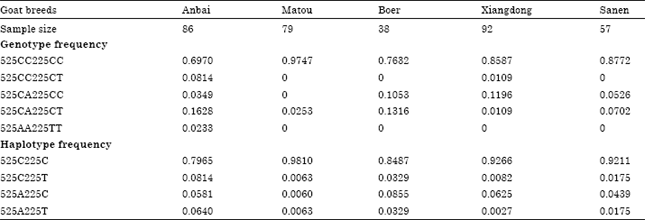
The genotype and haplotype frequencies of C-525A and C-225T linkage sites were shown in Table 2. The 525CC225TT, 525CA225TT, 525AA225CC and 525AA225CT were not detected. The 525CC225CC was preponderant genotype and the frequency was higher than 0.69 for all goat breeds, next the 525CA225CT genotype for Anbei (0.1628) and Boer (0.1316) and followed by the 525CA225CC genotype for Boer (0.1053) and Xiangdong (0.1196). The haplotype frequency of 525C225C was higher than 0.79.
Association of the polymorphisms with goat lipoidosis ability: The correlation coefficients between goat lipoidosis ability and genotype frequencies were shown in Table 3. The MFG and CFG were lower in goat breeds with higher 525CC225CC genotype frequency, whlie higher in the breeds with higher 525CA225CT genotype frequency (p<0.05).
| Table 2: | The genotype and haplotype frequencies of C-525A and C-225T linkage of goat PRKAG3 gene |
 | |
| Table 3: | The association of the genotypes with goat lipoidosis ability |
 | |
| *p<0.05 | |
The heterozygous at C-525A and C-225T resulted in the increase of goat lipoidosis ability.
DISCUSSION
There have been some reports about the polymorphisms of PRKAG3 gene and their genetic effects on meat quality in ruminant animals. Lu (2009) found 9 polymorphic loci of the PRKAG3 gene in 7 sheep breeds, the distribution of which differed. Some polymorphic sites have been detected in the 5’ regulatory region and exon 3, 9, 10 and 11 in the foreign and local chicken breeds (Zhao et al., 2006). Many mutation sites were found in the exons of pig PRKAG3 gene (Ciobanu et al., 2001; Lindahl et al., 2004). In the present study, we have also found 4 polymorphic loci in the 5’ regulatory region and exon 13 of PRKAG3 gene in native goat breeds of China. These results showed that the polymorphism of PRKAG3 gene was rich in many animals.
AMPK acts as a metabolic master switch regulating several intracellular systems including the cellular uptake of glucose, the β-oxidation of fatty acids and the biogenesis of glucose transporter 4 and mitochondria (Thomson et al., 2007). Li et al. (2009) found that the polymorphism of PRKAG3 gene and its association with beef meat tenderness, fleshcolor and some amino acids. The T2885C of two polymorphic sites in intron 4 of cattle PRKAG3 gene significantly related to meat tenderness. In addition, the mutation of cattle PRKAG3 gene resulted in change of bovine muscle glycogen content and pH value (Immonen et al., 2000). Lu (2009) confirmed that the meat color scores and pH values among genotypes of the sheep PRKAG3 gene significantly differ. In the chicken PRKAG3 genes, the 1744 genotype significantly affected on the water content and crude protein content of chicken meat (p<0.01) and rate of the chest (p<0.05) (Zhou, 2008; Yin et al., 2011). The mutation of pig PRKAG3 gene was significantly associted with carcass quality (Lindahl et al., 2004). Amino acid substitutions at some important places of PRKAG3 protein, I199V, G52S and T30N, might improve the quality of pork and reduce muscle glycogen content (Ciobanu et al., 2001). Obviously, the polymorphism of PRKAG3 gene causes the diversity of AMPK function, which results in change of the animal meat quality. Present investigation indicated that the heterozygous at C-525A and C-225T in the 5’ regulatory region of goat PRKAG3 gene resulted in the increase of goat lipoidosis ability (p<0.05), while the two mutational sites in exon 13 showed the codon non-random usage (Zhang et al., 2007) which prefers to uese the CCU for Pro and GAC for Asp.
AP-2 transcription factor binds to GC-rich sequences (Bosher et al., 1996). The cis-acting DNA sequences 5’-(G/C)CCCA(G/C)(G/C)(G/C)-3’ and the palindromic sequences 5’-GCCNNNGGC-3’ are considered as consensus AP-2 binding sites (Eckert et al., 2005; Debieve et al., 2011). The AP-2 is highly involved in the development and function of the central nervous system. Thus, AP-2 has been identified as an important regulator of gene expression during embryonic development of many neural tissues including the midbrain, hindbrain, spinal cord, cranial and dorsal root ganglia (Pfisterer et al., 2002). The CCAAT box is one of the most common elements in the promoters of candidate genes with important functions in animal metabolism (Hirwa et al., 2011), found in the forward or reverse orientation (ATTGG). The frequency of CCAAT boxes appears to be relatively higher in TATA-less promoters, particularly in the reverse ATTGG orientation (Mantovani, 1998). NF-Y transcription factor binging to the box plays an important role in the regulation of coordinate and tissue-specific gene expression (Dolfini et al., 2012). Present investigation found two mutational sites involved in AP-2 binding site at C-525A and NF-Y binding site at C-225T. It is interesting that Matou, Xiangdong and Sanen have higher AP-2 frequency and lower NF-Y frequency in the binding site of transcription factor and lower levels in MFG and CFG. These imply that PRKAG3 gene influences goat lipoidosis ability via regulating the development and function of goat central nervous system and itself expression level.
CONCLUSION
According to the results of this study, C-525A and C-225T of goat PRKAG3 gene are two SNP loci affacting MFG and CFG levels of goats. The muscle fat content and carcass fat content are lower in goat breeds with higher 525CC225CC genotype frequency, whlie higher in the breeds with higher 525CA225CT genotype frequency. C-525A and C-225T loci may be associated with the lipoidosis ability of goat breeds.
ACKNOWLEDGMENTS
The study was supported by The Modern Technology System of National Sheep Industry (nycytx-39), The Modern Technology System of Sheep Industry of Anhui Province and New Varieties of GMO Cultivation of Major Projects of the Ministry of Agriculture (2009ZX08008-007B).
REFERENCES
- Arain, M.A., M. Khaskheli, I.R. Rajput, S. Faraz, S. Rao, M. Umer and K. Devrajani, 2010. Effect of slaughtering age on chemical composition of goat meat. Pak. J. Nutr., 9: 404-408.
CrossRefDirect Link - Arain, M.A., M. Khaskheli, I.R. Rajput, S. Rao and S. Faraz et al., 2010. Examination of physical properties of goat meat. Pak. J. Nutr., 9: 422-425.
CrossRefDirect Link - Das, A.K., R.B. Sharma and N.P. Singh, 2007. Quality and storage stability of low acid goat meat pickle. Am. J. Food Technol., 2: 550-554.
CrossRefDirect Link - Barnes, B.R. and J.R. Zierath, 2005. Role of AMP-activated protein kinase in the control of glucose homeostasis. Curr. Mol. Med., 5: 341-348.
PubMed - Bosher, J.M., N.F. Totty, J.J. Hsuan, T. Williams and H.C. Hurst, 1996. A family of AP-2 proteins regulates C-ERBB-2 expression in mammary carcinoma. Oncogene, 13: 1701-1707.
PubMed - Carling, D., 2004. The AMP-activated protein kinase cascade a unifying system for energy control. Trends Biochem. Sci., 29: 18-24.
CrossRef - Chen, H.Q., H.Q. Wei, J. Qin and H. Chen, 2011. The novel genetic change in 5'-untranslated region of goose prolactin gene and their distribution pattern in different goose breeds. Asian J. Anim. Vet. Adv., 6: 1069-1075.
CrossRefDirect Link - Chen, H.Q., Y.P. Zhang, H. Chen, J. Qin and L.S. Wang, 2011. Polymorphisms in the 5' regulatory region of the gpr54 gene and their association with sexual maturity in cattle. Mol. Reprod. Dev., 78: 894-894.
CrossRefPubMedDirect Link - Ciobanu, D., J. Bastiaansen, M. Malek, J. Helm, J. Woollard, G. Plastow and M. Rothschild, 2001. Evidence for new alleles in the protein kinase adenosine monophosphate-activated {gamma}3-subunit gene associated with low glycogen content in pig skeletal muscle and improved meat quality. Genetics, 159: 1151-1162.
PubMedDirect Link - Dolfini, D., R. Gatta and R. Mantovani, 2012. NF-Y and the transcriptional activation of CCAAT promoters. Crit. Rev. Biochem. Mol. Biol., 47: 29-49.
PubMed - Eckert, D., S. Buhl, S. Weber, R. Jager and H. Schorle, 2005. The AP-2 family of transcription factors. Genome Biol., 6: 246-246.
CrossRefDirect Link - Han, B., H.L. Zhang, L. Zeng, B. Yang, R. Abula, X.L. Xu and Y. Chen, 2011. The Polymorphisms of β2-Adrenergic Receptor Gene on two Cattle Breeds in China Asian J. Anim. Vet. Adv., 6: 715-722.
CrossRef - Hardie, D.G., 2004. AMP-activated protein kinase: a key system mediating metabolic responses to exercise. Med. Sci. Sports Exerc., 36: 28-34.
PubMed - Hirwa, C.A., P. Wallace, X. Shen, Q. Nie, G. Yang and X. Zhang, 2011. Genes related to economically important traits in beef cattle. Asian J. Animal Sci. 5: 34-45.
CrossRefDirect Link - Immonen, K., M. Ruusunen, K. Hissa and E. Puolanne, 2000. Bovine muscle glycogen concentration in relation to finishing diet, slaughter and ultimate pH. Meat Sci., 55: 25-31.
PubMed - Lindahl G., A.C. Enfalt, G. von Seth, A. Joseli and I. Hedebro-Velander et al., 2004. A second mutant allele (V199I) at the PRKAG3 (RN) locus-II. Effect on colour characteristics of pork loin. Meat Sci., 66: 621-627.
CrossRefPubMedDirect Link - Long, Y.C. and J.R. Zierath, 2006. AMP-activated protein kinase signaling in metabolic regulation. J. Clin. Invest., 116: 1776-1783.
Direct Link - Mahmoudi, B., 2010. Genetic diversity of lori goat population based on microsatellite marker. Asian J. Anim. Sci., 4: 13-19.
CrossRefDirect Link - Mallick, C., R. Maiti and D. Ghosh, 2006. Antidiabetogenic effects of separate and composite extract of seed of jamun (Eugenia jambolana) and root of kadali (Musa paradisiacal) in streptozotocin-induced diabetic male albino rat: A comparative study. Int. J. Pharmacol., 2: 492-503.
CrossRefDirect Link - Mantovani, R., 1998. A survey of 178 NF-Y binding CCAAT boxes. Nucleic Acids Res., 26: 1135-1143.
Direct Link - Milan, D., J.T. Jeon, C. Looft, V. Amarger and A. Robic et al., 2000. A mutation in PRKAG3 associated with excess glycogen content in pig skeletal muscle. Science, 288: 1248-1251.
CrossRefPubMedDirect Link - Parizadian, B., M.S. Shargh and S. Zerehdaran, 2011. Study the effects of different levels of energy and l-carnitine on meat quality and serum lipids of japanese quail. Asian J. Anim. Vet. Adv., 6: 944-952.
CrossRef - Park, H.B., S. Marklund, J.T. Jeon, J.R. Mickelson, S.J. Valberg, K. Sandberg and L. Andersson, 2003. Molecular characterization and mutational screening of the PRKAG3 genen in the horse. Cytogenet. Genome Res., 102: 211-216.
PubMedDirect Link - Pfisterer, P., J. Ehlermann, M. Hegen and H. Schorle, 2002. A subtractive gene expression screen suggests a role of transcription factor AP-2α in control of proliferation and differentiation. J. Biol. Chem., 277: 6637-6644.
CrossRef - Shen, Q.W., W.J. Means, S.A. Thompson, K.R. Underwood and M.J. Zhu et al., 2006. Pre-slaughter transport, AMP-activated protein kinase, glycolysis and quality of pork loin. Meat Sci., 74: 388-395.
CrossRef - Stapleton, D., E. Woollatt, K.I. Mitchelhill, J.K. Nicholl and C.S. Fernandez et al., 1997. AMP-activated protein kinase isoenzyme family: Subunit structure and chromosomal location. FEBS Lett., 409: 452-456.
CrossRefPubMedDirect Link - Thomson, D.M., B.B. Porter, J.H. Tall, H.J. Kim, J.R. Barrow and W.W. Winder, 2007. Skeletal muscle and heart LKB1 deficiency causes decreased voluntary running and reduced muscle mitochondrial marker enzyme expression in mice. Am. J. Physiol. Endocrinol. Metab., 292: E196-E202.
CrossRefPubMedDirect Link - Vahdatpour, T., K. Nazer-Adl, Y. Ebrahim-Nezhad, N. Maheri-Sis and S. Vahdatpour, 2008. The effects of energy increasing and protein lowering by addition of fats to diet on broiler chickens: performance and serum lipids. AJAVA., 3: 286-292.
CrossRefDirect Link - Qiong, W., F. Chao, L. Wu-Jun, F. Yi and Y. Shi-Gang, 2011. A novel mutation at exon 4 of IGF-1 gene in three indigenous goat breeds in china. Asian J. Anim. Vet. Adv., 6: 627-635.
CrossRefDirect Link - Witters, L.A., B.E. Kemp and A.R. Means, 2006. Chutes and Ladders: The search for protein kinases that act on AMPK. Trends Biochem. Sci., 31: 13-16.
CrossRefPubMedDirect Link - Yadav, A. and B.R. Yadav, 2007. Genetic diversity among six breeds of Indian goat using RAPD markers. Biotechnology, 6: 57-60.
CrossRefDirect Link - Yin, H.D., X.L. Zhao, Z.C. Zhang, N.N. Zhou and Q. Zhu, 2011. Association of Prkag3 gene polymorphisms with meat quality traits in chinese meat-type quality chicken populations. Adv. Mater. Res., 343-344: 631-636.
CrossRef - Yu, H., N. Fujii, M.F. Hirshman, J.M. Pomerleau and L.J. Goodyear, 2003. Cloning and characterization of mouse 5'-AMP-activated protein kinase γ3 subunit. Am. J. Physiol. Cell Physiol., 286: C283-C292.
PubMed - Zhang, T.Y., H.Q. Chen, Z. Liang, Y.H. Li and X.W. Gan, 2007. Cloning and sequence analysis of melanocortin receptor-4 gene in goose. Acta Vet. Zootech. Sin., 38: 913-919.
Direct Link - Zhao, C.J., C.F. Wang, X.M. Deng, Y. Gao and C.H. Wu, 2006. Identification of single-nucleotide polymorphisms in 5' end and exons of the PRKAG3 gene in Hubbard White broiler, Leghorn layer and three Chinese indigenous chicken breeds. J. Anim. Breed. Genet., 123: 349-352.
CrossRefPubMedDirect Link - Zhou, Q.Q., H.Q. Chen, H.Q. Wei, J. Qin, H. Chen and Y.P. Zhang, 2011. Association of polymorphism in the coding region of THRSP gene with lipogenesis capability in pigs. Prog. Biochem. Biophys., 38: 84-90.
CrossRefDirect Link