
Yanyan Ke
Laboratory of Veterinary Anatomy, College of Veterinary Medicine, China Agricultural University, Beijing 100193, China
Wenjie Liu
Collage of Pharmaceutical Sciences, Xiamen University, Xiamen 361005, China
Zixu Wang
Laboratory of Veterinary Anatomy, College of Veterinary Medicine, China Agricultural University, Beijing 100193, China
Yulan Dong
Laboratory of Veterinary Anatomy, College of Veterinary Medicine, China Agricultural University, Beijing 100193, China
Yaoxing Chen
Laboratory of Veterinary Anatomy, College of Veterinary Medicine, China Agricultural University, Beijing 100193, China
Asian Journal of Animal and Veterinary Advances
Year: 2011 | Volume: 6 | Issue: 9 | Page No.: 935-943







ABSTRACT
The purpose of this study was to determine the role of sympathetic nerves on distribution of the gut immune-associated cells (Including Intraepithelial Lymphocyte (IEL), mast cell, IgA+, CD4+ and CD8+ cell) in small intestine. A model of chemical sympathectomy in mice was established by intraperitoneal injection of 6-hydroxydopamine (6-OHDA). The gut immune-associated cells were investigated by histology and immunohistochemistry method. This study found that from proximal to distal segments in small intestine of normal mice, the numbers of intraepithelial lymphocyte, IgA+ and mast cells decreased gradually, whereas CD4+ and CD8+ cell number increased gradually. When 6-hydroxydopamine (6-OHDA) was intraperitoneally injected into mice, IEL numbers were significantly decreased in whole small intestines. However, IgA+ and mast cell numbers decreased merely in the proximal segment while CD4+ and CD8+ cell numbers and their ratio reduced mainly in the distal segment. These findings indicated that sympathetic denervation disturbed the patterns of gut immune-associated cell distribution. It would substantiate the thesis of neuro-immune-endocrine and provide the new ideas for the intestinal disease prevention and drug developments.
PDF Abstract XML References Citation
Received: May 30, 2011;
Accepted: June 28, 2011;
Published: August 08, 2011
How to cite this article
Yanyan Ke, Wenjie Liu, Zixu Wang, Yulan Dong and Yaoxing Chen, 2011. The Role of Sympathectomy on the Distribution of Intraepithelial Lymphocyte, Mast Cell, IgA+, CD4+ and CD8+ Cell in Intestine of Mice. Asian Journal of Animal and Veterinary Advances, 6: 935-943.
DOI: 10.3923/ajava.2011.935.943
URL: https://scialert.net/abstract/?doi=ajava.2011.935.943
DOI: 10.3923/ajava.2011.935.943
URL: https://scialert.net/abstract/?doi=ajava.2011.935.943
INTRODUCTION
The intestinal epithelium has the largest body surface area so the nutrients can be efficiently absorbed and the rapid and effective protective immune responses are also needed to exclude pathogens. A number of important chronic diseases of mucosal surfaces, such as Crohn’s disease and ulcerative colitis were related to the mucosal immunity disorder in the intestine (Deshmukh et al., 2010; Nikfar et al., 2011). Thus, further studies of the mechanisms underlying the modulation of intestinal mucosal immunity would be important for preventing and treating those diseases. On the other hand, the intestinal immunity is influenced by extrinsic and intrinsic factors. The extrinsic factors include nutrition (Ramiro-Puig et al., 2008; Hamidi et al., 2010), stress (Jarillo-Luna et al., 2008), microorganism (Nasrollah, 2009; Swinkels et al., 2006), vaccines (Bailey et al., 2007) and antimicrobial peptides (Yurong et al., 2006; Oda et al., 2010). The intrinsic factors include age (Gonzalez-Ariki and Husband, 2000), endocrine (Cima et al., 2004) and neurotransmitters (Levite and Chowers, 2001). Peptidergic neurotransmitters such as substance P and nerve growth factors have been the main focus of neuroimmunomodulation research in the gut (Straub et al., 2006), but the studies about the neurotransmitters such as catecholamine which are released by sympathetic nervous were scarce. In fact, sympathetic nerve fibers enter the intestinal wall along arteries and terminate in the myenteric and submucosal plexuses and the extrinsic noradrenergic nerve fibers provide 90% of the noradrenaline present in the gut (Nijhuis et al., 2010). Therefore, the Sympathetic Nervous System (SNS) plays an important role in modulating intestinal function.
The previous study demonstrated that the sympathetic nerves regulated the CD8+ cells and IL-2 production in the uterus during early pregnancy (Dong et al., 2007). It indicated that the SNS also play an important role in non-lymphoid organs. In fact, the gut has dense sympathetic innervations and 70-80% of the body’s immune cells (Straub et al., 2006) and the immune cells possess adrenergic receptors (Emeny et al., 2007). Thus the SNS may have the potential to modify intestinal immunity. 6-Hydroxydopamine (6-OHDA) is a neurotoxin that selectively destroys sympathetic nerve fibers with the catecholamine high affinity carrier which is found on nerve terminals in the peripheral nervous system. It had been reported that the chemical sympathectomy could change the Intraepithelial Lymphocycle (IEL) number in the duodenum of mice (Jarillo-Luna et al., 2008) and IgA+ cell number in the jejunum of rat (Gonzalez-Ariki and Husband, 1998, 2000). However, less is known about the regulation of the sympathetic nerve in distribution pattern of gut immune-associated cells in various segments of small intestine. The gut mucosal immune-associated cells are the first line of defense against food-derived and bacterial antigens penetrating to the mucosal epithelium (Bar-Shira et al., 2003). These cells usually include IEL, IgA+, Mast Cell (MC), CD4+ and CD8+ cells which are the chief components of intestinal immunological barrier. The purpose of this study was to determine the role of sympathetic nerves on distribution of the gut immune-associated cells Including IEL, MC, IgA+, CD4+ and CD8+ cell) from proximal to the distal small intestines.
MATERIALS AND METHODS
Animals and 6-OHDA treatment: This study was conducted from April to May 2010. A total of 59 male (35-40 g) Kun-ming albino mice of 7-week-old were housed under controlled temperature (24±1°C), humidity (40±5%) and lighting (12 h light, 12 h dark cycle) conditions and had free access to standard food and water. After an adaptive period of 1 week, the mice were randomly divided into 6-OHDA group (n = 30) and control group (n = 29). In 6-OHDA group, the mice were intraperitoneally injected for 5 consecutive days with 100 mg kg-1 Body Weight (BW) of 6-OHDA (Sigma) diluted in a sterile saline solution containing 0.01% L-ascorbic acid (Dong et al., 2007). The control mice were treated with vehicle alone (0.01 mL g-1 BW). This research was conducted in accordance with the Guidelines for Chinese National Animal Experimentation.
All the mice were killed by cervical dislocation under nembutal deep anaesthesia (50 mg g-1 BW) on the sixth day after 6-OHDA injection. The intestinal segments (1 cm) were removed from the duodenum, jejunum and ileum under sterile conditions.
Histology and immunohistochemistry: Intestinal tissues were immediately immersed in 4% paraformaldehyde in 0.1 M PB (pH 7.4) fixed overnight and dehydrated in graded ethanol series, then embedded in paraffin. Sections (5 μm) were mounted on gelatinized glass slides and were deparaffinized, rehydrated and rinsed with 0.01M PBS.
For histological studies, sections were stained with hematoxylin and eosin. Eight cross-sections for each intestinal segment were prepared for each mouse. Number of intestinal IEL was counted from 5 intact villus of each section and data were expressed as the average number per 100 Intestinal Epithelium Cells (IECs). The other eight cross-sections for each intestinal segment were stained for 15 min with toluidine blue solution (8 g L-1 toluidine blue and 6 mg L-1 KMnO4 dissolved in distilled H2O). Density of Mast Cell (MC) was calculated on each histological section.
For immunohistochemistry, sections were treated with 3% hydrogen peroxide in methanol for 30 min and incubated with 2.5% normal goat serum (Sigma) for 20 min at room temperature. After rinsing with PBS, sections were incubated overnight at 4°C with monoclonal rat anti-mouse CD4+ (1:200, Santa Cruz Biotechnology, Inc., CA), CD8+ (1:100, Thermo) or IgA+ (Beijing Biosynthesis Biotechnology Co., LTD) primary antibodies, respectively. Then the sections were rinsed in PBS and incubated with biotinylated goat anti-rat secondary antibody (1:200, Sigma) for 2 h at room temperature. After washing, the tissues were incubated with streptavidin-horseradish peroxidase (1:250, Sigma) for 1 h at 25°C. Immunoreactivity was visualized by incubating in 0.01 M PBS containing 0.05% 3'3-diaminobenzidine tetrahydrochloride (DAB, Sigma) and 0.003% hydrogen peroxide for 10 min in the dark. After the final rinsing, the sections were mounted. The specificity of the immunostaining was checked by omitting incubation in primary antibodies. Number of positive cells was counted in the total areas of three cross-sections of each animal and the areas of the cross-sections were measured by the Scan Image software. Densities of these cells were calculated.
Statistical analysis: Results were expressed as Mean±SD and statistical analyses were carried out with ANOVA or independent-samples t-test of SPSS 16.0. The p-value of less than 0.05 was considered statistically significant.
RESULTS
Changes on gut mucosal immune-associated cell number in mice treated with 6-OHDA: IEL is a special lymphocyte distributed in the enterocytes in villi. As seem from the picture, most of the IELs are located in the basal membrane of the enterocytes (Fig. 1a, c). As shown in Table 1, in the control group, the number of IEL generally decreased from proximal (duodenum) to distal (ileum). The cell number in duodenum (16.1±2.8 cell 100 IECs-1) was larger than that in ileum (9.2±1.8 cell 100 IECs-1) (p<0.01). After 6-OHDA treatment, IEL number was significantly decreased in the whole of small intestine (p<0.05).
Mast cells scattered around the intestinal gland. They were identified by metachromatic granules with toluidine blue staining and displayed round shapes, fusiform shapes or elongated shapes in the intestine (Fig. 1b, d). In the control group, the cell number in duodenum (207.6±36.0 cell cm-2) was 131.6% larger than in ileum (89.6±14.3 cell cm-2) (p<0.01). After 6-OHDA treatment, the cell numbers in the duodenum (126.8±14.5 cell cm-2) were remarkably lower than that of the control (207.6±36.0 cell cm-2) (p<0.01). But there were no significant difference in jejunum and ileum compared with the control (Table 1).
IgA+ cells predominated in the lamina propria. All positively labelled cells had characteristic plasma cell morphology (Fig. 2a, d). In control group, the numbers of IgA+ decreased gradually from proximal to distal intestines. The cell number in duodenum (307.2±35.4 cell mm-2) was larger than in ileum (235.9±25.0 cell mm-2) (p<0.01).
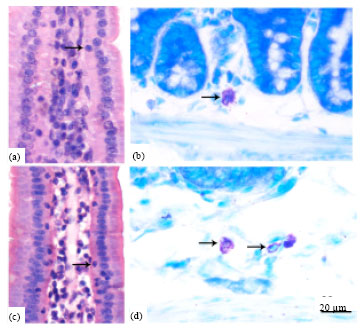 | |
| Fig. 1(a-d): | Photomicrographs represent the IEL (arrow) in the mice jejunum (a, c) and MC in the duodenum (b, d). The sections of a and c were stained with hematoxylin and eosin and the sections of B and D were stained with toluidine blue. Cell numbers of IEL and MC were significantly decreased in the 6-OHDA-treated group (a and b) than in the control group (c and d). Bar = 20 μm |
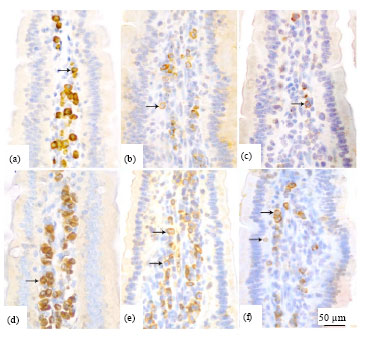 | |
| Fig. 2(a-f): | Photomicrographs depicting appearance of the IgA+ cells (a, d) in the mice duodenum and the CD4+ (b, e) and CD8+ (c, f) cells in jejunum. Note that the sections were stained with immunohistochemistry. Cell numbers of IgA+, CD4+ and CD8+ were significantly decreased in the 6-OHDA-treated group (a-c) than in the control group (d-f). Bar = 50 μm |
| Table 1: | The changes on the cell numbers of IEL, MC and IgA+ in each intestinal segment in mice treated with 6-OHDA (Mean±SD) |
 | |
| *, **Indicate significant differences (p<0.05; p<0.01 according to independent-samples t-test) between the control and 6-OHDA treated groups. The different letters within the same group (control group: A-C, 6-OHDA-treated group: a-c) are significantly different (p<0.05; LSD post hoc analysis conducted for each group after ANOVA) | |
| Table 2: | The changes on the cell numbers of CD4+, CD8+ and the ratio of CD4+/CD8+ in each intestinal segment in mice treated with 6-OHDA (Mean±SD) |
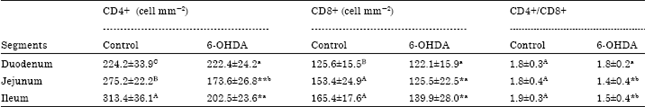 | |
| *, **Indicate significant differences (p<0.05; p<0.01 according to independent-samples t-test) between the control and 6-OHDA treated groups. The different letters within the same group (control group: A-C, 6-OHDA-treated group: a-c) are significantly different (p<0.05; LSD post hoc analysis conducted for each group after ANOVA) | |
After 6-OHDA treatment, the IgA+ cell numbers of duodenum (263.8±31.7 cell mm-2) and jejunum (240.1±27.3 cell mm-2) were remarkably lower than the duodenum (307.2±35.4 cell mm-2) and jejunum (269.8±23.7 cell mm-2) in control group (p<0.05) but no statistical differences with respect to the IgA+ number in ileum (Table 1).
The CD4+ (Fig. 2, e) and CD8+ (Fig. 2c, f) labelled cells were scattered in the lamina propria, some IEL were detected as the CD8+ positive cells. The CD4+ and CD8+ cell numbers increased gradually from proximal to the distal intestines in control group. There was no significant difference among three segments in the ratio of CD4+/CD8+ cells in control group (p>0.05). After 6-OHDA treatment, the CD4+ cell numbers of jejunum (173.6±26.8 cell mm-2) and ileum (202.5±23.6 cell mm-2) were remarkably lower than the jejunum (275.2±22.2 cell mm-2) and ileum (313.4±36.1 cell mm-2) in the control group (p<0.01). The reduction of CD8+ cell numbers was also found in jejunum and ileum (p<0.05) but no statistical difference was observed in duodenum. As a consequence, the ratios of CD4+/CD8+ cells of 6-OHDA treated group were significantly decreased in the jejunum (1.4±0.4) and ileum (1.5±0.4) than that of the control group (1.8±0.4 in jejunum, 1.9±0.3 in ileum) (p<0.05) (Table 2).
DISCUSSION
Present results showed that these gut mucosal immune-associated cells were distributed in the small intestine with a characteristic pattern. In this study, the numbers of IEL, IgA+ and MC decreased gradually from proximal to distal intestines, whereas the CD4+ and CD8+ cell number increased gradually. The similar findings were also reported in the Balb/c mice (Resendiz-Albor et al., 2010) and the rabbit (Liu et al., 2008). In contrast, some previous studies also found that the numbers of CD4+ and CD8+ cells were no difference among the intestinal segments in dog (Elwood et al., 1997) and the IgA+ cell number was the highest in the jejunum in cat (Waly et al., 2001). The different results between previous and present studies were probably caused by the difference between species. Nevertheless, these results suggested that the distribution of immune-associated cells showed a characteristic pattern in the small intestine. It was likely to be critical for maintaining constancy of normal mucosal environment.
Some previous studies found that the number of IgA+ cells was more in duodenum than in ileum. It could be due to a decreasing antigen load from duodenum to ileum as a consequence of degradation of ingesta (Kleinschmidt et al., 2008). The lack of epithelial cell-derived cytokines such as IL-7 and IL-15, significantly enhance IEL survival but T cell receptor stimulation and glucocorticoids exhibit antagonistic activity on IEL survival (Inagaki-Ohara et al., 1997; Brunner et al., 2001). In contrast, there were no significant changes in the number of whole IEL after repeated electric foot shock in C3H/HeN mice (Zhang et al., 2005). The present study observed that the injection of 6-OHDA decreased the cell numbers of IEL, IgA+, CD4+, CD8+, MC and depressed the ratio of CD4+/CD8+. However, the descending amplitude varied according to the immune-associated cell types and intestinal segments. The 6-OHDA induced the proximal intestine with a significant decrease in both cells of IgA+ and MC, whereas distal intestine presented no obvious alteration (14.1% versus 5.3% in IgA+ and 38.9% versus 8.5% in MC). In contrast, the reductions in cell numbers of CD4+ and CD8+ were larger in the distal segment than in the proximal segment (35.4% versus 0.8%, 15.4% versus 2.8%, respectively). Therefore, present results indicated that chemical sympathectomy disturbed the constancy of dynamic distribution in mucosal immune-associated cells of small intestine. Some researchers found that sympathectomy decreased the IgA+ cell number in jejunum of adult rats (Gonzalez-Ariki and Husband, 1998). On the other hand, some researchers demonstrated that noradrenaline treatment resulted in an inhibition of IgA secretion in porcine ileal (Schmidt et al., 1999). In fact, these studies were in accordance with this view.
The previous studies found that the CD8+ cell number increased ten-fold at E5 but no obvious changes to the CD4+ cell number occurred in the maternal uterus during pregnancy after 6-OHDA treatment (Dong et al., 2007). However, the present study indicated that chemical sympathectomy could dramatically decrease CD4+ cell number but slightly decrease CD8+ cell number in jejunum and ileum and then resulted in a significant reduction of CD4+/CD8+ ratio. Tsao et al. (1996) reported the reduction of CD4+ and CD8+ cell numbers in mouse thymus after 6-OHDA treatment. These data indicate the modulation of sympathetic nerves on the number change of CD4+ and CD8+ cells could vary depending on the target organ type. A similar phenomenon was also observed in the MC. The present study reported the reduced MC number in duodenum after 6-OHDA treatment. In other studies, the MC number was increased in the cardiovascular system (Facoetti et al., 2006) and skeletal muscle (Sanchez-Mejorada and Alonso-deFlorida, 1992) but was decreased in the cerebral dura mater (Artico et al., 1998), skin (Souza et al., 2005) after 6-OHDA treatment.
This study suggested that the sympathetic denervation led to an alteration of the intestinal immunological barrier by modulating the distribution of gut immune-associated cells. The study also could be contribute to further elucidation of the concrete mechanisms of intestinal inflammation, such as Crohn’s disease and Trinitrobenzene Sulphonic acid (TNBS) colitis model. Norepinephrine and dopamine tissue levels were markedly lower than in control subjects (Magro et al., 2002, 2004) in these inflamed tissues.
CONCLUSION
Consequently, chemical sympathectomy disturbed the constancy of dynamic distribution in mucosal immune-associated cells of small intestine. The present study indicated that sympathetic activity played an important role in maintaining the normal function of the intestinal immunological barrier.
ACKNOWLEDGMENTS
The study was funded by National Natural Science Foundation of China (30700575 and 30871835), Beijing Natural Science Foundation (6092016), specialized Research Fund for the Doctoral Program of Higher Education from Chinese Ministry of Education (20100008110022) and program for Cheung Kong Scholars and Innovative Research Team in University of China (IRT0866). We would like to express this gratitude to Dr. Vanessa Enriquez, Colorado State University, USA, for her critical reading of this![]()
![]() manuscript.
manuscript.
REFERENCES
- Artico, M., S. de Santis and C. Cavallotti, 1998. Cerebral dura mater and cephalalgia: Relationships between mast cells and catecholaminergic nerve fibers in the rat. Cephalalgia, 18: 183-191.
CrossRef - Bailey, J.S., A. Rolon, C.L. Hofacre, P.S. Holt and J.L. Wilson et al., 2007. Intestinal humoral immune response and resistance to salmonella challenge of progeny from breeders vaccinated with killed antigen. Int. J. Poult. Sci., 6: 417-423.
CrossRefDirect Link - Bar-Shira, E., D. Sklan and A. Friedman, 2003. Establishment of immune competence in the avian galt during the immediate post-hatch period. Dev. Comp. Immunol., 27: 147-157.
PubMed - Brunner, T., D. Arnold, C. Wasem, S. Herren and C. Frutschi, 2001. Regulation of cell death and survival in intestinal intraepithelial lymphocytes. Cell Death Differentiation, 8: 706-714.
Direct Link - Cima, I., N. Corazza, B. Dick, A. Fuhrer and S. Herren et al., 2004. Intestinal epithelial cells synthesize glucocorticoids and regulate T cell activation. J. Exp. Med., 200: 1635-1646.
Direct Link - Deshmukh, C.D., A.T. Pawar and V. Bantal, 2010. Effect of Emblica officinalis methanolic fruit extract on indomethacin induced enterocolitis in rats. Res. J. Med. Plant, 4: 141-148.
CrossRef - Dong, Y., Y. Chen, Z. Wang, J. Naito and J.L. Chen, 2007. Role of sympathetic nerves on early embryonic development and immune modulation of uterus in pregnant mice. Auton Neurosci., 131: 87-93.
CrossRefPubMedDirect Link - Elwood, C.M., A.S. Hamblin and R.M. Batt, 1997. Quantitative and qualitative immunohistochemistry of T cell subsets and MHC class-II expression in the canine small intestine. Vet. Immunol. Immunopathal., 58: 195-207.
PubMed - Emeny, R.T., D. Gao and D.A. Lawrence, 2007. Beta1-adrenergic receptors on immune cells impair innate defenses against listeria. J. Immunol., 178: 4876-4884.
PubMedDirect Link - Facoetti, A., S. Fallarini, S. Miserere, A. Bertolotti and I. Ferrero et al., 2006. Histochemical study of cardiac mast cells degranulation and collagen deposition: Interaction with the cathecolaminergic system in the rat. Eur. J. Histochem., 50: 133-140.
PubMed - Gonzalez-Ariki, S. and A.J. Husband, 1998. The role of sympathetic innervation of the gut in regulating mucosal immune responses. Brain Behavior Immunity, 12: 53-63.
PubMed - Gonzalez-Ariki, S. and A.J. Husband, 2000. Ontogeny of IgA(+) cells in lamina propria: Effects of sympathectomy. Dev. Comparative Immunol., 24: 61-69.
PubMed - Hamidi, H., R. Jahanian and J. Pourreza, 2010. Effect of dietary betaine on performance, immunocompetence and gut contents osmolarity of broilers challenged with a mixed coccidial infection. Asian J. Anim. Vet. Adv., 5: 193-201.
CrossRefDirect Link - Inagaki-Ohara, K., H. Nishimura, A. Mitani and Y. Yoshikai, 1997. Interleukin-15 preferentially promotes the growth of intestinal intraepithelial lymphocytes bearing gamma delta T cell receptor in mice. Eur. J. Immunol., 27: 2885-2891.
PubMed - Jarillo-Luna, A., V. Rivera-Aguilar, B.E. Martinez-Carrillo, E. Barbosa-Cabrera, H.R. Garfias and R. Campos-Rodriguez, 2008. Effect of restraint stress on the population of intestinal intraepithelial lymphocytes in mice. Brain Behavior Immunity, 22: 265-275.
CrossRef - Kleinschmidt, S., F. Meneses, I. Nolte and M. Hewicker-Trautwein, 2008. Distribution of mast cell subtypes and immune cell populations in canine intestines: Evidence for age related decline in T-cells and macrophages and increase of IgA-positive plasma cells. Res. Vet. Sci., 84: 41-48.
CrossRefPubMed - Levite, M. and Y. Chowers, 2001. Nerve-driven immunity: Neuropeptides regulate cytokine secretion of T cells and intestinal epithelial cells in a direct, powerful and contextual manner. Ann. Oncol., 2: 19-25.
PubMed - Liu, T., R. She, K. Wang, H. Bao and Y. Zhang et al., 2008. Effects of rabbit Sacculus rotundus antimicrobial peptides on the intestinal mucosal immunity in chickens. Poult. Sci., 87: 250-254.
CrossRef - Magro, F., S. Fraga, T. Ribeiro and P. Soares-da-Silva, 2004. Decreased availability of intestinal dopamine in transmural colitis may relate to inhibitory effects of interferon-gamma upon l-DOPA uptake. Acta physiol. Scandinavica, 180: 379-386.
CrossRef - Magro, F., M.A. Vieira-Coelho, S. Fraga, M.P. Serrao, F.T. Veloso, T. Ribeiro and P. Soares-da-Silva, 2002. Impaired synthesis or cellular storage of norepinephrine, dopamine and 5-hydroxytryptamine in human inflammatory bowel disease. Digestive Dis. Sci., 47: 216-224.
CrossRef - Vali, N., 2009. Probiotic in quail nutrition: A review. Int. J. Poult. Sci., 8: 1218-1222.
CrossRefDirect Link - Nijhuis, L.E., B.J. Olivier and W.J. de Jonge, 2010. Neurogenic regulation of dendritic cells in the intestine. Biochem. Pharmacol., 80: 2002-2008.
PubMedDirect Link - Nikfar, S., S. Ehteshami-Afshar and M. Abdollahi, 2011. A systematic review and meta-analysis of the efficacy and adverse events of infliximab in comparison to corticosteroids and placebo in active ulcerative colitis. Int. J. Pharmacol., 7: 325-332.
CrossRef - Oda, Y., F. Ueda, A. Kamei, C. Kakinuma and K. Abe, 2010. Biochemical investigation and gene expression analysis of the immunostimulatory functions of an edible Salacia extract in rat small intestine. BioFactors, 37: 31-39.
CrossRef - Ramiro-Puig, E., F.J. Perez-Cano, S. Ramos-Romero, T. Perez-Berezo and C. Castellote et al., 2008. Intestinal immune system of young rats influenced by cocoa-enriched diet. J. Nutr. Biochem., 19: 555-565.
PubMed - Resendiz-Albor, A.A., H. Reina-Garfias, S. Rojas-Hernandez, A. Jarillo-Luna, V. Rivera-Aguilar, A. Miliar-Garcia and R. Campos-Rodriguez, 2010. Regionalization of pigr expression in the mucosa of mouse small intestine. Immunol. Lett., 128: 59-67.
PubMed - Sanchez-Mejorada, G. and F. Alonso-deFlorida, 1992. Changes in mast-cell distribution in skeletal muscle after denervation. Muscle Nerve, 15: 716-719.
PubMed - Schmidt, P.T., L. Eriksen, M. Loftager, T.N. Rasmussen and J.J. Holst, 1999. Fast acting nervous regulation of immunoglobulin a secretion from isolated perfused porcine ileum. Gut, 45: 679-685.
PubMed - Souza, B.R., J.F. Cardoso, T.P. Amadeu, A. Desmouliere and A.M.A. Costa, 2005. Sympathetic denervation accelerates wound contraction but delays reepithelialization in rats. Wound Repair Regen., 13: 498-505.
CrossRef - Straub, R.H., R. Wiest, U.G. Strauch, P. Harle and J. Scholmerich, 2006. The role of the sympathetic nervous system in intestinal inflammation. Gut, 55: 1640-1649.
CrossRef - Swinkels, W.J., J. Post, J.B. Cornelissen, B. Engel, W.J. Boersma and J.M. Rebel, 2006. Immune responses in Eimeria acervulina infected one-day-old broilers compared to amount of Eimeria in the duodenum, measured by real-time PCR. Vet. Parasitol., 138: 223-233.
PubMed - Tsao, C.W., J.T. Cheng, C.L. Shen and Y.S. Lin, 1996. 6-hydroxydopamine induces thymocyte apoptosis in mice. J. Neuroimmunol., 65: 91-95.
PubMed - Waly, N., T.J. Gruffydd-Jones, C.R. Stokes and M.J. Day, 2001. The distribution of leucocyte subsets in the small intestine of healthy cats. J. Comp. Pathol., 124: 172-182.
PubMed - Yurong, Y., J. Yibao, S. Ruiping, Y. Qingqiang and P. Kaisong et al., 2006. Effects of chicken intestinal antimicrobial peptides on humoral immunity of chickens and antibody titres after vaccination with infectious bursal disease virus vaccine in chicken. Arch. Anim. Nutr., 60: 427-435.
PubMed - Zhang, X.Y., W.G. Li, Y.J. Wu and M.T. Gao, 2005. Amelioration of doxorubicin-induced myocardial oxidative stress and immunosuppression by grape seed proanthocyanidins in tumour-bearing mice. J. Pharmacy Pharmacol., 57: 1043-1052.
PubMed