
C.A. Mbajiorgu
Department of Animal Production, University of Limpopo,Private Bag X1106, Sovenga 0727, South Africa
J.W. Ng`ambi
Department of Animal Production, University of Limpopo,Private Bag X1106, Sovenga 0727, South Africa
D.D. Norris
Department of Animal Production, University of Limpopo,Private Bag X1106, Sovenga 0727, South Africa
Asian Journal of Animal and Veterinary Advances
Year: 2011 | Volume: 6 | Issue: 1 | Page No.: 20-28







ABSTRACT
The phenomenon of feed intake response trends to differing feed energy and protein levels is reviewed. Increased interest in this concept is attributed to problems associated with maintaining adequate feed intake in many farms. This becomes an important factor limiting productivity. Though the spectrum of factors that affect voluntary feed intake in poultry is very broad, it is important to highlight the influence of dietary factors, particularly, energy and protein densities on voluntary feed intake responses in chickens. In formulating poultry diets, the nutrient requirements of broiler chickens have frequently been expressed per unit of dietary metabolizable energy. This practice is based on the theory that birds will adjust their feed intake according to their metabolisable energy requirements. However, based on a re-evaluation of numerous research data, the application of specific nutrient-to-metabolisable energy ratios in broiler chickens becomes questionable. Many studies have shown that feed intake responses in chickens offered diets differing in feed energy and protein levels are influenced by the level of the first limiting nutrient in the feed rather than the feed energy level per se. This observation on limitations in feed intake, in effect, challenges the strongly held theory that all chickens will consume diets to meet their energy requirements and thereby achieve their genetic potential for growth. Thus, because of the important implications of these differences, both the energy and protein levels of the diet should be taken into account when formulating diets aimed at achieving optimal feed intake in growing chickens.
PDF Abstract XML References Citation
Received: April 25, 2010;
Accepted: April 30, 2010;
Published: July 10, 2010
How to cite this article
C.A. Mbajiorgu, J.W. Ng`ambi and D.D. Norris, 2011. Voluntary Feed Intake and Nutrient Composition in Chickens. Asian Journal of Animal and Veterinary Advances, 6: 20-28.
DOI: 10.3923/ajava.2011.20.28
URL: https://scialert.net/abstract/?doi=ajava.2011.20.28
DOI: 10.3923/ajava.2011.20.28
URL: https://scialert.net/abstract/?doi=ajava.2011.20.28
INTRODUCTION
Voluntary feed intake of chickens determines nutrient intake levels and thus has a great impact on efficiency of poultry production. Often, adequate feed intake is hard to maintain on many poultry operations in several farms and, thus, becomes an important factor limiting productivity. Stressors such as hot temperature, increased stocking density and reduced health status, together with genotype, influence feed intake and, thus, growth. Furthermore, dietary factors, including energy density, deficiencies or excesses of nutrients such as carbohydrates, protein and minerals can also influence feed intake in poultry. Though the spectrum of factors that affect voluntary feed intake in poultry is very broad, the purpose of this review is to highlight the influence of dietary factors, particularly energy and protein densities, on voluntary feed intake in chickens.
ENERGY REQUIREMENT OF CHICKENS
Energy by itself is not a nutrient but a property of energy yielding nutrients, primarily carbohydrates, lipids and proteins when they are oxidized during metabolism. Dietary energy levels have been shown to affect broiler chickens’ feed intake. Plavnik et al. (1997) and Nahashon et al. (2005) reported that as dietary energy level increases, birds satisfy their energy needs by decreasing feed intake. Decreases in feed intake with high energy levels in the diets of broiler chickens have also been reported by Leeson (2000) and Veldkamp et al. (2005). Thus, in formulating poultry diets, the nutrient requirements of broiler chickens have frequently been expressed per unit of dietary metabolisable energy (Gonzalez and Pesti, 1993). This practice is based on the theory that birds will adjust their feed intake according to their metabolisable energy requirements and was summarized by the NRC (1984) as an absolute requirement for energy in terms of kilocalories per kilogram of diet cannot be stated because poultry adjust their feed intake to obtain their necessary daily requirement. However, based on a re-evaluation of numerous research data, the NRC (1994) have revised their previous conclusions by stating that the practice of relating nutrient concentrations as a function of dietary metabolisable energy seems to apply more to leghorn type chickens fed diets with a low metabolisable energy concentration while, as a result of the over-consumption of energy on diets with a high metabolisable energy concentration, the application of specific nutrient-to-metabolisable energy ratios in broiler chickens and turkeys should be re-evaluated. Leeson et al. (1996) showed that broiler chickens fed up to 25 and 49 days of age were able to adjust their feed intake to a constant energy intake over a range of dietary metabolisable energy levels from 11.29 to 13.80 MJ ME/kg DM, which indicated that broiler chickens retain an innate ability to eat to a fixed energy requirement rather than to physical capacity as was suggested by Newcombe and Summers (1984). However, on closer observation of the data by Leeson et al. (1996), it can be seen that early feed intake to 25 days of age was not greatly affected by dietary metabolisable energy concentrations over the range of 12.13 to 13.80 MJ ME/kg DM and that it was only at the lowest metabolisable energy concentration of 11.29 MJ ME/kg DM that a significant increase in feed intake occurred. Also, the effects of metabolisable energy concentration on feed intake were very different between the early (0-25 days) and later (26-49 days) growth periods, with the metabolisable energy concentration having a far greater effect on increasing feed intake during the grower-finisher phase. This led to the overall conclusion by these authors that broiler chickens do indeed eat to a constant metabolisable energy intake when viewed over the entire 49-day growing period. In contrast to the above observation, Richards (2003) observed that modern broiler chickens selected for rapid growth do not regulate voluntary feed intake to achieve energy balance. This altered ability of broiler chickens to adjust feed intake due to differences in metabolisable energy density of the diet was postulated to result from continued selection for rapid juvenile growth rates, which may have altered hypothalamic mechanisms that regulate feed intake in broiler chickens (Burkhart et al., 1983; Bokkers and Koene, 2003). Other reports have also shown no effect of dietary metabolisable energy concentration on feed intake between two groups of broiler chickens fed ad-libitum diets containing two energy levels of 13.38 and 15 MJ ME/kg DM.
PROTEIN AND AMINO ACID REQUIREMENTS OF CHICKENS
Proteins have been described as complex organic compounds of high molecular weight composed of 22 different amino acids or derivatives that are linked by peptide bonds to form a primary chain structure. As a result of steric constraints this primary structure has been reported to form an α-helical structure stabilized by hydrogen bonds as well as by cross-linking of individual amino acid residues. The á-helix that describes the primary structure of the protein has been found to be subsequently folded and arranged into more complex secondary and tertiary structures which, with the specific number and sequences of different amino acids, ultimately determine the biological characteristics and functionality of the protein (Leeson and Summers, 2001; Horton et al., 2002). Because body proteins are in a dynamic state, with synthesis and degradation occurring continuously, an adequate intake of dietary amino acid is required. If dietary protein or amino acid is inadequate, there is a reduction or cessation of growth or productivity and a withdrawal of protein from less vital body tissues to maintain the functions of more vital tissues (NRC, 1994).
As mentioned earlier, there are 22 amino acids in body proteins and all are physiologically essential (NRC, 1994). Nutritionally, ten of these are indispensable because chickens are unable to synthesize them or can not synthesize them at a rate sufficient to meet their needs. These are methionine, lysine, threonine, leucine, valine, isoleucine, arginine, phenylalanine, histidine and trytophan (Austic, 1995; NRC, 1994). The amino acid requirements of poultry represent the requirements for the indispensable amino acids plus sufficient nitrogen in an appropriate chemical form for synthesis of the dispensible amino acids. Chickens are sensitive to the dietary balance of these amino acids (Austic, 1995). For the diet to be used with maximum efficiency, the chicken must receive the indispensable amino acids in the correct quantities and sufficient amino acids to meet the dispensable amino acids for metabolic demands must be available. The presence of adequate amounts of nonessential amino acids in the diet reduces the necessity of synthesizing them from essential amino acids. Amino acid requirements may be classified as those for maintenance, carcass growth, egg production and feather growth on the basis of their respective amino acid profiles (Hurwitz et al., 1978). In order for the bird to realize its genetic potential and achieve the best levels of performance through maximum rates of protein synthesis, amino acids must be provided in the necessary quantities, avoiding both excesses and deficiencies (Sainbury, 1984). Thus, stating dietary requirements for both protein and essential amino acids is an appropriate way to ensure that all amino acids needed physiologically are provided.
Protein and amino acid requirements vary considerably according to the physiological state of the bird, that is, the rate of growth or egg production. Other factors contributing to variations in amino acid requirements of the chickens include age, body size, sex and breed. Amino acid requirements decrease with age and at the same time, the ideal balance of amino acids changes gradually to reflect those of maintenance (Zubair and Leeson, 1996). For instance, the percentage of amino acid required in the diet is the highest for young growing animals and declines gradually to maturity, when only enough amino acid to maintain body tissue is required (Pond et al., 1995). The balance of amino acids needed for maintenance is not proportional to the balance of amino acids in a bird’s tissues, but rather reflects the relative rate of obligatory loss of each individual amino acid (Gous and Morris, 1985). For this reason, the balance needed for maintenance is considerably different from that needed for growth or egg production (Nemavhola, 2001). Dietary amino acid levels slightly below maintenance can sustain life, but muscle mass and functions are impaired (Leeson, 1996). Matching the amino acid profile of the diet with animal requirements is crucial for maximizing animal performance. For instance, turkey poults and broiler chickens have high amino acid requirements to meet the needs for rapid growth while the indigenous chickens such as the Venda breed will require less amino acid to meet their needs because of their slow growth rate and small body size. Because the contributions of maintenance and growth to total amino acid requirement change with body size and the ideal amino acid profiles for maintenance and growth are different, the composition of the ideal amino acid pattern will change continuously during the growth period (Mack et al., 1999).
| Table 1: | Amino acid requirements (g kg-1 feed) at different ages of broiler chickens |
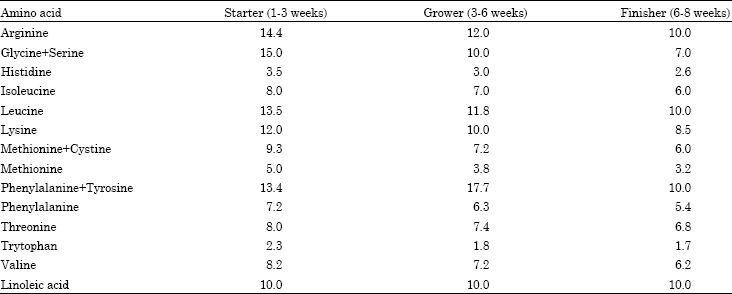 | |
| Source: NRC (1994) | |
Amino acid requirements at different ages of broiler chickens are shown in Table 1, it is now well documented that male broiler chickens have higher dietary amino acid requirements than females (Han and Baker, 1993; Thomas et al., 1986), because male chickens contain more protein and less fat in their weight gain (Edwards et al., 1973; Han and Baker, 1991).
Unlike the effect of diet energy concentration, the effect of protein density on feed intake responses in broiler chickens has not been consistent. Buyse et al. (1992) reported that broiler chickens reared on lower protein density of 15% crude protein in the diet increased their feed intake in an attempt to meet their protein requirement. Contrary to these findings, a decrease in feed intake with reduced protein density has been reported in broiler chickens by Kemp et al. (2005) and Berhe and Gous (2005). These authors observed that Ross 308 broiler chickens decreased their feed intake as dietary protein content was reduced, resulting in a lower growth rate.
RESPONSE TRENDS OF CHICKENS TO DIFFERING FEED ENERGY AND PROTEIN LEVELS
To be of any real value, attempts to optimize the feeding of chickens must be capable of predicting voluntary food intake. Gous (2007) suggested that where feed intake is seen as an input, as is most often the case, it is not possible to optimize feeding programs successfully since the composition of the food offered has a very important effect on voluntary food intake. As suggested by Emmans and Fisher (1986) appetite is dependent on the nutrient requirements of the animal and the contents of those nutrients in the feed and hence, responses in feed intake, therefore, are not independent of the composition of the feed and strain of the chicken as was previously believed (Hill and Dansky, 1954).
The theory of feed intake and growth in birds proposed by Emmans (1981, 1989) was based on the premise that birds attempt to grow at their genetic potential, which would imply that they would attempt to eat as much of a given feed as would be necessary to grow at that rate. Factors that would prevent them from achieving this goal would be the bulkiness of the feed or the inability to lose sufficient heat to the environment in order to enable them to remain in thermal balance. This theory has been shown to predict feed intake and hence growth and carcass composition with considerable accuracy in birds (Ferguson and Gous, 1997; Ferguson et al., 1997). Additionally, Cobb 500 broiler chickens (Burnham et al., 1992) and laying hens (Gous et al., 1987) have been shown to increase feed intake as dietary protein content in the feed is reduced, attempting thereby, to obtain more of the limiting protein irrespective of the feed energy level until a dietary concentration is reached where performance is so constrained that feed intake falls. Similarly, Mbajiorgu (2010) observed that indigenous Venda chickens increased their feed intake with increase in feed energy level and with decrease in feed protein content. This is contrary to the observation that broiler chickens eat to satisfy their energy requirements (Leeson, 2000; Nahashon et al., 2005, 2006; Veldkamp et al., 2005), or that broiler chickens will eat less of a feed higher in energy content than the one having a lower energy value (Palvink et al., 1997; Nahashon et al., 2006; Veldkamp et al., 2005). These findings together suggest that feed intake of broiler chickens is, first and foremost, closely linked to the feed energy level and hence birds attempt, as a priority, to adjust their feed intakes according to the energy level of the diet.
As suggested by Mbajiorgu (2010), indigenous Venda chickens, however, tended to behave differently in this respect. Tadelle et al. (2000) suggested that genetic limitation influences indigenous chicken growth responses because it affects their nutritional requirements. Thus, one possible consequence of the intrinsic genetic limitations of indigenous Venda chickens might be the loss of sensitivity to regulate feed intake according to dietary energy level. The physiological explanation for the present observation in indigenous Venda chickens is not clear and merits further investigation. However, it has been shown that chickens will increase their feed intake in response to marginal levels of first limiting feed nutrient, independent of the diet energy level (Boorman, 1979) since appetite is assumed to be dependent on the nutrient requirements of the animal and the contents of those nutrients in the feed (Emmans and Fisher, 1986). As such, feed intake of indigenous Venda chickens may have increased regardless of the energy value of the feed. Thus, Venda chickens ate more feed in an attempt to meet their protein requirements, which were limiting with decreasing dietary crude protein levels. This observation is similar to the results obtained with broiler chickens by Burnham et al. (1992) and with laying hens by Gous et al. (1987). These authors observed that chickens increased their feed intake as the limiting nutrient in the feed decreased in an attempt to obtain more of the limiting nutrient to satisfy their requirements for that nutrient. In fact, the nutritional factors involved in broiler chicken feed intake control mechanisms are not completely understood. Parsons et al. (1993) pointed out that in many experiments, where only responses to dietary energy level are involved, such feed intake responses could be confounded with variable intake of other nutrients such as protein and hence differences in feed intake response patterns to limiting feed protein content observed for Ross 308 broiler chickens and Cobb 500 chickens as indicated in Fig. 1 below. Importantly, it is interesting to note that these differing feed intake response patterns to limiting feed protein content were achieved regardless of the energy value of the feed. Contrary to the above observations, Kemp et al. (2005) and Berhe and Gous (2005) observed that the Ross 308 strain of broiler chickens does not apparently conform to the theory that birds attempt to consume sufficient of a feed to meet their requirement for the first limiting nutrient in the feed as proposed by Boorman (1979) and supported by the work of Emmans and Fisher (1986). These authors observed that instead of increasing food intake, the Ross 308 broiler chicken strains decreased their feed intake as dietary energy was increased and dietary protein content reduced, resulting in a lower growth rate than in the Cobb 500 strain whose feed intake increases as dietary protein content is reduced (Fig. 1).
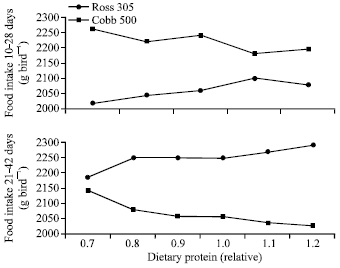 | |
| Fig. 1: | Response in food intake of two strains of broiler to increasing dietary balanced protein contents. Trial 1 from Kemp et al. (2005). Dietary protein contents relative to the Ross standard feeding levels (Aviagen, 2006) |
They concluded that Ross 308 broiler chickens have been selected for improved growth and feed efficiency using high protein feeds. The authors went further to emphasize the point that such selection results in heavier carcasses (Pym and Solvens, 1979) and perhaps a reduced ability to fatten when faced with feeds marginally deficient in protein. Harper and Rogers (1965) suggested that when there is a dietary protein deficit, the free amino acid patterns of both muscle and plasma become imbalanced and consequently trigger the appetite regulating system to reduce feed intake. This may be the scenario when Ross 308 broiler chickens receive feeds marginally deficient in protein unlike the Cobb 500 and indigenous Venda chickens. Apparently, genetic potential may influence the Ross 308 broiler chickens’ feeding behaviour as it affects their nutritional requirements (Gous et al., 1999). Ross 308 broiler chickens have a pronounced genetic advantage for fast growth using high protein feed compared to Cobb 500 and Venda chickens and this might explain the differences in feed intake response patterns to marginally limiting feed protein content.
These observations on limitations in feed intake response patterns in Ross 308 broiler chickens, Cobb 500 and indigenous Venda chickens contradict the strongly held theory that all chickens eat to satisfy their energy requirements (Hill and Dansky, 1954; Scott et al., 1982; Leeson et al., 1996) or that chickens will eat less of a feed higher in energy content than the one having a lower energy value (Nahashon et al., 2006; Palvink et al., 1997; Veldkamp et al., 2005). However, because of the important implications of these differences, both the energy and protein levels of the diet should be taken into account when formulating diets aimed at achieving optimal feed intake in growing birds.
Apparently, the above observations by Boorman (1979), Gous et al. (1987), Burnham et al. (1992), Richards (2003) and Mbajiorgu (2010) on limitations in feed intake support the revised thinking of the NRC (1994) that some chicken strains do not adjust their feed intake to changes in the dietary metabolisable energy density and, as a result, may be prone to over-consume metabolisable energy in an attempt to obtain sufficiency of a limiting nutrient when offered diets high in energy, thereby, making the long held theory that all chickens do adjust feed intake to a constant metabolisable energy intake to necessitate further investigation.
CONCLUSION
It is, therefore, concluded that feed intake responses in chickens offered diets differing in feed energy and protein levels are influenced by the level of the first limiting feed nutrient rather than the feed energy level per se. This observation on limitations in feed intake in effect challenges the strongly held theory that all chickens will consume diets to meet their energy requirements and thereby achieve their genetic potential for growth. However, because of the important implications of these differences, both the energy and protein levels of the diet should be taken into account when formulating diets aimed at achieving optimal feed intake in broiler chickens.
REFERENCES
- Bokkers, E.A.M. and P. Koene, 2003. Eating behaviour and preprandial and postprandial correlations in male broiler and layer chickens. Br. Poult. Sci., 44: 538-544.
CrossRefDirect Link - Burkhart, C.A., J.A. Cherry, H.P. Van Krey and P.B. Siegel, 1983. Genetic selection for growth rate alters hypothalamic satiety mechanisms in chickens. Behav. Genet., 13: 295-300.
CrossRefDirect Link - Burnham, D., G.C. Emmans and R. Gous, 1992. Isoleucine requirements of the chicken: The effect of excess leucine and valine on the response to isoleucine. Br. Poult. Sci., 33: 71-87.
Direct Link - Buyse, J., E. Decuypere, L. Berghman, E.R. Kuhn and F. Vandesande, 1992. The effect of dietary protein content on episodic growth hormone secretion and on heat production of male broilers chickens. Br. Poult. Sci., 33: 1101-1109.
PubMedDirect Link - Gonzalez, M.J. and G.M. Pesti, 1993. Evaluation of the protein to energy ratio concept in broiler and turkey nutrition. Poult. Sci. J., 72: 2115-2123.
PubMedDirect Link - Gous, R.M. and T.R. Morris, 1985. Evaluation of a diet dilution technique for measuring the response of broiler chickens to increasing concentrations of lysine. Br. Poult. Sci., 26: 147-161.
Direct Link - Gous, R.M., M. Griessel and T.R. Morris, 1987. Effect of dietary energy concentration on the response of laying hens to amino acids. Br. Poult. Sci., 28: 427-436.
Direct Link - Gous, R.M., E.T. Moran, H.R. Stilborn, G.D. Bradford and G.C. Emmans, 1999. Evaluation of the parameters needed to describe the overall growth, the chemical growth and the growth of feathers and breast muscles of broilers. Poult. Sci., 78: 812-821.
Direct Link - Gous, R.M., 2007. Predicting nutrient responses in poultry: Future challenges. Animal, 1: 57-65.
Direct Link - Han, Y.M. and D.H. Baker, 1991. Lysine requirement of fast and slow growing broiler chicks. Poult. Sci., 70: 2108-2114.
Direct Link - Han, Y. and D.H. Baker, 1993. Effects of sex, heat stress, body weight and genetic strain on the lysine requirement of broiler chicks. Poult. Sci., 72: 701-708.
Direct Link - Hurwitz, S., D. Sklan and I. Bartov, 1978. New formal approaches to the determination of energy and amino acid requirements of chicks. Poult. Sci., 57: 197-205.
PubMed - Leeson, S., L. Caston and J.D. Summers, 1996. Broiler response to diet energy. Poult. Sci., 75: 529-535.
CrossRefDirect Link - Mack, S., D. Bercovici, G. de Groote, B. Leclercq and M. Lippens et al., 1999. Ideal amino acid profile and dietary lysine specification for broiler chickens of 20-40 days of age. Br. Poult. Sci., 40: 257-265.
Direct Link - Mbajiorgu, C.A., 2010. Effect of dietary energy and protein ratio level on growth and productivity of indigenous Venda chickens raised in closed confinement from day-old up to 13 weeks of age. Ph.D. Thesis, Department of Animal production, Faculty of Science and Agriculture, University of Limpopo, South Africa.
- Nahashon, S.N., N. Adefope, A. Amenyenu and D. Wright, 2005. Effect of dietary metabolisable energy and crude protein concentrations on growth performance and carcass characteristics of French guinea fowl broilers. Poult. Sci. J., 84: 337-344.
Direct Link - Nahashon, S.N., N. Adefope, A. Amenyenu and D. Wright, 2006. Effect of varying metabolisable energy and crude protein concentrations in diets of pearl gray guinea fowl pullets 1. Growth performance. Poult. Sci. J., 85: 1847-1854.
Direct Link - NRC., 1994. Nutrient Requirements of Poultry. 9th Edn., National Academy Press, Washington, DC., USA., ISBN-13: 9780309048927, Pages: 176.
Direct Link - Parsons, C.M., K.W. Koelkebeck, Y. Zhang, X. Wang and R.W. Leeper, 1993. Effect of dietary protein and added fat levels on performance of young laying hens. J. Applied Poult. Res., 2: 214-220.
Direct Link - Plavnik, I., E. Wax, D. Sklan, I. Bartov and S. Hurwitz, 1997. The response of broiler chickens and turkey poults to dietary energy supplied either by fat or carbohydrates. Poult. Sci., 76: 1000-1005.
Direct Link - Pym, R.A.E. and A.J. Solvyns, 1979. Selection for food conversion in broilers: Body composition of birds selected for increased body‐weight gain, food consumption and food conversion ratio. Br. Poult. Sci., 20: 87-97.
CrossRefDirect Link - Richards, M.P., 2003. Genetic regulation of feed intake and energy balance in poultry. Poult. Sci., 82: 907-916.
Direct Link - Tadelle, D., Y. Alemu and K.J. Peters, 2000. Indigenous chickens in Ethiopia: Genetic potential and attempts at improvement. World's Poult. Sci. J., 56: 45-54.
CrossRefDirect Link - Zubair, A.K. and S. Leeson, 1996. Compensatory growth in the broiler chicken: A review. World's Poult. Sci. J., 52: 189-201.
CrossRefDirect Link - Veldkamp, T., R.P. Kwakkel, P.R. Ferket and M.W.A. Verstegen, 2005. Growth responses to dietary energy and lysine at high and low ambient temperature in male Turkeys. Poult. Sci. J., 84: 273-282.
PubMed