
H.K. Lim
School of Biotechnology, Hankyong National University, Anseong-City,
456-749, Korea
K. Choi
School of Biotechnology, Hankyong National University, Anseong-City,
456-749, Korea
P.K. Mandal
Department of Food Science and Biotechnology of Animal Resources, Konkuk University, Seoul, Korea
O. Baatartsogt
School of Biotechnology, Hankyong National University, Anseong-City,
456-749, Korea
C.H. Lee
Department of Food Science and Biotechnology of Animal Resources, Konkuk University, Seoul, Korea
J.H. Lee
Division of Animal Science and Resources, College of Agriculture and Life Sciences, Chungnam National University, Daejeon 305-764, Korea
H.B. Kim
Laboratory of Bioinformatics and Population Genetics, Department of Food and Animal Biotechnology, Seoul National University, Seoul, Korea
Asian Journal of Animal and Veterinary Advances
Year: 2009 | Volume: 4 | Issue: 2 | Page No.: 66-75







ABSTRACT
This study was carried out to investigate the differentially expressed genome between S. gallinarum infected and uninfected control in the spleen lymphocytes of Ross broiler chicks using microarray analysis. GeneChip Chicken Genome Array containing 32,773 transcripts corresponding to over 28,000 chicken genes for simultaneous expression was used. The signal intensity of each gene was normalized and expressed in fold change. A large numbers of genes were found with differential expression majority of which are still unknown in chicken genome. Thirty one known genes were found to have differential expression of which, 25 were up-regulated and 5 were down regulated. Majority of the up-regulated genes belong to immune response system viz., IL8, IL1B, IL10, IL18, IL17A, IL15, transferrin, IFNg, TLR2, TNFRSF1b, TNFRSF15 and the down regulated genes were B-FIV, B-LA, SDF1, B-LBI, belonging to MHC-I and II and CD1d. To validate the expression of these genes RT-PCR was done using primers of 12 selected genes′ with total mRNA isolated from spleen lymphocytes which has confirmed the similar pattern of expression of all the genes as in microarray. The findings in this study have lead to the identification of novel genes which may be useful in further studies to understand the patho-physiology of fowl typhoid towards development of diagnostics and therapeutics.
PDF Abstract XML References Citation
How to cite this article
H.K. Lim, K. Choi, P.K. Mandal, O. Baatartsogt, C.H. Lee, J.H. Lee and H.B. Kim, 2009. Transcriptional Profiling of Spleen Lymphocyte in Fowl Typhoid of Broilers. Asian Journal of Animal and Veterinary Advances, 4: 66-75.
DOI: 10.3923/ajava.2009.66.75
URL: https://scialert.net/abstract/?doi=ajava.2009.66.75
DOI: 10.3923/ajava.2009.66.75
URL: https://scialert.net/abstract/?doi=ajava.2009.66.75
INTRODUCTION
Fowl typhoid caused by Salmonella gallinarum (SG) is recognized worldwide as a disease of social and economic significance. Salmonella enterica serotype gallinarum is a non-motile, host adapted avian pathogens belonging to Salmonella serogroup D (Shivaprasad, 1997). The out break of fowl typhoid is characterized by increased mortality, anorexia, greenish-yellow diarrhea and a drop in egg production (Brown et al., 2007). Subacute outbreak leads to egg transmission to chicks which increases dead or weak chicks. Though, it has largely been eradicated from countries with intensive poultry industry for many years, fowl typhoid caused by SG is still of considerable economic significance to the poultry industry in many countries of Africa, the Middle East, Central and South America and Asia (Shivaprasad, 2000). The quick detection of this pathogen is therefore, extremely important.
Molecular understanding of the mechanism of the disease is important for developing new diagnostic tools and in designing of disease specific therapies. Conventional Kaufmann White scheme is still the only reliable method for serotyping of Salmonella, however, it is time consuming and cumbersome (Ewing, 1986). Use of classical methods based on biochemical tests /assays are tedious and time consuming (Christensen et al., 1992; Shah et al., 2001). Moreover, recent reports of intermediate strains with variable biochemical pattern casts doubts on the validity of these biochemical assays (Jia et al., 1993; Shah et al., 2005).
The large scale analysis of gene function, is central to functional genomics. Parallel quantitative display of genes is considered the most promising strategy for biomarker discovery. Genomic analysis, which refers to large-scale study of gene expression and function has gained great interest (Pandey and Mann, 2000). Recently, genomic analysis has gained great interest for the determination of biochemical processes involved in diseases (Fung et al., 2000). The comparative characterization of gene expression patterns in tissues has the potential to serve as the basis for new diagnostic tools and in designing of disease specific therapies (Sinz et al., 2002). Genomics is gaining popularity in the research on animal and poultry diseases. Because of its biomedical utility, the poultry will imminently join those vertebrates that have representative genomes sequenced (Burt and Pourquie, 2003).
There are recent reports of gene expression due to Salmonella enteritidis infection in chicken spleen (Zhou and Lamont, 2007) chicken macrophage (Withanage et al., 2004) and chicken intestine (Hemert et al., 2006). However, cellular and molecular mechanisms of fowl typhoid is not yet well understood, because of lack of large number of biomarkers for SG infection process in the poultry. Gene expression in response to Salmonella in vivo and in vitro in chicken mainly focused on cytokines and chemokines in various tissues (Kaiser et al., 2000; Withanage et al., 2004; Cheeseman et al., 2006). However, no detailed study on genome analysis of fowl typhoid has been done yet.
Microarray technology is expected to revolutionize the biological research field through the simultaneous analysis of gene expression patterns in the whole genome scale. Hence, GeneChip Chicken Genome Array containing comprehensive coverage of 32,773 transcripts corresponding to over 28,000 chicken genes was used for the simultaneous investigation of gene expression changes in spleen lymphocyte in chickens. The spleen plays a vital role in pathogenesis of Salmonella infection of chickens (Hendersson et al., 1999). Therefore, this study was carried out to investigate the differentially expressed genome between S. gallinarum infected and uninfected control in the spleen lymphocytes of chicken using microarray analysis.
MATERIALS AND METHODS
Experimental Plan
Twenty, one-day-old Ross broiler chicks were procured from Yanggi Hatchery, Pyeongtaek, S. Korea. After six weeks of normal feeding the chicks were randomly divided into two groups. One group was kept away as uninfected control the other group was infected by intramuscular injection with 1 mL of S. gallinarum (2.9x107 mL-1). Randomly, 3 chickens were selected from each group after one week of infection and spleens were collected after appropriate anaesthesia. Chicken spleens were pooled together and lymphocytes were separated (Histopaque H-1070, Sigma).
Isolation of Total RNA from Spleen Lymphocyte
Total RNA was isolated from chicken spleen lymphocyte samples collected from control and infected chicken using a Trizol Reagent (Invitrogen, USA). One milliliter of Trizol reagent was added to a 10 cm diameter dish with cells and the cells were passed through a pipette several times. The homogenized sample was incubated for 5 min at room temperature to permit the complete dissociation of nucleoprotein complexes. The sample was mixed with 0.2 mL of chloroform per 1 mL of Trizol reagent by shaking vigorously by hand for 15 sec and incubated for 2 to 3 min. Then, the samples were centrifuged at 12000 rpm for 15 min at 4°C. Following centrifugation, the mixture was separated into a lower red phenol-chloroform phase, an interphase and a colorless upper aqueous phase. The aqueous phase was transferred to a fresh tube and precipitated the RNA from the aqueous phase by mixing with isopropyl alcohol. Then 0.5 mL isopropyl alcohol was added per 1 mL Trizol reagent used for the initial homogenization and samples were incubated at room temperature for 10 min and centrifuged at 12000 rpm for 10 min at 4°C. RNA pellet was washed by with 1 mL of 75% ethanol per 1 mL of Trizol reagent. The RNA pellet was briefly dried and 20 μL of RNAase-free water was added and incubated for 10 min at 56°C. Finally, the RNA concentration was measured in the UV spectrophotometer.
Hybridising and Analysis of Microarray
GeneChip Chicken Genome Array which contains comprehensive coverage of 32,773 transcripts corresponding to over 28,000 chicken genes was used for the simultaneous investigation of gene expression changes in spleen lymphocyte in chickens.
The generation of GeneChip data from RNA isolated from spleen lymphocytes of normal and infected chicken was performed by Seoulin Bioscience Corporation (Seoul, Korea). About 5 μg total RNA from the normal and infected chicken cells were used for labeling. Probe synthesis from total RNA samples, hybridization, detection and scanning were performed according to standard protocols from Affymetrix (California, USA). Briefly, cDNA was synthesized using the One-Cycle cDNA Synthesis Kit (Affymetrix). Single-stranded (ss) cDNA was synthesized using Superscript II reverse transcriptase and T7-oligo (dT) primers at 42°C for 1 h. Double-stranded (ds) cDNA was obtained using DNA ligase, DNA polymerase I and RNase H at 16°C for 2 h, followed by T4DNA polymerase at 16°C for 5 min. After cleanup using a Sample Cleanup Module (Affymetrix, CA), ds cDNA was used for in vitro transcription (IVT). cDNA was transcribed using the GeneChip IVT Labeling Kit (Affymetrix) in the presence of biotin-labeled CTP and UTP. Then the biotin-labeled IVT-RNA was fragmented and hybridized to the porcine genome GeneChip array at 45°C for 16 h, according to the manufacturer′s instructions. After hybridization, the arrays were washed in a GeneChip Fluidics Station 450 with a non-stringent wash buffer at 25°C, followed by a stringent wash buffer at 50°C. After washing, the arrays were stained with a streptavidin-phycoerythrin complex. After staining, intensities were determined with a GeneChip scanner, controlled by GeneChip Operating Software (GCOS; Affymetrix).
RT-PCR Analysis
To validate the differential expression of genes in infected chicken spleen lymphocytes semi-quantitative RT-PCR analysis was performed using specific primers for 12 selected genes and GAPDH as control (Table 1). cDNA was synthesized from RNA isolated from lymphocytes using a SuperScriptTM II (Invitrogen, USA). The thermo cycler profile was 5 min 94°C (initial denaturation) and then 22-30 cycles of 40 sec at 94°C (denaturation), 30 sec at 60-65°C (annealing), 40 sec at 72°C (extension) and followed by a 10 min final extension at 72°C. The PCR products were analyzed by electrophoresis (2.0% agarose gels) in 40 mM Tris-acetate containing 1 mM EDTA (1x TAE) (in ethidium bromide fluoresce). Quantitative analysis of PCR product was done using a Sigma Gel software (Jandel Scientific) comparing with GAPDH standard.
| Table 1: | Primers used for validation of the expression of selected genes in mRNA from spleen lymphocytes of fowl typhoid infected and uninfected control chicken in RT-PCR analysis |
 | |
 | |
| Fig. 1: | Image of gene chip hybridized with mRNA from spleen lymphocyte of control (A) and Salmonella gallinarum infected chicken (B) gene chip |
RESULTS
This experiment was conducted to study the gene expression profile in fowl typhoid by applying microarray analysis. Six week old chicks were experimentally infected with S. gallinarum and lymphocytes from spleen of control and Salmonella gallinarum infected chicken were isolated. The gene expression profiles of spleen lymphocytes using GeneChip® Chicken Genome Array were analyzed and results are explained (Fig. 1).
Genome-wide microarray analysis showed differential expression of 802 genes, 430 genes were upregulated of which 25 are known and 372 genes were down regulated of which 6 are known in chicken cells. However, a large number of differentially expressed genes have not yet been described in chicken genome database. Gene Chip Operating Software (GCOS) and randomized microarray (RMA) statistical analysis showed reproducibly differentially expressed genes. The differentially expressed genes were functionally annotated with reports from literature. Visualization of these differentially expressed genes by hierarchical clustering demonstrated that expression of spleen lymphocytes from normal and infected chicken were similar to each other. Gene Ontology (http://www.geneontology.org; GO) annotations were determined for each gene product represented. GO terms are consistent descriptions of gene products on terms of the biological processes they are involved in, the cellular components in which they exist and molecular functions they perform.
| Table 2: | Up regulated genes as expressed in spleen lymphocyte between fowl typhoid infected and uninfected control chicken in microarray analysis |
 | |
| Table 3: | Down regulated genes as expressed in spleen lymphocyte between fowl typhoid infected and uninfected control chicken in microarray analysis |
 | |
A total of 31 known genes were found to have differential (2 fold change) expression of which 25 genes were up-regulated and 6 genes were down regulated. Among the up-regulated genes majority belong to immune response system (Table 2) and the down regulated genes mostly belong to MHC-I and MHC-II (Table 3). The important up-regulated expression of genes observed in spleen lymphocytes are IL1B, IL8, IL10, IL15, IL17A, IL18,IL 21/22 transferrin, IFNg, TLR2, TNFRSF1B, TNFRSF15 and the down regulated genes were B-FIV, B-LA, SDF1, B-LBI, belonging to MHC-I and II and CD1d. To validate the expression of genes RT-PCR was done with mRNA isolated from spleen lymphocytes which has confirmed the similar expression of all the 12 genes (Fig. 2). A comparison of gene expression in microarray and RT-PCR analysis is presented in Table 4.
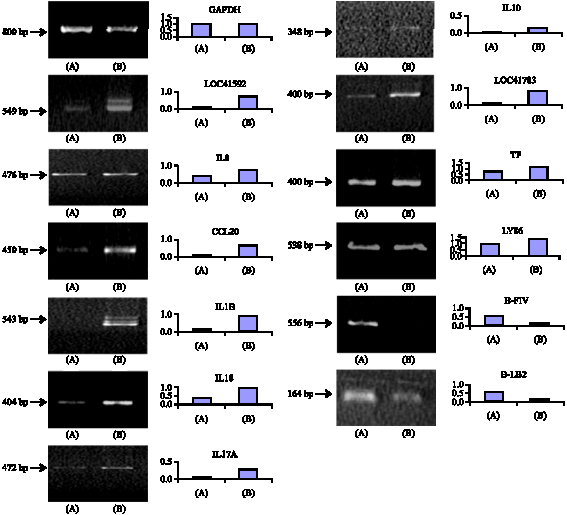 | |
| Fig. 2: | Comparison of expressions of selected genes in mRNA from spleen lymphocyte of fowl typhoid infected and uninfected control chicken in RT-PCR analysis |
| Table 4: | Comparision of the level of differential expressions of selected genes in spleen lymphocyte between fowl typhoid infected and uninfected control chicken in microarray and RT-PCR analysis |
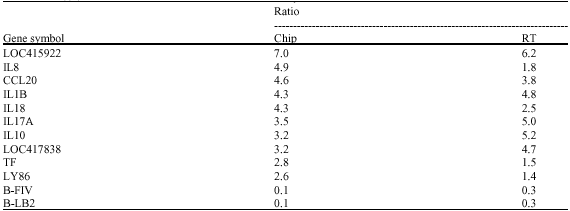 | |
DISCUSSION
Study of the interaction between host and pathogen at molecular level has become a major research area in functional genomics. In this study a search was made for biomarkers in fowl typhoid, though, no report is available about the functional genomic search on fowl typhoid. Differential expression of cytokines and chemokines such as interleukin-6 (IL-6), IL-1b, IL-10, IL-12a, IL-12b, IL-18, CXCR1, CXCR4 with infection of Salmonella enteritidis or Salmonella typhimurium in chickens and chicken lymphocytes have been reported in many studies (Kaiser et al., 2000; Kogut et al., 2003; Beal et al., 2005; Withanage et al., 2004, 2005; Wigley et al., 2005, 2006; Cheeseman et al., 2006). Microarray technology has the advantage of simultaneously investigating the expression of thousands of genes, to enable the study of gene interactions and signal pathways. The current study utilized a chicken microarray to investigate differential gene expression between Salmonella gallinarum inoculated and non-inoculated birds.
This is the first study that has determined the genome-wide molecular expression pattern of spleen lymphocyte from experimental fowl typhoid and thus provided comprehensive insight into avian salmonellosis. In the present study experimental fowl typhoid was successfully induced by intramuscular inoculation in 6 weeks old chickens and the same has been confirmed by pathogen isolation, gross necropsy finding and clinical symptoms. Large number of important genes were found with differential expression using microarray analysis many of them are related to immune response system which were upregulated (IL1B, IL8, IL10, IL15, IL17A, IL18, transferrin, IFNg, TLR2). A number of genes related to MHC class I and class II (B-FIV, B-LA, SDF1, B-LBI) chemokine CXC and CD1d were down regulated.
Chemokines, cytokines and their receptors play very important roles in Salmonella infection in chickens (Withanage et al., 2004). Many chemokines, cytokines and their receptors were differentially expressed with SG infection in the current study and IL 1b is an important among them. In poultry IL1B recognized as pro-inflammatory and stimulates immune systems. IL1B activates a range of cell including macrophages and T cell which lead to produce other cytokine and chemokines (Wigley and Kaiser, 2003). The up-regulated expression of IL1B gene might lead to the high level of this cytokines which as a consequence lead to fever (Wigley and Kaiser, 2003). IL-1b mRNA level was elevated with infection of SG in the present study. An increased IL-1b mRNA level was also observed in heterophils (Ferro et al., 2004) and in spleen tissue (Zhau and Lamont, 2007) chicken infected with SE. Withanage et al. (2004) reported higher mRNA level of IL-1b in spleen, cecum, ileum and liver at different time points post-infection with S. typhimurium in Rhode Island Red chickens. A higher level of IL-1b mRNA was detected in spleen and cecal tonsils at day 6 with challenge of S. typhimurium in birds (Beal et al., 2005). Upregulation of pro-inflammatory cytokine might be crucial to recruit lymphocyte cells to the site of infection, to subsequently mount an efficient innate and adaptive immune response to infection.
IL8 is a chemokine having chemotactic activity for specific leukocytes (Wuyts et al., 1998) which appears to specifically activate neutrophils in response to infection in human. In chicken IL8 is equivalent to mammalian IL8 though functionally it is designated as chicken chemotactic and angiogenic factor (CAF) (Kaiser et al., 1999). It has been reported that IL8 produced an influx of heterophils in chicken gut with S. typhimurium infection (Henderson et al., 1999) which suggested the induction of IL8. Therefore, it appears to be natural to have induction of this chemokine at gene label in spleen lymphocytes in present study as also reported in mammalian Salmonella infection induced IL8 in the gut epithelium (Wallis and Galyov, 2000).
IL 15 in mammals is a T cell growth factor stimulate the growth of T lymphocytes, NK cells (Kennedy et al., 1998). IL15 induces proliferation, cytokine production and migration of T cells and also stimulates proliferation of number of other cells including B lymphocytes. Recently, it has been proposed that IL-15 plays a major role in driving autoimmune thyroiditis in obese strains of chickens (Kaiser et al., 2002). IL-18 in mammals induces production of IFN-g (Okamura et al., 1995). It is produced at high levels in the liver by macrophages and Kupffer cells and plays important role in the initiation of cell mediated immune response (Wigley and Kaiser, 2003). The chicken IL-18 gene is a quarter in length and a bioassay has shown the similar response of chicken IL-18 as its human counterpart (Kaiser et al., 2001). IFN-g is type II interferon which plays vital role in macrophage activation and modulation of the immune system (DeMayer and DeMayer, 1988). It is produced primarily by T lymphocytes and natural killer (NK) cells leading to activation of macrophage antimicrobial activity. It has potent macrophage activating factor activity that is heat and pH labile (Lowenthal et al., 1995).
Besides, several other chemokines, their receptors and cytokines were also differentially regulated in the current study as mentioned earlier. Upregulated gene expression of MIP-1, IL-8, CXCR4 with infection of SE and/or S. typhimurium has been found in other studies (Ferro et al., 2004; Swaggerty et al., 2004, 2006; Withanage et al., 2004; Wigley et al., 2006). The upregulation of these genes suggests that these cytokines are important in the initiation of the pro-inflammatory response to defend the host against SE infection. Several genes related to MHC were down regulated in this study. However, several genes of the T-cell surface glycoprotein family were significantly upregulated in the broiler with SE infection (Zhou and Lamont, 2007). This might be a different kind of response in case of SG which needs further confirmation. The CD complex, is essential for signal transduction of the T cell receptor and proper T-cell responses against infections (Janeway et al., 1999). Upregulation of the CD3 complex in chickens with SE infection was reported by Zhau and Lamont (2007) to be associated with activation of the T cell receptor, which would then modulate T cell immune response to fight SE infection. However, in this study a down regulation of CD1 was observed.
The search for differentially expressed genes is an important means to understand pathological pathways to find markers of disease for diagnosis and for treatments. There are recent efforts towards this direction for Salmonella enteritidis in chicken spleen (Zhou and Lamont, 2007) and chicken intestine (Hemert et al., 2006). As far as we know it has not been tried before for understanding the fowl typhoid using spleen lymphocytes of chicken with microarray analysis, hence it is difficult to compare the observations. The current study presents the microarray analysis Salmonella gallinarum infection in chickens and provides an new initiative to study molecular and cellular mechanisms of gene expression in fowl typhoid disease. These findings demonstrated the power of microarray technology to study molecular mechanisms of Salmonella infection in chickens. This also has laid a foundation to further study cellular and molecular mechanisms of Salmonella gallinarum infection in poultry.
ACKNOWLEDGMENT
This research was fully supported by a grant from Biogreen21 program, Rural Development Administration, Republic of Korea.
REFERENCES
- Beal, R.K., C. Powers, P. Wigley, P.A. Barrow, P. Kaiser and A.L. Smith, 2005. A strong antigen-specific T-cell response is associated with age and genetically dependent resistance to avian enteric salmonellosis. Infect. Immunol., 73: 7509-7516.
PubMed - Burt, D. and O. Pourquie, 2003. Genetics. Chicken genome science nuggets to come soon. Science, 300: 1669-1669.
PubMed - Cheeseman, J.H, M.G. Kaiser, C. Ciraci, P. Kaiser and S.J. Lamont, 2006. Breed effect of early cytokine mRNA expression in spleen and secum of chickens with and without Salmonella eteritidis infection. Dev. Comp. Immunol., 31: 52-60.
PubMed - Christensen, J.P., J.E. Olsen, H.C. Hansen and M. Bissgard. 1992. Characterization of Salmonella enterica serovar gallinarum biovars gallinarum and pullorum by plasmid profiling and biochemical analysis. Avian Pathol., 21: 461-470.
PubMed - Ferro, P.J., C.L. Swaggerty, P. Kaiser, I.Y. Pevzner and M.H. Kogut, 2004. Heterophils isolated from chickens resistant to extra-intestinal Salmonella enteritidis infection express higher levels of pro-inflammatory cytokine mRNA following infection than heterophils from susceptible chickens. Epidemiol. Infect., 132: 1029-1037.
PubMedDirect Link - Fung, E.T., G.L. Jr. Wright and E.A. Dalmasso, 2000. Proteomic strategies for biomarker identification: Progress and challenges. Curr. Opin. Mol. Ther., 2: 643-650.
Direct Link - Hemert, S.V., A.J.W. Hoekman, M.A. Smits and J.M.J. Rebel, 2006. Gene expression responses to a Salmonella infection in the chicken intestine differ between lines. Vet. Immunol. Immunopathol., 114: 247-258.
PubMed - Henderson, S.C., D.I. Bounous and M.D. Lee, 1999. Early events in the pathogenesis of avian salmonellosis. Infect. Immunol., 67: 3580-3586.
PubMed - Li, J., N.H. Smith, K. Nelson, P.B. Crichton, D.C. Old, T.S. Whittam and R.K. Selander, 1993. Evolutionary origin and radiation of the avian-adapted non-motile salmonellae. J. Med. Microbiol., 38: 129-139.
PubMed - Kaiser, P., S. Hughes and N. Bumstead. 1999. The chicken 9E3/CEF4 CXC chemokine is the avian orthologue of IL8 and maps to chicken chromosome 4 syntenic with genes flanking the mammalian chemokine cluster. Immunogenetics, 49: 673-684.
PubMed - Kaiser, P., L. Rothwell, E.E. Galyov, P.A. Barrow, J. Burnside and P. Wigley, 2000. Differential cytokine expression of avian cells in response to invasion by Salmonella typhimurium, Salmonella enteritidis and Salmonella gallinarum. Microbiology, 146: 3217-3226.
PubMed - Kaiser, P., L. Rothwell, D. Vasicek and K. Hala, 2002. A role for IL-15 in driving the onset of spontaneous autoimmune thyroiditis. J. Immunol., 168: 4216-4220.
PubMed - Kogut, M.H., L. Rothwell and P. Kaiser, 2003. Differential regulation of cytokine gene expression by avian heterophils during receptor-mediated phagocytosis of opsonized and nonopsonized Salmonella enteritidis. J. Interferon Cytokine Res., 23: 319-327.
PubMed - Lowenthal, J.W., M.R. Digby and J.J. York, 1995. Production of interferon-y by chicken T cells. J. Interferon Cytokine Res., 15: 933-938.
PubMed - Pandey, A. and M. Mann, 2000. Proteomics to study genes and genomes. Nature, 405: 837-846.
CrossRefDirect Link - Okamura, H., H. Tsutsi, T. Komatsu, M. Yutsudo, A. Hakura and T. Tanimoto, 1995. Cloning of new cytokine that induces IFN-gamma production by T cells. Nature, 378: 88-91.
PubMed - Shah, D.H., A. Roy and J.H. Purohit, 2001. Characterization of Salmonella gallinarum avian strains isolated from Gujarat State. Indian J. Comp. Microbiol. Immunol. Infect., 22: 131-133.
Direct Link - Shah, D.H., J.H. Park, M.R. Cho, M.C. Kim and J.S Chae, 2005. Allele-specific PCR method based on rfbS sequence for distinguishing Salmonella gallinarum from Salmonella pullorum: Serotype-specific rfbS sequence polymorphism. J. Microbiol. Methods, 60: 169-177.
CrossRefPubMedDirect Link - Shivaprasad, H.L., 2000. Fowl typhoid and Pullorum disease. Rev. Sci. Technol. Office Int. Epizootics, 19: 405-424.
PubMed - Sinz, A., M. Bantscheff, S. Mikkat and B. Ringel, 2002. Mass spectrometric proteome analysis of synovial fluids and plasmas from patients suffering rheumatoid arthritis and comparison to reactive arthritis or osteoarthritis. Electrophoresis, 23: 3445-3456.
Direct Link - Swaggerty, C.L., M.H. Kogut, P.J. Ferro, L. Rothwell, I.Y. Pevzner and P. Kaiser, 2004. Differential cytokine mRNA expression in heterophils isolated from Salmonella-resistant and-susceptible chickens. Immunology, 113: 139-148.
PubMed - Swaggerty, C.L., P. Kaiser, L. Rothwell, I.Y. Pevzner and M.H. Kogut, 2006. Heterophil cytokine mRNA profiles from genetically distinct lines of chickens with differential heterophil-mediated innate immune responses. Avian Pathol., 35: 102-108.
PubMed - Wallis, T.S. and E.E. Gaylov, 2000. Molecular basis of Salmonella-induced enteritis. Mol. Biol., 36: 997-1005.
PubMed - Wigley, P. and P. Kaiser, 2003. Avian cytokines in health and disease. Brajilian J. Poult. Sci., 5: 1-14.
Direct Link - Wigley, P., S. Hulme, C. Powers, R. Beal, A. Smith and P. Barrow, 2005. Oral infection with the Salmonella enteric serovar Gallinarum 9R attenuated live vaccine as a model to characterise immunity to fowl typhoid in the chicken. BMC Vet. Res., 1: 2-2.
PubMed - Wigley, P., S. Hulme, L. Rothwell, N. Bumstead, P. Kaiser and P. Barrow, 2006. Macrophages isolated from chickens genetically resistant or susceptible to systemic salmonellosis show magnitudinal and temporal differential expression of cytokines and chemokines following Salmonella enterica challenge. Infect. Immunol., 74: 1425-1430.
PubMed - Withanage, G.S., P. Kaiser, P. Wigley, C. Powers and P. Mastroeni et al., 2004. Rapid expression of chemokines and proinflammatory cytokines in newly hatched chickens infected with Salmonella enterica serovar typhimurium. Infect. Immunol., 72: 2152-2159.
CrossRefPubMedDirect Link - Withanage, G.S.K., P. Wigley, P. Kaiser, P. Mastroeni and H. Brooks et al., 2005. Cytokine and chemokine responses associated with clearance of a primary Salmonella enterica serovar Typhimurium infection in the chicken and in protective immunity to rechallenge. Infect. Immunol., 73: 5173-5182.
CrossRefPubMedDirect Link - Zhou, H. and S.J. Lamont, 2007. Global gene expression profile after Salmonella enterica Serovar enteritidis challenge in two F8 advanced intercross chicken lines. Cytogenet. Genome Res, 117: 131-138.
PubMed