
Nabila Berrighi
Laboratory of Food Technology and Nutrition, Mostaganem University, Algeria
Louiza Belkacemi
Laboratory of Food Technology and Nutrition, Mostaganem University, Algeria
Kaddour Bouderoua
Laboratory of Food Technology and Nutrition, Mostaganem University, Algeria
Marina Santaella
Department of Food Technology, Nutrition and Food Science, Veterinary Faculty University of Murcia, Campus de Espinardo,
30100 Espinardo, Murcia, Spain
Gaspar Ros
Department of Food Technology, Nutrition and Food Science, Veterinary Faculty University of Murcia, Campus de Espinardo,
30100 Espinardo, Murcia, Spain
Gema Nieto
Department of Food Technology, Nutrition and Food Science, Veterinary Faculty University of Murcia, Campus de Espinardo,
30100 Espinardo, Murcia, Spain
Asian Journal of Animal Sciences
Year: 2017 | Volume: 11 | Issue: 2 | Page No.: 88-95







ABSTRACT
Background and Objective: Interest in meat fatty acid composition stems mainly from the need to find ways to produce healthier meat, with a higher ratio of polyunsaturated to saturated fatty acids and a more favorable balance between n-6 and n-3. The purpose through this study is to assess the effect of diet on fatty acids and colour characteristics of lamb meat. Materials and Methods: In this study two separate groups of Rembi breed was used either reared in a highland area and feeding on pasture supplemented with hay or reared in a steppe area and feeding on pasture supplemented with concentrate. The study was carried out from March to June, 2015. After slaughter, samples of the Biceps femoris were removed from each carcass and placed in ice in isothermal boxes to be transported to the laboratory. The dissected muscles were trimmed, minced in a meat grinder and stored at -20°C for further analysis. Results: The results showed more fat in the meat from the steppe group lambs than the highland one (3.80 vs 1.94%, respectively, p<0.05). The polyunsaturated fatty acid (PUFA) levels were higher in meat issued from lambs reared in the steppe than those from the highland region. The n-3 polyunsaturated fatty acid and C18:3 n-3 levels were significantly higher in the tissue from the highland group, but there was no significant difference between the highland and the steppe lamb meat with regards to C18:2 n-6 levels. The C18:2 n-6/C18:3 n-3 and n-6/n-3 ratios could be considered ideal for human health in both groups: 6.55 and 8.04% for highland animals, respectively (p<0.001) and 10.50 and 12.68% for the steppe-bred animals (p<0.001), respectively. Overall, the feeding system played an important role in the colour of the resulting meat and the results point to significant differences in the amounts of colour parameters i.e., redness, luminosity and yellowness. Conclusion: Lamb meat produced on Algerian steppe and highland areas seems to have better sensoriel and nutritional quality, especially in term of fatty acids quality, which linked to the graze diversity and quality present in these lands. Based on these results and from a fattening perspective, grazing is declared to be an economically sustainable husbandry system.
PDF Abstract XML References Citation
Received: December 20, 2016;
Accepted: January 23, 2017;
Published: February 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Nabila Berrighi, Louiza Belkacemi, Kaddour Bouderoua, Marina Santaella, Gaspar Ros and Gema Nieto, 2017. Fatty Acids Composition and Sensory Properties of Lamb Meat Fed on Steppe and Highland Pastures. Asian Journal of Animal Sciences, 11: 88-95.
DOI: 10.3923/ajas.2017.88.95
URL: https://scialert.net/abstract/?doi=ajas.2017.88.95
DOI: 10.3923/ajas.2017.88.95
URL: https://scialert.net/abstract/?doi=ajas.2017.88.95
INTRODUCTION
In steppe, sheep farming is one of the most traditional agricultural activities and occupies a very important place in domain of animal production and is the first red meat provider. According to the Algerian Ministry of Agriculture, the total sheep population was estimated to 22 million head in 2015 and grazing in arid regions, mainly steppe, covered 12 million hectares. Thus, because of its importance, it plays an essential function in the rural economy and is actively involved in the production of red meats.
Lamb production in steppe and some other regions is primarily based on year-round extensive finishing systems. Under these finishing systems, animals are often feeding on widely differing diets. It is known that diet has a major effect on polyunsaturated fats in meat1. Pannier et al.2 reported that extensive lamb production sometimes produces meat with high levels of long chain omega-3 fatty acids, but not always. This is not surprising because of the wide range of finishing diets used in extensive grazing systems. Production factors, such as breed, age, feeding system or food composition have been shown as important constituents in the fatty acid composition of lamb3.
Changing the fatty acid profile of lamb meat to obtain a lower proportion of Saturated Fatty Acids (SFA) is an important way to produce a healthier meat for the consumer. According to the literature, this can be achieved through the use of lipid sources in the diet1,4.
The colour of lamb meat is crucial to ensure customer appeal and strongly contributes to the product value. Meat that remains red during display is stable in colour and lamb meat is regarded as less stable than beef meat5. The aim of this study is to investigate the main effects of the rearing system i.e., feeding diets in the steppe and the highlands on meat quality.
MATERIALS AND METHODS
Study location: The study was carried out on 27 month-old lambs, with an average body weight (W) of 32.2±2.38 kg. Lambs were divided into two equal groups (n = 10). The first group was put out to pasture from the 21th March to the 21th June, 2014 on a highland farm (latitude 35°37'85"N and longitude 1°32'57"E) and the second group was reared on a steppe farm (latitude 34°66'67"N and longitude 0°3'25"E).
Description of studied farms: The lambs were randomly selected from a large flock of sheep. The steppe and highland pastures were divided into five paddocks about 1 ha each one and the animals were allowed to graze freely on young spring grass until the end of the spring season, with a free access to water. On the both farms and during the experiment, the lambs were put out to pasture twice a day from 7-11 am and from 4-6 pm.
Data collection and recording: The sampling plan for the diets consisted of a randomly hand-picking about 3-4 kg of crude grass throughout all of the territories grazed for further analysis. The pastures consisted of small and medium sized herbaceous forage resistant to arid conditions. The dominant species of plants in the steppe pasture were wormwood (Artemisia herba-alba), halfa grass (Stipa tenacissima), sparta grass (Lygeum spartum) and Atriplex halimus, with various other important species in the highland, such as Zizyphus lotus, Pistacia atlantica, Capsella bursa pastoris, ray-grass (Lolium perenne L.), laser white (Laserpitium latifolium), kava fruit (Pometia pinnata), salad rocket (Eruca sativa), knotweed bistort (Bistorta officinalis) and sweet broom (Arthrophytum scoparium). The highland lambs were also given hay and the steppe animals a standard concentrate feed in order to attain a desirable commercial body weight. The composition of the diets is shown in Table 1.
Collecting samples: After three months of grazing, all the lambs from both highland and steppe groups were slaughtered and eviscerated in a local commercial slaughterhouse. Samples of the Biceps femoris were removed from each carcass and placed in ice in isothermal boxes to be transported to the laboratory. The dissected muscles were trimmed, minced in a meat grinder and stored at -20°C for further analysis.
Diet and meat analyses: The chemical composition was determined in both diet and meat. Both samples were placed in a drying oven at 103°C for 24 h and 550°C for 4 h to determine respectively the dry matter and crude ash contents6. The total lipid content of each sample was extracted using chloroform-methanol (2:1) according to the method described by Folch et al.7.
| Table 1: | Diets composition in percentage |
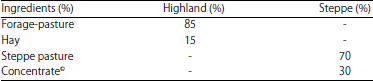 | |
| ©Concentrate was composed of 15% corn, 10% soybean meal, 4% brain and 1% minerals | |
The neutral detergent fiber, acid detergent fiber and acid detergent lignin were measured using the Van Soest method8. The crude proteins were determined according to the Kjeldahl method6. The total phenol in the diets was measured according to Borowska et al.9. Ground forage (10 g) was twice extracted with 80 mL of methanol containing 0.1% HCL. After mechanical stirring, the homogenate was centrifuged at 4.000 rpm for 15 min and the supernatant was collected. Finally, the methanolic extract was concentrated under reduced pressure at 60°C using a rotary evaporator BUCHIR R-210.
Fatty acids profile: Extracted lipids were dissolved in hexane and the fatty esters were methylated by addition of 2 N potassium hydroxide in methanol. Separation and quantification of the different fatty acids were made by gas liquid chromatography, using an Agilent Technologies 7890A chromatograph, (Agilent Technologies Palo Alto, CA, USA) equipped with a Flame Ionization Detector (FID) operated with a split ratio of 20:1. The column was a DB-23 (60 m, 0.25 mm i.d., 0.25 mm coating thickness (Agilent technologies). The injector and detector temperatures were held constant at 250 and 280°C, respectively. The oven temperature was programmed at 50°C for 1 min and ramped at 25°C min–1 to 175°C followed by further ramping at 4°C min–1 to 220°C which was held constant for 18 min. The carrier gas, helium was held at a constant pressure of 230 kPa. Chromatographic air and hydrogen (400 and 35 mL min–1, respectively) were supplied to the FID. Methyl esters were identified by comparing the retention times with those of known Fatty Acid Methyl Ester (FAME) standards (Supelco 37 component FAME mix, both from Supelco, St., Louis, MO, USA). The relative proportion of each fatty acid in the meat was reported as a percentage of the total fatty acid in the meat (FAME) present in the injected sample. The sum of the saturated, monounsaturated, polyunsaturated and highly unsaturated fatty acids and the ratio between n-3 and n-6 fatty acids were also calculated.
TBARS measurement: The measurement of "TBA" index was performed according to the method adapted by Genot10. A sample of 2 g meat was placed in a 25 mL tube containing 16 mL of 5% (w/v) trichloroacetic acid and optionally 100 μL of ascorbic acid (Vitamin C). The mixture was homogenized 3 times for 15 sec using a homogenizer (Ultra-turrax) at 20,000 rpm. The ground material was passed through a filter paper to obtain a filtrate. From this filtrate 2 mL were added to 2 mL of thiobarbituric acid. The closed tubes were immersed in a water bath at 70°C during 30 min then in a cold water bath. Thereafter, the absorbance of the reaction mixture was measured at 532 nm using UV-visible spectrophotometer (JENWAY) and the results were expressed in mg equivalent MDA (malondialdehyde) kg–1.
Colour of meat: The color was determined through a slice of meat about 1 cm thick which was cut to expose a portion of the muscle in the air. After 30 min of oxygenation, parameters luminosity (L*), the redness index (a*) and the yellowness index (b*) were determined with a colorimeter (Chroma meter CR data processor 300 and DP-301, Minolta Co., Ltd., Japan).
Statistical analysis: The data analyses included a single-factor analysis of variance with a block organization and an average pair wise comparison. The data were first analysed using descriptive methods (mean, standard deviation, box-plot and based on the criterion of dietary effect) and then by multivariate analysis (principal component analysis applied to game data online, including the fatty acids, lipid stability and colour in the muscles from steppe and highland-raised animals. The statistical calculations were carried out using a monofactorial variance analysis (ANOVA), followed by a comparison of the means according to the Newman-Keuls test. The results are presented in terms of p-values, with a level of significance p<0.05.
RESULTS
Chemical composition of the diets: The results for the diets are summarized in Table 2. In general, there was a slight difference (p<0.05) in the crude protein content between the steppe diet and the highland one the neutral and acid detergent fibers and acid detergent lignin were much higher for the highland forage than the corresponding steppe forage. However, the percentages of crude fiber were higher in the pasture and concentrate given to the steppe group lambs than in the pasture and hay given to the highland animals (Table 2). Moreover, the fat content was higher in steppe pasture (3.34%) and concentrate (4.73%) than in the highland pasture (3.29%) and hay (1.34%), (p<0.05). The steppe regimen saturated and monounsaturated fatty acids content was much higher than the highland one (Table 2). The FA profile shows that the oleic content of the steppe diet was higher. At the same time, the α-linolenic acid was clearly higher in the steppe pasture and concentrate than in the highland diet (Table 2).
Meat composition: The meat composition (dry matter, moisture, ash and fat) showed significant differences (p<0.05) between the highland and steppe groups.
| Table 2: | Diet chemical composition in percentage |
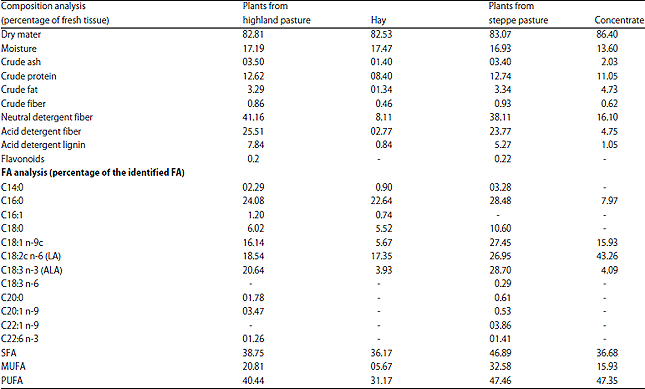 | |
| n = 4 | |
| Table 3: | Chemical characteristics of lamb raw meat |
 | |
| n = 10, SEM: Standard error of the mean | |
Concerning the protein content, it was more important in the highland than in the steppe group (15.50 vs. 15.41%, respectively, p<0.05) (Table 3).
Meat fatty acids: The meat total lipids contents from steppe diet were lower than those from pastures highland (Table 4). In comparison with steppe group, meat from highland lambs displayed more SFA (p<0.05). However, the PUFA displayed much more substantial quantities in meat from the steppe group (p<0.05). The total LA/ALA, n-6/n-3 and PUFA/SFA were significantly higher (p<0.001) in the lamb meat issued from steppe area (Table 4).
Sensory properties: Table 5 underline the low degradation that occurs in meat as a result of oxidation. An important level of MDA was observed in the lamb meat from steppe than those from highland one (respectively 0.13 vs 0.09 mg MDA kg–1 meat), even though no statistically no significant difference was reported. The results showed significant effects of the feeding system on meat colour (Table 5) between the steppe and highland groups. The L*, a* and b* values increased (p<0.05) in the steppe lamb meat and decreased in the highland one (p<0.05).
DISCUSSION
The composition of the improved pasture used had a lower dry matter whose the level was comparable to those of Blankson et al.11 study. On the contrary, the concentration of protein in the steppe diet was higher. As to fatty acid composition, the pasture from the steppe contained higher C16:0 (palmitic acid) and C18:0 (stearic acid) concentrations than those of the highland.
| Table 4: | Meat fatty acid composition (percentage of identified FA) |
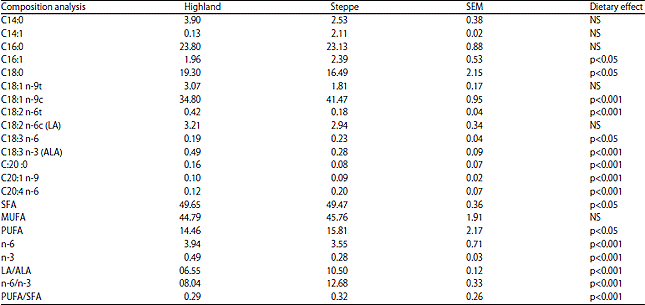 | |
| n = 10, SEM: Standard error of the mean | |
| Table 5: | Sensory properties (MDA content and colour) of lamb raw meat |
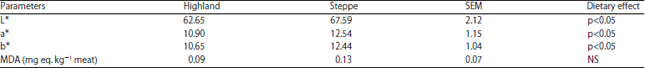 | |
| n = 10, SEM: Standard error of the mean, L*: Luminosity, a*: Redness, b*: Yellowness | |
These dietary proportions agreed with the findings of a previous study12. The C18:3 n-3 was as well as much higher in the steppe pasture compared to the highland one (28.70 vs 20.64%, respectively) than in the steppe concentrate compared to hay in the highland (4.09 vs 3.93%, respectively). These results are comparable to those obtained by Aurousseau et al.13. The concentrations of MUFA, represented by oleic acid (C18:1 n-9) and the PUFA percentages (C18:2 n-6 linoleic and C18:3 n-3 α-linolenic acid) were higher in the steppe pasture and concentrate compared to the highland pasture and hay. McAfee et al.14 have shown that long chain omega-3 PUFA, such as eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) have beneficial effects over other fatty acids present in diets for the maintenance of long term health. The DHA level was observed to be considerably higher in steppe than in the highland pasture, most likely due to much more green pasture associated with more rainfall15. More recently, nutritionists have focused on the type of PUFA and the balance in the diet between n-3 PUFA formed from α-linolenic acid (18:3) and n-6 PUFA formed from linoleic acid (18:2)16.
There were significant differences (p<0.05) in the chemical meat composition between the two groups. This is due to the variety of the botanical species and the differences in soil between the two regions. In addition, under grazing conditions, animal might select differently a specific roughage species. Our finding suggest that the diets had no significant effect on the protein content of meat except for the higher intramuscular fat content observed in the meat from the lambs grazing on the steppe. Despite exhibiting differences, the protein contents in the meat were within the expected range17. The results suggests that the total protein content is less variable in lamb meat observed in the biceps femoris muscles and this is independent of diet, breed, the genetic group and the physiological conditions18.
The low average fat content of 2.8% in the lamb meat steppe group is consistent with the results of Nold et al.19 who have reported that intramuscular lipids of semimembranosus, biceps femoris and gluteus medius had 2.15, 2.12 and 1.75%, respectively.
The meat from the steppe lambs fed on the pasture and concentrate diet showed a lower intramuscular lipid content (1.94%) than those from group fed with highland pasture and hay (3.80%). The muscle fatty acids can be affected by factors such as diet, species, over/underweight, age, gender, breed and season4,20. In the present study, the fatty acid composition differed between the meats and was affected by the diets. The mean average of stearic acid (C18:0) content was significantly higher in the highland meat, while oleic acid was the predominant fatty acid in the muscles of the steppe lamb meat (41.47 vs 34.80% respectively, p<0.001). These findings are consistent with those of a previous study of Popova21. Trans-oleic acid (C18:1n-9t) levels showed no significant differences between the two groups in this study, while another researcher has found a high accumulation in the rumen of conjugated linoleic acid via Δ9 desaturase22. In the present study, meat contained, in average, 15.81% PUFA, 45.76% MUFA and 49.47% SFA in the steppe lamb meat and 14.46% PUFA, 44.79% MUFA and 49.65% SFA in the highland one. The MUFA and PUFA concentrations obtained in this study were higher than those obtained by Padre et al.23. The effect of rearing on fatty acid composition seems to be limited to differences in SFA and MUFA concentration. Alfaia et al.24 attributed the higher protection from bio-hydrogenation in the rumen to the fatty acids contained in fresh pastures relative to grains and silage. The levels of the PUFA were particularly higher in the meat from the lambs grazing in the steppe (15.81%) than in highland (14.46%) (p<0.05). This finding is similar to the results of the study Adnoy et al.25, who collected samples from different pastures including mountains. This could first be explained by the higher botanical diversity found in the steppe and/or by the higher proportion of legumes, such as Lotus pedunculatus and Trifolium repens (white clover), growing in the steppe. Lourenco et al.26 reported that lambs fed a grass and legume mixture had higher proportions of polyunsaturated fatty acids in their muscle tissues than lambs fed grass only. There is consensus among researchers cited that the higher PUFA n-3 content in pastures accounts for the deposition of C18:3 n-3 in the meat and in the present study, the meat from pasture highland had 2 fold more α-linolenic acid than those from the steppe lambs. Although a large part of that fatty acid is hydrogenated in the rumen to stearic acid, a proportion escapes ruminal fermentation and is absorbed at the small intestine3. There was significantly higher proportion of C20:4 n-6 in the muscles from the steppe group than in those from the highland one (0.20 vs 0.12%, respectively, p<0.001). These findings were probably related to the fatty acids metabolism, season or rearing system27. The fatty acid concentrations were linear in terms of the amount of total fat, but their slope depended on the diet concentrate-based diets have higher PUFA n-6 content24; however, in the present study, no differences in C18:2 n-6 concentration in the meat of lambs fed on highland and steppe pastures were observed (respectively 3.21 and 2.94% in average) (Table 4). There was a significant effect of feeding on the concentrations of n-3 PUFA, the C18:3 n-3 (p<0.001), resulting in a greater total ratio of n-3 fatty acids (p<0.001), with higher values for fed-pasture lambs as well in highland than in the steppe. Sanudo et al.3 reported that the grass fed animals had higher muscle concentrations of n-3 PUFA and the concentrate fed animals had higher concentrations of the n-6 PUFA. Pasture-based systems rather than concentrate-based feeding systems increase the n-6/n-3 and PUFA/SFA ratios in cattle muscle lipids28. However, the LA/ALA and n-6/n-3 ratios which are much more desirable for human health have higher proportions in the steppe lamb meats (10.50 and 12.68%, respectively, p<0.001). A significant decrease in the LA/ALA and n-6/n-3 ratios in the grass fed highland lambs was observed (6.55 and 8.04%, respectively, p<0.001). According to Raes et al.22, the ratio of n-6:n-3 PUFA is particularly low in ruminant meats, especially from animals that have consumed grass which contains high levels of 18:3. This equally applies to the omega 3 content of lamb meat derived from lambs that have previously been grown in green pasture, meeting the nutritional recommendations of the World Health Organization (WHO), with regard to the nutritionally positive effects of polyunsaturated n-3 fatty acids on the prevention of cardiovascular disease, obesity and cancer. Fatty acid compositions in meat have received an increased interest considering these implications for human health and product quality29. The ratios of polyunsaturated fatty acids to saturated fatty acids are widely used to evaluate the nutritional value of fat.
In the present study there was no effect on the lipid stability values between the both groups. However, lipid oxidation increased in the meats of animals receiving concentrate as a lipid source, exceeding the cut off value of 0.13 mg MDA kg–1 of meat. This important oxidation possibly occurred due to the greater PUFA concentrations found in the biceps femoris muscles from these animals. According to McKenna et al.30, the muscular rate of lipid oxidation may also act as an indicator of the degree of meat pigment susceptibility to oxidation, bearing in mind the close relationship between these two oxidation processes.
The meat color data are summarized in Table 5 and discussed with regard to the dietary effects between highland and steppe (p<0.05). For the L*, a* and b* values, the changes in meat color are probably best represented by variations in lightness (L*). The color indices (L*, a* and b*) of legs muscles from the animals that consumed pastures and concentrate exhibited a consistent increase (p<0.05) and lamb meat from the steppe is more red. This fact is consistent with the statement that the effect of nutrition on meat color, especially the redness index (a*) is associated with the instability of heme pigments31. This could possibly be explained by difference in oxygen uptakes of post mortem muscles, so the more lamb meat absorbs oxygen, the more it is exposed to oxidation. Faustman and Cassens32 reported that lipid oxidation could promote myoglobin oxidation.
Consequently, the factors affecting lipid oxidation in meat can also influence meat color). The selection for muscles reduced oxidative capacity of muscle in sheep, which would lead to the observed improvement in colour. Recent study has shown that the color of the meat can be influenced by the weight of the carcass at slaughter, because heavier carcass is associated with more red and darker meat33.
CONCLUSION
The results of this study suggest that steppe and highland diets play important roles in the fatty acid composition of lamb meat. The proportion of n-3 PUFA was higher in the highland lamb meat; however, the C18:2 n-6/C18:3 n-3 ratio in steppe lamb meat was more consistent with the recommendations of nutritionists with regard to the highland one. Overall, the impact of the feeding regimen on the colour of meat from lambs fed concentrates is darker than that of lambs fed with grass. Changes in the a* values appear to be driven by lipid oxidation and are strongly correlated with the TBARS values.
Finally, the results from this study show that lambs allowed to graze in natural pastures, especially in the steppe and highland, presented a modified meat composition which provides a significant increase in a functional lipids which have positive influences on health.
SIGNIFICANCE STATEMENT
Sheep farming is one of the most traditional agricultural activities and has a big impact in the rural economy and for the livestock industry. Ruminant meats from animals allowed to feed on steppe and highland pastures are natural and organic products with sensory qualities much appreciated in terms of colour and fatty acid profile. Concentrate elevated PUFA contents and increased susceptibility of muscle fat to lipid oxidation; however, the lamb meat produced exclusively on highland pastures presented higher concentration of components that are considered beneficial to human health, such as n-3 fatty acids and a lower n- 6/n-3 ratio. Grass is richer in polyunsaturated fatty acids than concentrates. This modification of fatty acid profile is very important because in the last decades consumers’ perception of red meat has been influenced by alarming rumours, mainly related to nutritional implication on human health, leading to an indiscriminate and unjustified avoidance of these products. Therefore, this study revalorizes lamb meat and livestock industry.
ACKNOWLEDGMENT
The authors thank to Antonia Tovar from the Veterinary Faculty of the University of Murcia for technical assistance.
REFERENCES
- Daley, C.A., A. Abbott, P.S. Doyle, G.A. Nader and S. Larson, 2010. A review of fatty acid profiles and antioxidant content in grass-fed and grain-fed beef. Nutr. J., Vol. 9.
CrossRefDirect Link - Pannier, L., E.N. Ponnampalam, G.E. Gardner, D.L. Hopkins and A.J. Ball et al., 2010. Prime Australian lamb supplies key nutrients for human health. Anim. Prod. Sci., 50: 1115-1122.
CrossRefDirect Link - Sanudo, C., M.E. Enser, M.M. Campo, G.R. Nute, G. Maria, I. Sierra and J.D. Wood, 2000. Fatty acid composition and sensory characteristics of lamb carcasses from Britain and Spain. Meat Sci., 54: 339-346.
CrossRefDirect Link - Wood, J.D., M. Enser, A.V. Fisher, G.R. Nute and P.R. Sheard et al., 2008. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci., 78: 343-358.
CrossRefDirect Link - Gutzke, D. and G.R. Trout, 2002. Temperature and pH dependence of the autoxidation rate of bovine, ovine, porcine, and cervine oxymyoglobin isolated from three different muscles-Longissimus dorsi, Gluteus medius and Biceps femoris. J. Agric. Food Chem., 50: 2673-2678.
CrossRefDirect Link - Folch, J., M. Lees and G.H.S. Stanley, 1957. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem., 226: 497-509.
CrossRefPubMedDirect Link - van Soest, P.J., J.B. Robertson and B.A. Lewis, 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci., 74: 3583-3597.
CrossRefPubMedDirect Link - Borowska, E.J., B. Mazur, R.G. Kopciuch and B. Buszewski, 2009. Polyphenol, anthocyanin and resveratrol mass fractions and antioxidant properties of cranberry cultivars. Food Technol. Biotechnol., 47: 56-61.
Direct Link - Blankson, H., J.A. Stakkestad, H. Fagertun, E. Thom, J. Wadstein and O. Gudmudsen, 2000. Conjugated linoleic acid reduces body fat mass in overweight and obese humans. J. Nutr., 130: 2943-2948.
PubMedDirect Link - Luciano, G., F.J. Monahan, V. Vasta, P. Pennisi, M. Bella and A. Priolo, 2009. Lipid and colour stability of meat from lambs fed fresh herbage or concentrate. Meat Sci., 82: 193-199.
CrossRefDirect Link - Aurousseau, B., D. Bauchart, E. Calichon, D. Micol and A. Priolo, 2004. Effect of grass or concentrate feeding systems and rate of growth on triglyceride and phospholipid and their fatty acids in the M. longissimus thoracis of lambs. Meat Sci., 66: 531-541.
CrossRefPubMedDirect Link - McAfee, A.J., E.M. McSorley, G.J. Cuskelly, B.W. Moss, J.M.W. Wallace, M.P. Bonham and A.M. Fearon, 2010. Red meat consumption: an overview of the risks and benefits. Meat Sci., 84: 1-13.
CrossRefPubMedDirect Link - Ponnampalam, E.N., K.L. Butler, M.B. McDonagh, J.L. Jacobs and D.L. Hopkins, 2012. Relationship between muscle antioxidant status, forms of iron, polyunsaturated fatty acids and functionality (retail colour) of meat in lambs. Meat Sci., 90: 297-303.
CrossRefPubMedDirect Link - Rotta, P.P., R.M. do Prado, I.N. do Prado, M.V. Valero, J.V. Visentaine and R.R. Silva, 2009. The effects of genetic groups, nutrition, finishing systems and gender of Brazilian cattle on carcass characteristics and beef composition and appearance: A review. Asian-Aust. J. Anim. Sci., 22: 1718-1734.
CrossRefDirect Link - De Menezes, L.F.G., G.V. Kozloski, J. Restle, F.C. Deschamps, I.L. Brondani, A.P. dos Santos and J. Fiamoncini, 2006. Long chain fatty acids profile and meat quality of feedlot finished steers with different levels of sodium monensin at the diet. Cienc. Rural, 36: 186-190.
CrossRefDirect Link - Nold, R.A., J.R. Romans, W.J. Costello and G.W. Libal, 1999. Characterization of muscles from boars, barrows and gilts slaughtered at 100 or 110 kilograms: Differences in fat, moisture, color, water-holding capacity and collagen. J. Anim. Sci., 77: 1746-1754.
PubMedDirect Link - Popova, T., 2007. Effect of the rearing system on the fatty acid composition and oxidative stability of the M. longissimus lumborum and M. semimembranosus in lambs. Small Rumin. Res., 71: 150-157.
CrossRefDirect Link - Raes, K., L. Haak, A. Balcaen, E. Claeys, D. Demeyer and S. de Smet, 2004. Effect of linseed feeding at similar linoleic acid levels on the fatty acid composition of double-muscled Belgian blue young bulls. Meat Sci., 66: 307-315.
CrossRefPubMedDirect Link - Padre, R.D.G., J.A. Aricetti, F.B. Moreira, I.Y. Mizubuti and I.N. do Prado et al., 2006. Fatty acid profile and chemical composition of Longissimus muscle of bovine steers and bulls finished in pasture system. Meat Sci., 74: 242-248.
CrossRefPubMedDirect Link - Alfaia, C.P.M., S.P. Alves, S.I.V. Martins, A.S.H. Costa and C.M.G.A. Fontes et al., 2009. Effect of the feeding system on intramuscular fatty acids and conjugated linoleic acid isomers of beef cattle, with emphasis on their nutritional value and discriminatory ability. Food Chem., 114: 939-946.
CrossRefDirect Link - Adnoy, T., A. Haug, O. Sorheim, M.S. Thomassen, Z. Varszegi and L.O. Eik, 2005. Grazing on mountain pastures-does it affect meat quality in lambs? Livest. Prod. Sci., 94: 25-31.
CrossRefDirect Link - Lourenco, M., G. van Ranst, B. Vlaeminck, S. de Smet and V. Fievez, 2008. Influence of different dietary forages on the fatty acid composition of rumen digesta as well as ruminant meat and milk. Anim. Feed Sci. Technol., 145: 418-437.
CrossRefDirect Link - Priolo, A., D. Micola, J. Agabriela, S. Prachea and E. Dransfield, 2002. Effect of grass or concentrate feeding systems on lamb carcass and meat quality. Meat Sci., 62: 179-185.
CrossRefPubMedDirect Link - Scollan, N., J.F. Hocquette, K. Nuernberg, D. Dannenberger, I. Richardson and A. Moloney, 2006. Innovations in beef production systems that enhance the nutritional and health value of beef lipids and their relationship with meat quality. Meat Sci., 74: 17-33.
CrossRefPubMedDirect Link - Wood, J.D., R.I. Richardson, G.R. Nute, A.V. Fisher and M.M. Campo et al., 2004. Effects of fatty acids on meat quality: A review. Meat Sci., 66: 21-32.
CrossRefPubMedDirect Link - McKenna, D.R., P.D. Mies, B.E. Baird, K.D. Peiffer, J.W. Ellebracht and J.W. Savell, 2005. Biochemical and physical factors affecting discoloration characteristics of 19 bovine muscles. Meat Sci., 70: 665-682.
CrossRefPubMedDirect Link - Mancini, R.A. and M.C. Hunt, 2005. Current research in meat color. Meat Sci., 71: 100-121.
CrossRefPubMedDirect Link - Faustman, C. and R.G. Cassens, 1990. The biochemical basis for discoloration in fresh meat: A review. J. Muscle Foods, 1: 217-243.
CrossRefDirect Link - Vestergaard, M., M. Therkildsen, P. Henckel, L.R. Jensen, H.R. Andersen and K. Sejrsen, 2000. Influence of feeding intensity, grazing and finishing feeding on meat and eating quality of young bulls and the relationship between muscle fibre characteristics, fibre fragmentation and meat tenderness. Meat Sci., 54: 187-195.
CrossRefPubMedDirect Link