
K.M. Okukpe
Department of Animal Production, Faculty of Agriculture, University of Ilorin, P.M.B. 1515, Ilorin, Kwara State, Nigeria
A.A. Adeloye
Department of Animal Production, Faculty of Agriculture, University of Ilorin, P.M.B. 1515, Ilorin, Kwara State, Nigeria
M.B. Yousuf
Department of Animal Production, Faculty of Agriculture, University of Ilorin, P.M.B. 1515, Ilorin, Kwara State, Nigeria
O.I. Alli
Department of Animal Production, Faculty of Agriculture, University of Ilorin, P.M.B. 1515, Ilorin, Kwara State, Nigeria
M.A. Belewu
Department of Animal Production, Faculty of Agriculture, University of Ilorin, P.M.B. 1515, Ilorin, Kwara State, Nigeria
O.A. Adeyina
Department of Animal Production, Faculty of Agriculture, University of Ilorin, P.M.B. 1515, Ilorin, Kwara State, Nigeria
Asian Journal of Animal Sciences
Year: 2011 | Volume: 5 | Issue: 6 | Page No.: 365-372







ABSTRACT
The effect of Omega-3-fatty-acid on performance was carried out in sixteen (16) West African Dwarf does between 12 to 18 months of age with an average weight of 10 kg. Supplementation of Omega-3-fattyacid was varied from 0 mg for control to 500, 1000 and 1500 mg for low, medium and high level (s), respectively. Blood samples were collected on days 14, 28, 42 and 56 after Omega-3-fatty acid administration and were analyzed for haematological/biochemical parameters. Different levels of omega-3 fatty acid supplementation had no significant (p>0.05) effect on weight gain or feed efficiency of West Africa Dwarf goats. However, there was significant difference (p<0.05) in feed intake. West African Dwarf goats on 500 mg level of omega-3 fatty acid supplementation was observed to have relatively (p<0.05)the lowest feed efficiency and lowest daily weight gain. Results indicated no significant difference (p>0.05) between the mean values of serum total protein (50.50±6.45 to 43.25±6.45 g L-1) and that of serum cholesterol (3.08±0.43 to 2.93±0.43 mmol L-1) and a significant difference (p<0.05) between the mean values of serum urea level (4.45±0.73 to 6.38±0.73 mmol L-1). Although, there was no statistical difference between the mean values of serum total protein and serum cholesterol there was a tendency for a decrease at high (1500 mg) level of supplementation. There were no significant difference in their serum-glucose concentration, It was concluded that Omega-3-fatty acid can be used as dietary supplement for West African Dwarf does without posing any kind of stress on the health of the animal thereby increasing essential fatty acid in animal products.
PDF Abstract XML References Citation
Received: March 01, 2011;
Accepted: November 17, 2011;
Published: December 01, 2011
How to cite this article
K.M. Okukpe, A.A. Adeloye, M.B. Yousuf, O.I. Alli, M.A. Belewu and O.A. Adeyina, 2011. Physiological Response of West African Dwarf Goats to Oral Supplementation with Omega-3-fatty Acid. Asian Journal of Animal Sciences, 5: 365-372.
DOI: 10.3923/ajas.2011.365.372
URL: https://scialert.net/abstract/?doi=ajas.2011.365.372
DOI: 10.3923/ajas.2011.365.372
URL: https://scialert.net/abstract/?doi=ajas.2011.365.372
INTRODUCTION
The growth and reproductive performance of West African dwarf goat being critical to the socio-economic status of Nigerian rural dwellers could be improved in the area of nutrition and reproduction. We often hear the expression “We are what we eat” (Aworinde, 2010). However, a true expression would be we are what our animals eat. There are various researches that certain feed supplements increase n-3 fatty acids of animal products, including eggs, meat from major domestic species and milk from cows, sheep and goats (Chilliard and Ferlay, 2004; Correa, 2005; Baiomy, 2011).
Recently, it has been observed that feeding cows with supplemental fat cause improved energy balance and early oestrus-cycle due to the quality of fat supplement (Titi and Awad, 2007; Geiringer et al., 2010).
Fatty acids are the precursors of prostaglandins (PG) and steroid hormones (through cholesterol) which have an effect on fertility (Iribhogbe et al., 2011). In general, feeding supplemental fat such as calcium soaps of long chain fatty acids, fish meal and tallow increases conception rates. However, a lowered conception rate at first service has been reported when there was a paralleled increase in milk production (range of 2.2 to 4.5 kg d-1) (Geiringer et al., 2010). There are two main families of essential fatty acids, omega-3 and omega-6 fatty acids, that could affect fertility. The main source of omega-6 fatty acids is dietary linoleic acid (C18:2n-6) and this is converted to arachidonic acid (C20:4n-6), which inter alia is the precursor of the dienoic (2-series) PG, such as PGF2α. The same elongase and desaturaze enzymes also convert the main dietary omega-3 fatty acids (α-linolenic acid; C18:3n-3) to eicosapentaenoic acid (EPA; C20:5n-3), the precursor of the trienoic (3-series) PG, such as PGF3α (Da Cunha et al., 2007; Webb and O’Neil, 2008). Increasing the supply of omega-3 fatty acids will decrease production of dienoic PG and reduce competition for site of PG synthetase (Lammoglia et al., 2000; Davidson et al., 2007). In many cases the trienoic PG have lower biological activity than the corresponding dienoic PG (Calder and Deckelbaum, 2008) and this may directly affect aspects of fertility. For example, treatments that reduce ovarian and endometrial synthesis of PGF2α, at the expense of PGF3α, may contribute to a reduction in embryonic mortality (Mattos et al., 2000).
The essential fatty acid contained in omega-3-fatty acid are Eicosapentaenoic acid (EPA), Docosahexaenoic acid (DHA) and Alpha linoleic acid. These are not synthesized in the body of animals and humans and are necessary in nutrition because of its health benefits. Omega-3-fatty acid is used to cure different diseases such as diabetes, cancer, lung and kidney diseases, reproduction disorders, visceral disorders, metabolic disorders and rheumatoid arthritis (Chilliard and Ferlay, 2004; Babalola et al., 2008; Geiringer et al., 2010). Feeding supplemental fat has been reported not only to increase energy density but also to improve energetic efficiency depending on the fat quality (Lammoglia et al., 2000; Laaksonen et al., 2005; Kamara et al., 2011; Eghoghosoa et al., 2011). Fat and long-chain fatty acids, whether in adipose tissue or muscle, contribute to important aspects of meat quality and are central to the nutritional and sensory values of meat (Pambu-Gollah et al., 2000; Stewart, 2006; Webb and O‘Neil, 2008). Although, the mechanism by which fat supplementation alters reproductive performance is not yet understood, studies have been conducted to investigate the influence of dietary fat supplement on reproductive performance for cattle with few directed toward these effects on goats reproductive performance (Titi and Awad, 2007).
The objectives of this study therefore, was to determine the level(s) of omega-3-fatty acid that is suitable for effective performance of West African dwarf goats in relation to feed intake, body weight gain, serum metabolites such as protein, glucose, urea and cholesterol levels.
MATERIALS AND METHODS
The experimental work was carried out at the Small ruminant unit of the Animal pavilion of the Faculty of Agriculture, University of Ilorin between January to April, 2010.
Sixteen West African Dwarf goats between 12-18 months and weighing between 7-11 kg were used. They were quarantined for 18 days to acclimatize, during which they were treated against external/internal parasites and contagious caprine pleuropneumonia using Ivomectin injection (i/m) and Tissue Cultured Rinderpest Vaccine (TCRV), respectively. Two milliliter of Vitamin B-Complex was administered (i/m) to each goat to aid feed intake while 1 mL of long acting oxy-tetracycline injection was given to prevent pneumonia.
| Table 1: | Proximate composition of experimental diet |
 | |
Housing: The does were housed in pens with slatted floors and a little runs provided for exercise. The pens were cleaned with detergent and disinfected with Lysol before the arrival of the animals.
Feeding: Water and feed were supplied using plastic bowls and wooden feeding troughs, respectively in their pens and well secured to prevent feed wastage and water spillage. The animals were fed at 3% of their body weight with concentrate feed to meet their body requirement for growth and reproduction as shown in Table 1.
Experimental design: The does were randomly assigned to four treatment groups A (control), B (500 mg Omega-3-fatty acid), C (1000 mg Omega-3-fatty acid) and D (1500 kg Omega-3-fatty acid) (containing Docosahexaenoic acid (DHA) and Eicosapentaenoic acid (EPA)) with four replicates each. The goats were housed intensively and were checked for any palpable deformities of the reproductive organ. The experiment lasted for a period of eight weeks. Animals were weighed and blood, 3 mL for haematological analysis and 7 mL for biochemical analysis collected fortnightly. Serum total protein level(s) was analyzed using Biuret Method (Layne, 1957), Urea level(s) through Modified Berthelot Enzymatic Methods (Reinhold, 1953) and Cholesterol level(s) using CHOD-PAP (Lyophilized) kit (Zoppi and Fellini, 1976).
Statistical analysis: All data collected were subjected to a completely randomized design model and significant treatment means were compared using the Duncan multiple range test (Duncan, 1955; Steel and Torrie, 1990).
RESULTS
The chemical composition of the experimental diet is presented in Table 1 with feed having a dry matter content of 92.51% and crude protein of 18.81%. Table 2 shows the effects of different level of omega-3 fatty acids on feed intake, weight gain and feed efficiency of West African dwarf goats. Omega-3-fatty acid had no effect on the weight of the animal but seems to increase feed consumption, though not significantly (p>0.05). The control group (A) had the highest feed efficiency compared to B, C and D, although they were not significantly different (p>0.05).
There was no significant difference (p>0.05) between the mean values. The lowest serum urea value was in the control group (A) while the highest was in group D (Table 3). The mean value at high level of supplementation was statistically different (p<0.05) compared to the other treatments and the control.
| Table 2: | Effect of omega-3-fatty acids on feed intake, weight gain and feed efficiency |
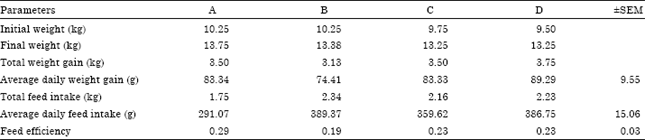 | |
| Table 3: | Effects of dietary omega 3- fatty acid intake on hematological and biochemical parameters of West African dwarf goats |
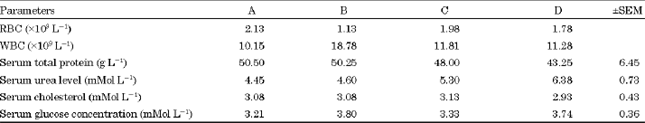 | |
| Means with different superscript are significantly different from each other (p<0.05). SEM = Standard Error of Mean | |
The mean serum cholesterol values ranged from 2.93±0.43 to 3.13±0.43 with group C having ±0.43 the highest while group D had the lowest, though not significant. The mean serum glucose concentration also ranged from 3.21±0.36 to 3.80±0.36 with the lowest found in group A (control) while group B had the highest, though not significant. All this did not follow a definite trend.
DISCUSSION
There was no significant difference in final body weight among the treatments, although animals in groups B and C had a marginal increase of 23% when compared with the control. In terms of weight gain group D (1500 mg Omega-3 fatty acid) had the highest but in terms of feed efficiency the control group was better than others. It could be adduced that Omega-3 fatty acid improves weight gain, though not significantly. Supplemental fat may improve energy balance and enhance follicular growth and development (Titi and Awad, 2007; Eghoghosoa et al., 2011). Further supported by Geiringer et al. (2010) that it was fatty acids and not the additional energy provided by the fatty acid that stimulated ovarian function and improve conception rate. This also improves the ability to use nutrients from food and convert them into muscle protein (Simopoulos, 2002). The improved health status of the animals could also be attributed to the Omega-3 fatty acid supplement as supported by various findings (Lammoglia et al., 2000; Leaf et al., 2003; Stewart, 2006). It has been shown that blood metabolites give an immediate indication of an animal’s nutritional status at a point in time (Pambu-Gollah et al., 2000). Altering fatty acid composition of foods by increasing the polyunsaturated to saturated fat ratio represents a useful method of disease prevention and health (Chilliard and Ferlay, 2004) Polyunsaturated fatty acid content of beef (Scollan et al., 2001) and polyunsaturated fatty acid content of lamb muscle (Demirel et al., 2004) has been increased by administration of polyunsaturated fatty acid. Omega-3-fatty acid concentration in plasma circulatory cells and all organs is dependent on the intake of preformed fatty acid (Chilliard and Ferlay, 2004). Although, the availability of this fatty acid to cells involves a series of physiological processes including digestion, absorption, transport and metabolism of fat (Kang, 2001; Cardozo et al., 2006).
The white blood cells seems to decrease with increasing supplementation, this shows that Omega-3 fatty acid has no negative effect on the animal but improves their health and performance (Lammoglia et al., 2000; Geiringer et al., 2010).
The non-significant difference in serum total protein with Omega-3-fatty acid supplementation is in agreement with what was observed in fishes fed dietary lipids (Babalola et al., 2008). Chilliard and Ferlay (2004) and Benchaar et al. (2007) indicated that the purpose for feeding dietary polyunsaturated fatty acid is to modulate milk fat and that altering all types of lipid supplement can increase goat milk fat without modifying yield of protein and serum protein which is incorporated in milk. So, Omega-3-fatty acid would not influence serum total protein anyway. Although, the mean values of serum total protein level showed no statistical significant difference it reduced as the omega-3- fatty acid supplementation increased. This trend could be as a result of increased protein metabolism since it has been hypothesized in rats that Omega-3-fatty acid may improve blood flow and consequently metabolism in cells (Wang et al., 2000; Laaksonen et al., 2005). Reduction in serum total protein could also be due to Omega-3- fatty acid oxidation. Castillejos et al. (2006) and Webb and O’Neil (2008) indicated that less saturated fats containing a number of fatty acids with double bond are easily oxidized either by direct chemical action or through intermediary activity of lipolytic enzymes.
The high serum urea level recorded for group D could be as a result of protein catabolism and excessive mobilization of muscle fat (Chimonyo et al., 2002) hence the low serum protein level. This supports the work of other researchers that high fat diets increase rumen ammonia and also plasma urea nitrogen (Chilliard and Ferlay, 2004; Tatsouka et al., 2008). This therefore, relates to the reduced level of serum protein at high level of supplementation. High urea level has been shown to be non toxic as was observed in this work (McIntosh et al., 2003).
Omega-3-fatty acid supplementation at the various levels had little or no effect on serum cholesterol level in West African Dwarf does. Although, there was no statistical difference between the mean values of serum cholesterol. There was a tendency for a decrease in serum cholesterol level at high level (1500 mg) of Omega-3-fatty acid supplementation which recorded the lowest value 2.93±0.43 mmol L-1 with a negligible difference in comparison with the value (3.08±0.43 mmol L-1) obtained for control. This in a way agrees with Mattos et al. (2000) who stated that omega-3-fatty acid has a major effect on plasma lipid by reducing triglycerol.
According to Webb and O’Neil (2008), the mechanism by which dietary fatty acid affects serum lipids and lipoprotein levels is unknown. In generally, it is assumed that serum lipid fatty acid composition reflects the fatty acid composition of a diet, which in turn affects serum cholesterol, triglyceride, lipoprotein and azoprotein. Thus the level of some free fatty acids in the serum can be influenced by the quantities in the diet but these changes are not dramatic. This could probably be the reason for the little effect of the dietary fatty acid (Omega-3-fatty acid) on serum cholesterol levels in this study.
Serum glucose also increase with increasing Omega-3 fatty acid supplementation supporting the report of various authors that it improves energy balance as well as increase energy density (Titi and Awad, 2007).
CONCLUSION
Omega-3-fatty acid supplementation fed at low, medium and high levels to West African Dwarf does showed little or no significant effect on weight gain, feed efficiency, serum cholesterol and total protein level but had a significant effect on serum urea levels. From the trend of the values obtained in this study there is a strong indication that supplementation beyond 1500 mg may produce a real effect. Omega-3-fatty acid can be used as a source of dietary fatty acid supplement in West African Dwarf does without posing any kind of stress on the health of the animal. This may provide a way to enrich animal products such as meat and milk with essential fatty acid required by humans.
REFERENCES
- Eghoghosoa, A.R., O.L. Aderemi, S.A. Olufemi and O.I. Peter, 2011. Effect of angiotensin I-converting enzyme inhibitor, captopril, on body weight and food and water consumption in oral contraceptive-treated rats. Am. J. Biochem. Mol. Biol., 1: 95-100.
CrossRefDirect Link - Baiomy, A.A., 2011. Influence of live yeast culture on milk production, composition and some blood metabolites of Ossimi Ewes during the milking period. Am. J. Biochem. Mol. Biol., 1: 158-167.
CrossRefDirect Link - Babalola, T.O.O., A.M. Adebayo, D.F. Apata and J.S. Omotosho, 2008. Effect of dietary alternative lipid sources on haematological parameters and serum constituents of Heterobranchus longifilis fingerlings. Trop. Anim. Health Prod., 41: 371-377.
CrossRef - Benchaar, C., H.V. Petit, R. Berthiaume, D.R. Ouellet, J. Chiquette and P.V. Chouinard, 2007. Effects of essential oils on digestion, ruminal fermentation, rumen microbial populations, milk production and milk composition in dairy cows fed alfalfa silage or corn silage. J. Dairy Sci., 90: 886-897.
CrossRefPubMedDirect Link - Cardozo, P.W., S. Calsamiglia, A. Ferret and C. Kamel, 2006. Effects of alfalfa extract, anise, capsicum and a mixture of cinnamaldehyde and eugenol on ruminal fermentation and protein degradation in beef heifers fed a high-concentrate diet. J. Anim. Sci., 84: 2801-2808.
CrossRefPubMedDirect Link - Castillejos, L., S. Calsamiglia and A. Ferret, 2006. Effect of essential oil active compounds on rumen microbial fermentation and nutrient flow in in vitro systems. J. Dairy Sci., 89: 2649-2658.
CrossRefDirect Link - Chilliard, Y. and A. Ferlay, 2004. Dietary lipids and forages interactions on cow and goat milk fatty acid composition and sensory properties. Reprod. Nutr. Dev., 44: 467-492.
CrossRefPubMedDirect Link - Chimonyo, M., N.T. Kusina, H. Hamudikuwana and I. Ncube, 2002. Changes in stress-related plasma metabolite concentrations in working Mashona cows on dietary supplementation. Livestock Prod. Sci., 73: 165-173.
CrossRef - Davidson, M.H., E.A. Stein, E.H. Bays, K.C. Maki and R.T. Doyle et al., 2007. Efficacy and tolerability of adding prescription ω-3 fatty acids 4g/d to simvastatin 40 mg/d in hypertriglycridemic patients: An 8-week, randomized, double-blind, placebo-controlled study. Clin. Ther., 29: 1354-1367.
PubMedDirect Link - Demirel, G., A.M. Wachira, L.A. Sinclair, R.G. Wilkinson, J.D. Wood and M. Enser, 2004. Effects of dietary n-3 polyunsaturated fatty acids, breed and dietary vitamin E on the fatty acids of lamb muscle, liver and adipose tissue. Br. J. Nutr., 91: 551-565.
CrossRefPubMedDirect Link - Iribhogbe, O.I., U. Akpamu, J.E. Emordi, A. Aigbiremolen, E.O. Nwoke and B. Idinoje, 2011. Antioxidants and electrolyte profile in early pregnancy: In vivo studies. Am. J. Biochem. Mol. Biol., 1: 82-88.
CrossRefDirect Link - Kamara, M.T., I. Amadou, Z. Kexue, M.B. Kelfala Foh and Z. Huiming, 2011. The influence of debittering and desalting on defatted foxtail millet (Setaria italica L.) protein hydrolysate. Am. J. Biochem. Mol. Biol., 1: 39-53.
CrossRefDirect Link - Laaksonen, D.E., K. Nyyssonen, L. Niskanen, T.H. Rissanen and J.T. Salonen, 2005. Prediction of cardiovascular mortality in middle-aged men by dietary and serum linoleic and polyunsaturated fatty acids. Arch. Internal Med., 165: 193-199.
PubMedDirect Link - Lammoglia, M.A., R.A. Bellows, E.E. Grings, J.W. Bergman and S.E. Bellows et al., 2000. Effects of dietary fat and sire breed on puberty, weight and reproductive traits of F1 beef heifers. J. Anim. Sci., 78: 2244-2252.
PubMedDirect Link - Layne, E., 1957. Spectrophotometric and turbidimetric methods for measuring proteins. Methods Enzymol., 3: 447-454.
CrossRefDirect Link - Mattos, R., C.R. Staples and W.W. Thatcher, 2000. Effects of dietary fatty acids on reproduction in ruminants. Rev. Reprod., 5: 38-45.
CrossRefDirect Link - McIntosh, F.M., P. Williams, R. Losa, R.J. Wallace, D. Beever and C.J. Newbold, 2003. Effects of essential oils on ruminal microorganisms and their protein metabolism. Appl. Environ. Microbiol., 69: 5011-5014.
CrossRefPubMedDirect Link - Pambu-Gollah, R., P.B. Cronje and N.H. Casey, 2000. status in free ranging indigenous goats. S. Afr. J. Anim. Sci., 30: 115-120.
Direct Link - Scollan, N.D., N.J. Choi, E. Kurt, A.V. Fisher, M. Enser and J.D. Wood, 2001. Manipulating the fatty acid composition of muscle and adipose tissue in beef cattle. Meat Sci., 85: 115-124.
CrossRefPubMedDirect Link - Simopoulos, A.P., 2002. Omega-3 fatty acids in inflammation and autoimmune diseases. J. Am. Coll. Nutr., 21: 499-505.
Direct Link - Tatsouka, N., K. Hara, K. Mlkuni, K. Hara, H. Hashimoto and H. Itabashi, 2008. Effects of the essential oil cyclodextrin complexes on ruminal methane production in vitro. Anim. Sci. J., 79: 68-75.
CrossRefDirect Link - Titi, H.H. and R. Awad, 2007. Effect of dietary fat supplementation on reproductive performance of goats. Anim, Reprod., 7: 23-30.
Direct Link - Wang, Y., T.A. McAllister, L.J. Yanke, Z. Xu, P.R. Cheeke and K.J. Cheng, 2000. In vitro effects of steroidal saponins from Yucca schidigera extract on rumen microbial protein synthesis and ruminal fermentation. J. Sci. Food Agric., 80: 2114-2122.
CrossRefDirect Link - Webb, E.C. and H.A. O'Neil, 2008. The animal fat paradox and meat quality. Meat Sci., 80: 28-36.
CrossRef