
S.R.E. Abo-Hegazy
Department of Agronomy, Faculty of Agriculture, Cairo University, Giza, Egypt
Noha F. El-Badawy
Plant Pathology Research Institute, ARC, Giza, Egypt
M.M. Mazen
Plant Pathology Research Institute, ARC, Giza, Egypt
H. Abd El-Menem
Plant Pathology Research Institute, ARC, Giza, Egypt
Asian Journal of Agricultural Research
Year: 2012 | Volume: 6 | Issue: 2 | Page No.: 60-72







ABSTRACT
Thirteen faba bean (Vicia faba L.) genotypes were assessed under greenhouse and field conditions in 2008/2009 and 2009/2010 seasons, using morphological and molecular characterization methods for resistance to chocolate spot diseases caused by Botrytis fabae A significant variation among the tested genotypes and their reaction to the disease was found under greenhouse condition. Sakha 1, 2 and Nubaria 1 were resistant, indicating by the lowest disease severity. Whereas, Giza 40, line 375 and Cairo 25 where susceptible. Other faba bean genotypes were moderately susceptible. RT-PCR analysis using chit 1 and 2 primers amplified fragment of chitinase gene (900 bp) in six faba bean genotype upon infection with the pathogen 48 h after inoculation. This fragment detected only in the resistance cultivars Sakha 1, 2 and Nubaria 1 and some moderate resistance genotype including Giza 3, line 24 H and line 36. At the same time no signal was detected in other infected genotypes or healthy ones. Field results of testing genotype against chocolate spot diseases often do not correspond completely with greenhouse results since the environment is less controlled and possibly natural infection could introduced unwanted pathogen(s) or other factors. The estimates of heritability in broad sense ranged from 0.90-0.99, high values of heritability were obtained for seed yield/plant (0.99), followed by plant dry weight, plant height, seeds pod-1 across the two seasons. However, No relationship was found between resistance of genotypes against B. fabae and their morphological characterizes under field conditions.
PDF Abstract XML References Citation
Received: January 11, 2012;
Accepted: March 22, 2012;
Published: May 15, 2012
How to cite this article
S.R.E. Abo-Hegazy, Noha F. El-Badawy, M.M. Mazen and H. Abd El-Menem, 2012. Evaluation of Some Faba Bean Genotypes against Chocolate Spot Disease Using cDNA Fragments of Chitinase Gene and Some Traditional Methods. Asian Journal of Agricultural Research, 6: 60-72.
DOI: 10.3923/ajar.2012.60.72
URL: https://scialert.net/abstract/?doi=ajar.2012.60.72
DOI: 10.3923/ajar.2012.60.72
URL: https://scialert.net/abstract/?doi=ajar.2012.60.72
INTRODUCTION
Faba bean (Vicia faba L.) is one of the most important food legumes in Egypt. The cultivated area of faba bean reached 244000 feddan with total production 207645 ton (FAO, 2009). It is widely cultivated for use in both human and animals in many parts of the world. The acreage’s and seed yields vary from season and location to another. This is attributed to various biotic and abiotic stresses. These stresses include disease, pests, less favorable environments etc.
The most important fungal disease is chocolate spot caused mainly by Botrytis fabae Sard. and to some extent, by B. cinerea Pers. ex. Fr. (Harrison, 1988; Rahman et al., 2002). Chocolate spot disease causes about 22-25% reduction of the seed yield in the northern region of the Nile Delta of Egypt (Khalil et al., 1993), where low temperature and high relative humidity, favoring its spread and prevailing (Mohamed, 1982). The fungus attacks leaves, stems, flowers and bodes.
Several attempts have been made to find out a ways to curb or minimize the impact of plant diseases on the yield. These includes breeding for disease resistance (Khalil et al., 1993; Zaki, 2010), fungicide control (Khaled et al., 1995), biological treatment (Mazen, 2004), plant extracts and agricultural practices (El-Sayed, 2005), induced resistance using biotic or abiotic agents to control Botrytis fabae was reported by Ismail et al. (2007). More recently, biotechnology has been used as a tool to increase field crop productivity in the contrast of sustainable agriculture (Tecson, 2002). Inoculation of plants with compatible and incompatible pathogens triggers a variety of plant defense responses, including the activation of genes encoding pathogenesis-related proteins (Van Loon, 1997). Many of these proteins have been shown to exhibit antifungal activity in vitro (Ponstein et al., 1994), for example the PR2-class (B-1, 3 glucanase) and PR3 class (chitinase) proteins hydrolyzing the important fungal cell wall components (Mauch et al., 1988). This suggested that PR proteins may play an important role in pathogen defense.
Plant disease resistance genes (R-genes) encode proteins that detect pathogens. R-genes have been used in resistance breeding programs for decades, with varying degrees of success. Recent molecular research on R-proteins and downstream signal transduction networks has provided exciting insights, which will enhance the use of R-genes for disease control. Definition of conserved structural motifs in R-proteins has facilitated the cloning of useful R-genes, including several that are functional in multiple crop species and/or provide resistance to relatively wide range of pathogens (Kirubakaran and Sakthivel, 2007).
R-gene-mediated resistance has several attractive features for disease control. When induced in a timely manner, the concerted responses can efficiently halt pathogen growth with minimal collateral damage to the plant (Palomineo et al., 2006).
This research aimed to evaluate thirteen faba bean genotypes, using morphological and molecular characterization methods for resistance to chocolate spot caused by Botrytis fabae to determine specific gene for resistance under greenhouse and field conditions.
MATERIALS AND METHODS
Field experiments: Thirteen faba bean (Vicia faba L.) genotypes were assessed in the growing seasons 2008/2009 and 2009/2010 seasons in Etay El-Baroad Research Station, El-Behaira Governorate, for Chocolate spot tolerance on the basis of the traits; plant dry weight (g), plant height (cm), No. of branches plant-1, pods plant-1, seeds plant-1, seeds pod-1, seed weight plant-1 (g), seed index (SI) and harvest index (HI).
Three faba bean inbred lines (Line 24 H, Line 36, 379) developed via single seed descent for more than seven generations were derived from Orobanche tolerant land races (Abdalla and Darwish, 1999). One line was derived from Cairo 375 (coded 375). Cairo 4, Cairo 5 and Cairo 25 a three new varieties (Abdalla and Darwish, 2008), Giza 40 susceptible Giza 3, Sakha 1 and 2 are resistant varieties to chocolate spot s diseases, Miser 1 a moderate resistance variety to chocolate spot disease and Nubaria 1 a highly resistant variety to chocolate spots disease.
Therefore, the research efforts are being focused on the identification of high yielding genotypes and high resistant to chocolate spot disease. The experiment was carried out as randomized complete block design with three replications. Each genotype was represented in each replicate by three ridges, 4 m long and 60 cm apart (plot size = 12 m2). Seeds were sown in double-seeds/hill, spaced 20 cm at one side of the ridge, plants were provided with normal irrigation. At harvest, ten guarded plants were randomly taken from each plot to measure yield and its components.
Inoculum preparation: The pathogen used in this investigation was isolated from naturally infected faba bean plants. The fungus was purified and identified according to Morgan (1971). Isolate was grown on Faba Bean Leaf Agar medium FBLA (Tivoli et al., 1986) (200 mL leaf extract, 30 g sucrose, 20 g sodium chloride and 20 g agar); the volume was adjusted to 1000 mL distilled water. Petri dishes were inoculated with B. fabae and incubated at 20±2°C for 12 days with 24 h fluorescent day’s length. Ten milliliter of distilled water were poured in each Petri dish. Spores were separated using a brush and filtered through cheese cloth. Spores were counted using a haemocytometer and calculated to number of spores mL-1, then diluted with sterilized distilled water to final concentration of (2.5x105 spore mL-1) and used immediately.
Seeds of faba bean genotypes were sown in sterilized pots contained disinfested soil; five seeds were sown in 30 cm-diameter pot representing one replicate. Three replicates of each genotype were assigned and kept under greenhouse conditions.
The plants were sprayed with spore suspension of Botrytis fabae (2.5x105 spores mL-1) at 45 days after sowing. All plants were covered with polyethylene bags for 24 h to maintain high relative humidity and then kept under greenhouse conditions for the following experiments.
Evaluation of disease severity (DS): Percentage of chocolate spot disease severity was assessed 24 and 72 h after inoculation with B. fabae was calculated using the scale 0-9 and formula adopted by Hanounik (1986):
Where:
| NPC | = | No. of plants in each class rate |
| CR | = | Class rate |
| NIP | = | No. of infected plants |
| MSC | = | Maximum severity class rate |
The response of genotypes was expressed as the DS values. Six resistance levels were used:
| • | HR (highly resistance), DS = ranging between 0 and 2.0 |
| • | R (resistant), DS = ranging between >2.0-15.0 |
| • | MR (moderately resistant), DS = ranging between >15.0-40.0 |
| • | MS (moderately susceptible), DS = ranging between >40.0-60.0 |
| • | S (susceptible), DS = ranging between >60.0-80.0 |
| • | HS (highly susceptible), DS = ranging between >80.0-100 |
Molecular studies
Isolation of total RNA from Vicia faba leaves: The protocol reported here is a modification of Chomczynski and Sacchi (1987), which based on guanidinium thiocyanate as an extraction buffer (4 M guanidinium thiocyanate, 100 mM HCl (pH 8.0), 75 mM EDTA and 2 (v/v)% β-mercaptoethanol) without phenol extraction step.
The frozen faba bean tissues were transferred to a pre-cooled mortar and homogenized into a fine powder. Fifty milliliter of the homogenizing buffer were added/5 g homogenized faba bean tissues and mixed well, then 5 mL of 2 M sodium acetate (pH 4.0) were added and mixed by inversion. An equal volume of chloroform-isoamyl alcohol (24-1) was added and shacked vigorously for 3 min and chilled on ice for 20 min. The solutions were transferred to autoclaved 50 mL centrifuge tube that has been pre cold on ice, centrifuged at 10,000 rpm for 25 min at 4°C. The RNA will be present in upper aqueous layer. DNA and protein are in the lower organic phase and inter phase. The aqueous phases were transferred to a fresh tube and the chloroform-isoamyl alcohol extraction step was repeated aqueous phases were transferred to a fresh tube and mixed with an equal volume of isopropanol, cold at -70°C for 30 min or at -20°C for 2 h. to precipitate RNA, centrifuge at 10,000 rpm for 25 min at 4°C. Supernatants were discarded. RNA present in the pellet. The pellets were dissolved in 5 mL of homogenizing buffer, gentle pipetting of the pellet may be required. An equal volume of isopropanol was added and mixed. The solutions were cold at -70°C for 15 min and were centrifuged at 10,000 rpm for 10 min at 4°C. The supernatants were removed from RNA pellet, wash with 75% ethanol. Dry under vacuum for 5 min and resuspended RNA in 1 mL DEPC- treated water. RNA was purified using tri-reagent RNA kit (Sigma, Lot No. T9424), then the purified RNA pellet was dissolved in 100 μL DEPC-treated water.
The extracted total RNA was electrophoresed on formamide formaldehyde agarose gel (1.2%) (50 mL 1X MOPS buffer, 2.7 mL formaldehyde 37% and 0.5 g agarose). Add 10X MOPS (3- morpholinopropane sulfonic acid) running buffer (800 mL DEPC treated water, 41.8 g MOPS, 16.6 mL sodium acetate and 20 mL 0.5 M EDTA, pH 8.0 and formaldehyde (12.2 M) to give a final concentration 1X and 0.66 M, respectively. Ethidium bromide was added to give final concentration 0.5 μg mL-1 before preparing the gel. The buffer reservoirs are filled with 1X MOPS running buffer, pH 7.0. After the samples were loaded (250 μL Formamide, 38 μL formaldehyde 37%, 50 μL 10X MOPS buffer and 0.01% Bromophenol blue) freshly prepared, the gel was run at 100 volts until the bromophenol blue has moved approximately half-way through the gel.
RT-PCR amplification
cDNA synthesis: cDNA synthesis was carried out in a reaction mixture (40 μL final volume) containing 10 μL of freshly prepared RNA as a template, 10 pmol of oligo (dT) anti-sense primer (Roche): 20 U of M-MuLV reverse transcriptase (Promega, USA), 100 mM of each dNTP; 1 mM DTT; 50 mM tris HCl. pH 8.3; 75 mM KCl and 6 mM MgCl2. The reaction mixture was incubated at 37°C for 1 h and stored at -20°C until used.
Chitinase primers were designed according to the published sequence clone pHS2, that was isolated from Ulmus americana (American elm) trees (Hajela et al., 1993). Accession No. L22032. The forward primer sequence (F1) was, 5'- ATT ATT GTT CTT TTA GTC CT -3' and the reverse primer sequence (R1) was, 5'- CTT TGT TCT TAT TCC ATT GA -3'.
PCR amplification: Induced chitinase gene, mRNA, was amplified by PCR, using T-GRADIENT thermal cycler from Biometra in 25 μL total volume containing 2.5 μL of cDNA; 25 pmol of forward primer (Chit 1) and reverse primer (Chit 2); 10 mM of each dNTPs; 1 U of Taq DNA polymerase; 10 mM tris-HCl, pH 9.0; 50 mM KCl; 1.5 mM MgCl2. The cDNAs were amplified as: Denaturation at 94°C for 3 min. followed by 35 cycles of 1 min at 94°C; 2 min at 45°C and 1 min at 72°C with a final extension step at 72°C for 7 min.
Agarose gel electrophoresis: Agarose gel electrophoresis was performed in DNA electrophoresis submarine mini cell. Agarose concentration was selected according to DNA size of expected PCR products and the electrophoresis was performed in 1 X TAE buffer (0.04 M Tris acetate, 0.001 EDTA, pH 8.0). DNA samples were mixed with 6 X gel loading dye (10 mM tris-HCl, pH 7.0,0.03% bromophenol blue, 0.03% xylene cynol FF, 60% glycerol, 60 mM EDTA). DNA was stained with ethidium bromide that was mixed with the gel and added both to the gel and to the buffer at a concentration of 0.5 g mL-1. DNA was visualized on gel documentation system (AAB: Advanced American Biotechnology 1166 E. Valencia Dr. Unit 6C, Fullerton CA 92631).
Statistical manipulations: The combined analysis of variance was carried out according to Steel and Torrie (1980), Broad sense heritability (h2) was estimated by using variance components method (Fehr, 1987):
The genotypic and phenotypic variances (δ2g and δ2ph) were calculated from the partitioning mean squares expectation as follows:
where, δ2e = MSe.
However, those over seasons are calculated from the pertinent mean squares expectation as follows:
where, δ2e= MSe.
Broad sense heritability (h2B) was calculated as follows:
RESULTS AND DISCUSSION
Evaluation of different faba bean genotypes to infection by chocolate spot disease under greenhouse: Data in Table 1 shows the reaction of thirteen different faba bean genotypes to chocolate spot disease, a significant variation among the tested genotypes and their reaction to the disease was found. Three faba bean genotypes showed a resistant against chocolate spot. These varieties were Nubaria 1, Sakha 1 and 2, respectively. The severity of chocolate spot on these plants were 6.6, 6.6 and 7.3% after 24 h and 14.1, 14.1 and 12.6% after 72 h. It was found that the severity of infection increased with increasing the incubation period from 24 to 72 h for all tested faba bean genotypes. Nevertheless, three faba bean genotypes out of thirteen were susceptible to chocolate spot disease these genotypes were Giza 40 followed by Line 375 and Cairo 25. Severity of infection were recorded 22.9, 21.5 and 20.7% after 24 h and 62.3 61.5 and 60.5% after 72 h after incubation period, respectively.
| Table 1: | Response of different faba bean genotypes to infection by chocolate spot disease under greenhouse |
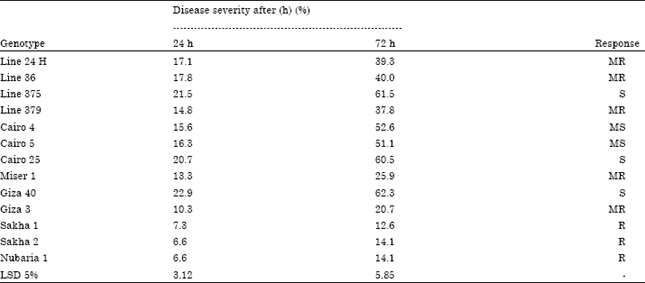 | |
| MS: Moderately susceptible, MR: Moderately resistant, S: Susceptible, R: Resistant | |
| Table 2: | Response of different faba bean genotypes to infection by chocolate spot disease under field condition |
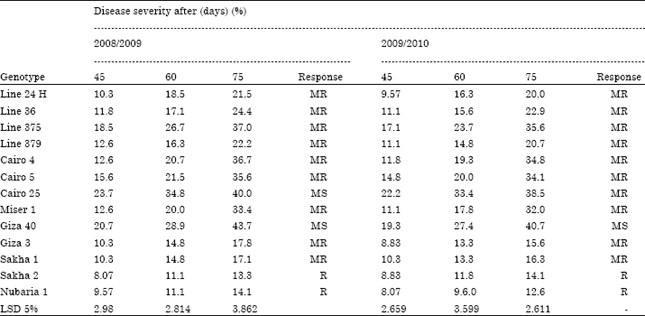 | |
| MS: Moderately susceptible, MR: Moderately resistant, S: Susceptible, R: Resistant | |
Other faba bean genotypes recorded moderately to chocolate spot disease. Its reaction to the disease ranged between 10.3 to 17.8% after 24 h and 20.7 to 40% after 72 h.
Evaluation of different faba bean genotypes to infection by chocolate spot disease under field conditions: Faba bean plants responded differently to chocolate spot infection under field conditions Table 2 during 2008/2009 growing season Sakha 2 and Nubaria 1 ranked as resistant genotypes judged by the lowest disease severity (13.3 and 14.1). Whereas, Giza 40 and Cairo 25 showed to be moderate susceptible to B. fabae having the highest degree of infection (43.7, 40), respectively compared to other tested genotypes. After 75 days. Other faba bean genotypes recorded as moderate resistant to fungal infection. For the second season (2009/2010), Nubaria 1 (12.6) and Sakha 2 (14.1) remained the resistant ones followed by Giza 3 (15.6) and Sakha 1(16.3). Whereas, Giza 40 (40.7) ranked as the moderate susceptible.
The obtained result showed slight differences in faba bean genotypes response to chocolate spot disease between greenhouse Table 1 and field experiments Table 2. For example Sakha 1 which recorded (R) under greenhouse turned to be (MR) under field condition, Cairo 4 and 5 (MS) in greenhouse become (MR) in the field, line 375, Cairo 25 and Giza 40 which scored (S) turned to be MR, MS and MS in the field, respectively. This finding can be explain in the light of the fact that field results often do not correspond completely with greenhouse results since the environment is less controlled and possibly natural infection could introduced unwanted pathogen(s) or other factors. Similarly, Omar (1984) found that 24 determinate and indeterminate faba bean varieties varied in their reactions to Botrytis fabae infection between indoor and outdoor experiments.
Mean performance
Agronomic characters: Table 3 illustrates that wide range of variability was recorded for some agronomic characters. The differences among the tested genotype were significant for plant dry weight, plant height, No. of branches plant-1, pods plant-1, seeds plant-1, seeds pod-1, seed weight plant-1, SI and HI.
Ranges for plant dry weight were from 101.7-157.2 g. with mean 135.9 g. Nubaria 1 recorded the highest plant dry weight genotype overall the two combined seasons. Meanwhile, the line 379 recorded the lowest plant dry weight genotype all over the two seasons. With respect to plant height, it ranged from 79.4-117.3 cm as a mean of the two seasons again Nubaria 1 recorded the highest one, number of branches plant-1 ranged from 3.2-6.3 as a mean of the two seasons, it is clear that the resistant and moderate resistant genotypes (Nubaria 1, Sakha 2, 1, Cairo 4, 5 and line 379, 375 and line 36 has the best agronomic characters. Nubaria 1 recorded the tallest genotype in both seasons.
| Table 3: | Mean performance for yield and yield component characters of 13 genotypes during 2008-2009 and 2009-2010 seasons (combined data) |
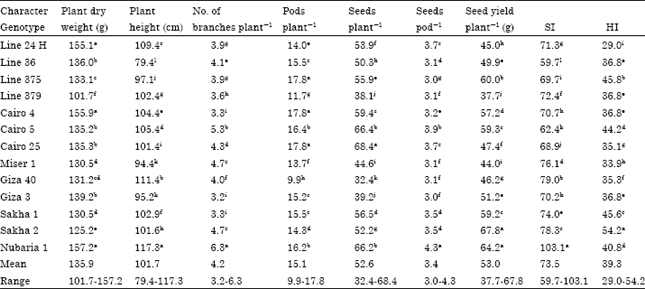 | |
| Means in each column followed by similar letters are not significantly different at 5% level, Values are means of three replications of 10 plants plot-1, SI: Seed index, HI: Harvest index | |
Yield characters: Also data in Table 3 shows a big range between the numbers of pods plant-1 all over the two seasons, were 9.9 to 17.8. Cairo 4, 25 and Line 375 produced greatest number of pods plant-1, followed by Cairo 5 and Nubaria 1 all over the two seasons. In the other hand, Giza 40 had the lowest number of pods plant-1.
The experiment showed that seed yield plant-1. distribution based on the means obtained all over the two seasons. It is obvious that the range 37.7-67.8 g, suggesting large variability in genes controlling seed yield plant-1.
With respect to seed index, it varied significantly. It was ranged from 59.7-103.1 g with mean 73.5 g all over the two seasons. The variety Nubaria 1 recorded the highest seed index. In contrast, Line 36 gave the lowest seed index in combined seasons.
Statistical parameters and heritability: The estimates of phenotypic variation (σ2ph) for seed yield plant-1 was high (283.13, followed by plant dry weight (218.55) across the two seasons, while it was low for seeds pod-1 (0.16). High values of genotypic variation (σ2g) were marked for seed yield plant-1 (281.0) followed by plant dry weight (216.04) in the combined two seasons. The magnitude of σ2g was low for seeds pod-1 (0.16) across the two seasons (Table 4).
Heritability in broad sense is defined as a ratio of genotypic to phenotypic variance. The estimates of heritability in broad sense (Table 4), ranged from 0.90 to 0.99 High values of heritability were obtained for seed yield plant-1 (0.99), followed by plant dry weight, plant height, seeds pod-1 and DSF 90 (0.98) across the two seasons and it was moderate for the others which were least affected by environmental changes and selection based on phenotypic performance could be possible. The higher magnitude of heritability indicated that these traits could be improved by direct selection. These results confirmed previous results of Sivasubramanian and Madhavamenon (1973), Alan and Hakan (2007) and Alghamdi and Ali (2004).
Correlation studies: Phenotypic correlation coefficients for all comparisons among the studied traits are presented in Table 5. Significant positive correlation values were detected between seed yield plant-1 (g) and all of traits except DSF 45, DSF 60 and DSF 90 were negative and high significant correlation coefficient.
| Table 4: | Estimates of coefficient of variation (CV), phenotypic (σ2ph), genotypic (σ2g), environmental (σ2e) variances, heritability (h2B) in broad sense for studied traits in 2008/2009, 2009/2010 and combined data |
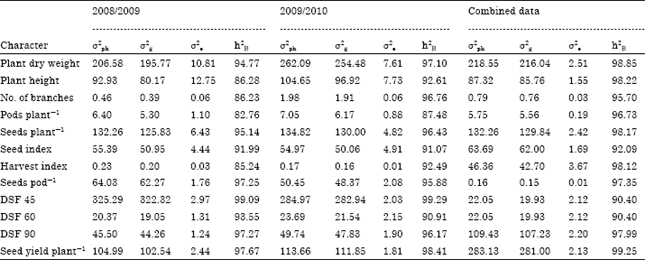 | |
| DSF (Disease severity after) | |
| Table 5: | Phenotypic correlation coefficients among studied traits of faba bean genotypes (Combined data) |
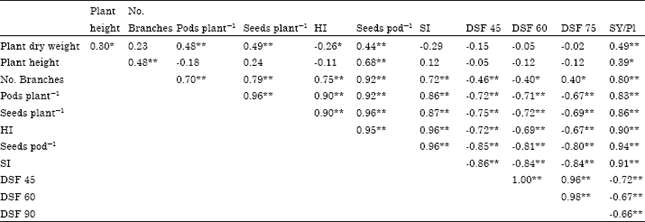 | |
| *’**Significant at 0.05 and 0.01 levels of probability, respectively | |
These findings indicate that selection for each trait would be accompanied by high yielding ability under such conditions. Negative and significant correlation coefficients were also obtained between disease severity after (45, 60 and 75 days) and each of pods plant-1 (-0.72**, -0.71** and -0.67**, respectively), Seeds pod-1 (-0.85**,-0.81** and -0.80**, respectively) and Harvest Index (-0.72**, -0.69** and -0.67**, respectively).
Thus the finding in the present study suggested two methods to select genotypes for specific traits to improve seed yield in faba bean. The first method is to select the genotype with high number of branches plant-1 which is directly responsible for number of seeds plant-1 based on the high positive correlation between the two traits as can be seen from Table 5. Therefore, Seed yield in faba bean can be improved by indirect selection of genotypes producing a large number of branches plant-1 coupled with high number of seeds plant-1 and negative association with disease severity. The second method for seed yield improvement in faba bean may be to select the genotypes with high number of pod plant-1 as this has indicated a highly significant and positive association with seed yield. These results are in agreement with those obtained by Alan and Hakan (2007) and Ulukan et al. (2003).
Molecular studies
Detection of chitinase gene by RT-PCR: Plant disease resistance genes (R genes) encode proteins have been used in resistance breeding programs for decades. Recent molecular research on R proteins and downstream signal transduction networks has provided exciting insights, which will enhance the use of R genes for disease control. The tools of molecular studies have been applied to investigate the mechanisms involved in disease resistance. First a standard inoculation procedure of Botrytis fabae on faba bean plants was established to obtain highly efficient and synchronized infection. Under the experimental conditions used, the first symptoms were visible 19-20 h. after inoculation as small necrotic brownish spots all over the leaf surface. The attention was focused on the plant genes expression during the early stages of infection (penetration). Materials were collected 8 h after inoculation when penetration was occurred (but no detectable symptoms were seen yet), 24 and 48 h after inoculation for analysis. Total RNA was isolated from healthy and infected faba bean leaves by using guanidinium thiocyanate as an extraction buffer without phenol extraction step as described under "Materials and Methods" and purified using Tri-reagent RNA extraction Kit (Sigma Lot No. T9424).
The integrity and quantity of purified RNA were confirmed by gel electrophoresis as shown in Fig. 1. The method was successful and showed the characteristic 2:1 ethidium bromide staining ratio of 28S to 18S ribosomal RNA indicating no significant RNA degradation.
The total RNA from thirteen samples that was previously infected with Botrytis fabae and thirteen samples of healthy plants were reverse transcribed using Reverse Transcriptase (RT) enzyme. PCR was performed on cDNAs synthesized on these mRNA samples using the forward primer Chit 1 and the Chit 2 as a reverse primer which were designed according to the published sequence clone pHS2, RT-PCR was used to amplify a fragment of about 900 bp from Chitinase gene in six genotypes of the infected plants after 48 h from inoculation which are Giza 3, Sakha 1, 2, Nubaria 1, Line 24 H and Line 36. While, no signal was detected in the negative control (healthy plants). The size of the PCR product was estimated by comprising its electrophoretic mobility with those of standard DNA marker as shown in Fig. 2, 3. On the other hand no signals were detected in the other genotypes in both infected and healthy plants.
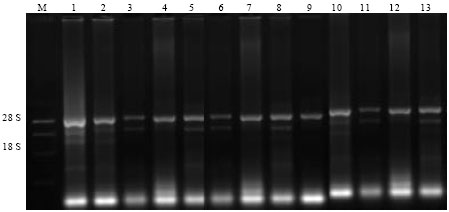 | |
| Fig. 1: | Total RNA extraction from infected faba bean tissues by B. fabae (Lanes and genotypes 1, TO 13), M. RNA marker (G319A, PROMIGA, USA). 1: Line 24 H, 2: Line 36, 3: Line 375, 4: Line 379, 5: Cairo 4, 6: Cairo 5, 7: Cairo 25, 8: Miser 1, 9: Giza 40, 10: Giza 3, 11: Sakha 1, 12: Sakha 2, 13: Nubaria 1 |
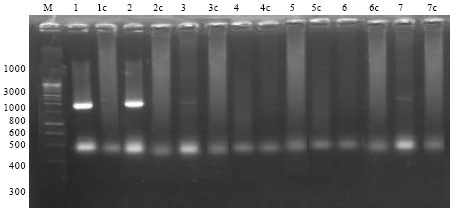 | |
| Fig. 2: | RT-PCR products, using Chit 1 and Chit 2 primers with infected faba bean genotypes (1 to 7) after inoculation period 48 h. and healthy faba bean tissues. M: 100 bp DNA ladder marker. Healthy genotypes (1c: Line 24 H, 2c: Line 36, 3c: Line 375, 4c: Line 379, 5c: Cairo 4, 6c: Cairo 5. 7c: Cairo 25). Infected genotypes (1: Line 24 H, 2: Line 36, 3: Line 375, 4: Line 379, 5: Cairo 4, 6: Cairo 5, 7: Cairo 25) |
 | |
| Fig. 3: | RT-PCR products, using Chit 1 and Chit 2 primers with infected faba bean genotypes (8-13) after inoculation period 48 h. and healthy faba bean tissues. M: 100 bp DNA ladder marker. Healthy genotypes (8c: Miser 1, 9c: Giza 40, 10c: Giza 3, 11c: Sakha 1, 12c: Sakha 2 13c: Nubaria 1). Infected genotypes (8: Miser 1, 9: Giza 40, 10: Giza 3, 11: Sakha 1, 12: Sakha 2, 13: Nubaria 1) |
Parasites must possess a machinery to cope with their hosts in order to obtain nutrients and overcome host defenses. This machinery is based on what is so called pathogenicity and virulence genes that are tightly regulated and generally expressed only when the parasite is growing in the host (Woloshuk and Kolattukudy, 1986; Peter and Verma, 1990). Plant pathogens usually express several virulence mechanisms that enhance their ability to colonize and damage host plant tissue, for example, the pathogen produces pectic enzymes that degrade the pectic fraction of the host plant cell wall, resulting in tissue disintegration and nutrient release that facilitate growth of the pathogen.
Plants are equipped with different mechanisms of resistance against pathogens; some of them are natural barriers such as cell wall, cuticle and the root pericycle (Keen, 1992). Another important active defense system in plants is called the Hypersensitive Response (HR). The expression of many plant genes is activated during the HR, including genes encoding enzymes of the phenolic pathway, peroxidases, glucanases and chitinases (Hahlbrock and Scheel, 1989; Dixon and Lamb, 1990).
The primer pair specific to chitinase gene was used in order to amplify a PCR product of about 900 bp from the sample tested which is in agreement with Eman (1994), Attia et al. (2007) and Kong et al. (2005) who constructed a cDNA library using mRNA isolated from wheat plants after inoculation with conidiospore suspension of Fusarium graminearum. They found that the cDNA clone encodes an acidic isoform of class 1 chitinase containing 960 bp coding region.
Our data strongly suggests that accumulation of the defense response gene transcripts is a general response of faba bean to infection by Botrytis fabae conidia. These data suggest that the spores of fungi produces a general elicitor(s) that is sensed by a general receptor(s) leading to defense response gene transcription, as elucidated by Kruger et al. (2003).
In spite of the extensive examination of the literature about the molecular markers associated with chocolate spot disease resistance genes in faba bean as a method in evaluation of new cultivars. No published data could be found to be dealt with this point. So this study is considered to be the first novel one to determine specific gene for resistance in new lines.
REFERENCES
- Abdalla, M.M.F. and D.S. Darwish, 1999. Breeding Faba Bean for Orobanche Tolerance using the Concept for Uniform Resistance. In: Advances in Parasitic Weed Control at On-Farm Level: Supra-Regional Project Ecology and Management of Parasitic Weeds. Vol. II. Joint Action to Control Orobanche in the WANA region, Kroschel, J., M. Abderabihi and H. Betz (Eds.)., Magraf Verlag, Weikersheim, Germany, pp: 205-213.
- Alan, O. and H. Geren, 2007. Evaluation of heritability and correlation for seed yield and yield components in faba bean (Vicia faba L.). J. Agron., 6: 484-487.
CrossRefDirect Link - Attia, M.F., N.M. Abou-Zeid, K.A. Abada, M.H. Soliman and N.F. El-Badawy, 2007. Isolation of chitinase gene induced during infection of Vicia faba by Botrytis fabae. Arab J. Biotech., 10: 289-300.
Direct Link - Chomczynski, P. and N. Sacchi, 1987. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem., 162: 156-159.
CrossRefPubMedDirect Link - Fehr, W.R., 1987. Principles of Cultivar Development. Vol. 1, Macmillan Publishing Co., New York.
Direct Link - Hahlbrock, K. and D. Scheel, 1989. Physiology and molecular biology of phenylpropanoid metabolism. Annu. Rev. Plant Physiol. Plant Mol. Biol., 40: 347-369.
CrossRefDirect Link - Harrison, J.G., 1988. The biology of Botrytis spp. on Vicia beans and chocolate spot disease-a review. Plant Pathol., 37: 168-201.
CrossRefDirect Link - Keen, N.T., 1992. The molecular biology of disease resistance. Plant Mol. Biol., 19: 109-122.
CrossRef - Khalil, S.A., M.M. El-Hady, R.F. Dissouky, M.I. Amer and S.A. Omar, 1993. Breeding for high yielding ability with improved level of resistance to chocolate spot (Botrytis fabae) disease in faba bean (Vicia faba L.). J. Agric. Sci. Mansoura Univ., 18: 1315-1328.
Direct Link - Kirubakaran, S.I. and N. Sakthivel, 2007. Cloning and overexpression of antifungal barley chitinase gene in Escherichia coli. Protein Exp. Purific., 52: 159-166.
CrossRef - Kruger, W.M., L.J. Szabo and R.J. Zeyen, 2003. Transcription of the defense response genes chitinase IIb, PAL and peroxidase is induced by the barley powdery mildew fungus and is only indirectly modulated by R genes. Phys. Mol. Plant Pathol., 63: 167-178.
Direct Link - Kong, L., J.M. Anderson and H.W. Ohm, 2005. Induction of wheat defense and stress-related genes in response to Fusarium graminearum. Genome, 48: 29-40.
PubMedDirect Link - Mauch, F., L.A. Hadwiger and T. Boller, 1988. Antifungal hydrolases in pea tissue: I. Purification and characterization of two chitinases and two beta-1, 3-glucanase differentially regulated during development and in response to fungal infection. Plant Physiol., 87: 325-333.
PubMedDirect Link - Morgan, D.J., 1971. Numerical toxonomic studies of the genus Botrytis: II. Other Botrytis taxa. Trans. Br. Mycol. Soc., 56: 327-335.
CrossRef - Palomineo, C., Z. Satovic, J.I. Cubero and A.M. Torres, 2006. Identification and characterization of NBS-LRR class resistance gene analogs in faba bean (Vicia faba L.) and chickpea (Cicer arietinum L.). Genome, 49: 1227-1237.
PubMedDirect Link - Rahman, M.Z., Y. Honda, S.Z. Islam and S. Arase, 2002. Effect of metabolic inhibitors on red light induced resistance of broad bean (Vicia faba L.) against Botrytis cinerea. J. Phytopathol., 150: 463-468.
CrossRefDirect Link - Steel, R.G.D. and J.H. Torrie, 1980. Principles and Procedures of Statistics: A Biometrical Approach. 2nd Edn., McGraw Hill Book Co., New York, USA., ISBN-13: 9780070609266, Pages: 633.
Direct Link - Ulukan, H., M. Guler and S. Keskin, 2003. A path coefficient analysis some yield and yield components in faba bean (Vicia faba L.) genotypes. Pak. J. Biol. Sci., 6: 1951-1955.
CrossRefDirect Link - Van Loon, L.C., 1997. Induced resistance in plants and the role of pathogenesis related proteins. Eur. J. Plant Pathol., 103: 753-765.
Direct Link - Woloshuk, C.P. and P.E. Kolattukudy, 1986. Mechanism by which contact with plant cuticle triggers cutinase gene expression in the spores of Fusarium solani f. sp. pisi. Proc. Natl. Acad. Sci. USA., 83: 1704-1708.
PubMedDirect Link
hamdy mohamed abo shedida Reply
This paper is useful l need all paper related this subject
Dr. Samir Abo Hegazy
Dear Dr. Hamdy
I am so sorry because i can't send it now for you, because i am not in Egypt now, may be after 2 monthes when i come back to Egypt, send you these papers
hamdy mohamed gamal eldeen aboshedida
dear dr samir/iam hamdy i requested from you the papers ralated this subject before and you tell me you will send them to me when you come back to egypt and i should be grateful to you if you send me these things