
R. G. Ahmed
Department of Zoology, Faculty of Science, Cairo University, Beni-SuefBranch, Beni-Suef, Egypt
International Journal of Zoological Research
Year: 2006 | Volume: 2 | Issue: 1 | Page No.: 48-59







ABSTRACT
Because of the temperature is one of the most encountered stressful factors in the environment, it was deemed important to survey the literature for reports on high temperatures or hyperthermia exposure durations at which biological effects occur. Since that time, several method of heating the entire body have evolved, including the artificial induction of fever, the wrapping of an anesthetized patient in plastic and dipping them in hot wax and heating the blood supply. The aim of this review was to determine the changes in tissue temperature and the duration of this effect. In general, the higher the temperature or the longer the hyperthermia, the greater the chance for observing a perturbation to the biological effects. It appears reasonably well established that, short exposure to sharply-elevated temperatures result in a protective effect against further thermal insult; the generation of heat shock proteins by cells coincides with the onset of such "thermal protection". It can be concluded that, thermal damage increases as the time at an elevated temperature increases.
PDF Abstract XML References
How to cite this article
R. G. Ahmed, 2006. The Relation between Biological Consequences and High Temperature in Mammals. International Journal of Zoological Research, 2: 48-59.
DOI: 10.3923/ijzr.2006.48.59
URL: https://scialert.net/abstract/?doi=ijzr.2006.48.59
DOI: 10.3923/ijzr.2006.48.59
URL: https://scialert.net/abstract/?doi=ijzr.2006.48.59
INTRODUCTION
In view to the recent studies, the biological effect of high temperatures or hyperthermia exposure durations are mediated through both thermal and nonthermal mechanisms of interaction. Both mechanisms are important and either may be the predominant mechanism depending on the exposure conditions. Also, the effects of hyperthermia are strongly influenced by thermotolerance (Lepock and Kruuv, 1980).
In addition, mitochondrial respiratory chain activity is central to all hypothesis of cellular heat generation-thermogenesis (Nicholls and Locke, 1983). There is an association between the relative number of mitochondria per cell and homothermic and heterothermic organisms; the former having more mitochondria and the mitochondria having a greater surface area (Giardier and Stock, 1983).
On the other hand, several reports are listed on the harmful effect of high temperatures and hyperthermia exposure (Poswillo et al., 1974; Johnson et al., 1976; Fajardo, 1984; Sminia et al., 1989; Sharma and Cervos-Navarro, 1990; Sharma et al., 1992; Bongiovanni et al., 1999; Lee et al., 2000; Kay and Marino, 2000; Hirobumi et al., 2002; Chou et al., 2003; Edwards et al., 2003; Radmilovich et al., 2003; Sharma et al., 2003; Chang et al., 2004). The following is a brief concerted attempt to describe the backgrounds and recent findings which provide specific suggestions for explore the effect of high temperatures or hyperthermia exposure duration.
Effects on the Life Processes
There are a number of fascinating observations of living organisms maintained under a wide variety of temperatures. For example, in vitro mammalian cells retain their viability for nearly indefinite periods if kept in the vapor of liquid nitrogen (-79°C), will die if frozen at 0°C, have cell growth kinetics which appear maximal at 33-39°C and are generally lethally affected by temperatures of about 45°C; however, a very brief exposure to a very high temperature can confer "thermotolerance" and thus render the cell capable of surviving subsequent exposures to high temperature (Miller and Ziskin, 1989).
The temperature is not constant throughout the body. Core temperature is usually 37°C, but skin temperatures can range from a low of 31.5°C (calf) to 35.0°C (Olesen, 1984). During work a person's core temperature can rise to 38.4°C (Gas and Camp, 1984); a competitive marathon runner's temperature was 41°C (Dark and Edholm, 1985). During sunbathing, a person's skin can rise about 5°C on the 'sunny' side (Clark and Edholm, 1985), with dissipating mechanisms preventing further increase in temperature. Miller and Ziskin (1989) found that ultrasonically-induced heat is whether maternal temperature per se or fetal temperature specifically is the important parameter in attempting to establish some above-normal temperature below which no thermal effects are anticipated.
Also, there is a range of temperatures over which the body can function. A person's temperature can vary with type and duration of activity. DuBios (1948) has reported an estimate of the range of body temperature (rectal, oral) for various activities, from early morning (sleeping, nonactivity) to normal ranges to vigorous exercise and hard work. The temperatures range from slightly less than 36°C to about 40°C, with the usual range of normal roughly 36 to 38°C (Hardy, 1982).
Teratogenic Effects in Mammals
Hyperthermia is thought to be a teratogen in many animal species including primates and also in humans (Smith, 1982; Shepard, 1982; Sasaki et al., 1995). The retrospecture human studies have a related hyperthermia to both neural tube and head defects (Ladye et al., 1980; Fisher and Smith, 1981; Spragett and Fraser, 1982). Furthermore, Sasaki et al. (1995) reported that hyperthermia caused by sauna, hot tub, or fever during the early stages of pregnancy is related to an increased risk for neural tube defects. Also, Edward (1986) found that, the hyperthermia causes several congenital abnormalities in a number of domestic and laboratory species of mammals, including defects on the central nervous system.
Layde et al. (1980) proposed that hyperthermia in the pregnant woman is associated with neural tube defects in her offspring. Also, maternal hyperthermia in early pregnancy can cause neural tube defects in man, especially anencephaly (Shiota, 1982). Milunsky et al. (1992) found that exposure to heat in the form of hot tub, sauna, or fever in the first trimester of woman pregnancy was associated with an increased risk for neural tube defects (NTDs). Moreover, Edwards et al. (2003) found that hyperthermia during pregnancy can cause embryonic death, abortion, growth retardation and developmental defects. Furthermore, Arora et al. (1979) noticed that oedema, microencephaly and microphthalmia on day 4, 6, or 8 when rats exposed to ambient temperatures of 43-44°C at various stages of pregnancy. Berman et al. (1990) found that increasing ambient temperature was effective in decreasing maternal weight gain and fetal body weight and increasing fetal relative brain weight. In humans, epidemiological studies suggest that an elevation of maternal body temperature by 2°C for at least 24h during fever can cause a range of developmental defects, but there is little information on thresholds for shorter exposures (Edwards et al., 2003). Also, Cronje (1977) noted that pregnant women with elevated intrauterine temperatures (38.8 vs 37.4°C for controls) had a higher incidence of fetal tachycardia. Laurence et al. (1968) reported an excess of first trimester pyrexia in mothers delivering fetuses with anencephaly, memmigomyelocoele, or hydrocephalus. Fraser and Skelton (1978) recorded that possible teratogenicity of maternal fever were present. In limited study with pregnant baboons whose core temperature was raised to 42°C (39°C is normal) over 3 to 4 h, a variety of fetal disorders; metabolic acidosis, fetal cardiac arrest, fetal asphyxiation, rise in heart beat, fall in blood pressure, including abortion were noted (Morishima et al., 1975).
Exposure of embryos to experimentally elevated temperature during organogenesis has long been known to be embryotoxic (Johnson et al., 1975; Hutchinson and Bowler, 1984; Edwards, 1986; Upfold et al., 1989). Hyperthermia at critical stages during embryonic development causes several developmental abnormalities (Edwards, 1986) including defects of the central nervous system. Experiments carried out on rat embryos cultured for 48 h at 40.5°C resulted in a significant microcephaly and oedema of the pericardium (Cockroft and New, 1978). From these observations, we can conclude that, the temperature may delay partially the development of the embryos.
The range of defects induced by hyperthermia in experimental animals includes: anencephaly/ exencephaly, encephalocele (Webster and Edwards, 1984; Cawdell-Smith et al., 1992), micrencephaly (Edwards, 1969; Edwards et al., 1984; Upfold et al., 1989), microphthalmia, talipes, arthrogryposis, abdominal wall defects and limb reduction defects (Edwards, 1986). Such defects have been induced by heat in a variety of mammals, including guinea pigs, hamsters, rats, mice, rabbits, sheep, pigs, monkeys and humans (Edwards, 1986), though the confounding effects of the febrile illnesses themselves and their therapies remain problematic in the interpretation of the human data.
Furthermore, the central nervous system defects appear to be the most common consequence of hyperthermia in all species and cell death or delay in proliferation of neuroblasts is believed to be one major explanation for these effects (Edwards et al., 1974; Wanner et al., 1975; Upfold et al., 1989). It is apparent from the above mentioned results that, the hyperthermia or temperature not only may a potential behavioral teratogen but also may obstruct the biological processes.
On the other hand, the signs of Malignant Hyperthermia (MH) are muscle spasm, rapid increase in temperature, tachycardia, increased blood pressure, increased oxygen demand, increased arterial PaCO2 and acidosis (Harley et al., 2005). This usually occurs on induction but its onset is, on rare occasions, delayed by a few hours. In generally, heatstroke is a multisystem disorder that can result in death (Flanagan et al., 1995). So, from the pre-said studies, the temperature may cause some malformation in the pregnant, which may retard or delay partially the growth of its fetus and reduce its biological functions.
Effect of Temperature on the
Protein Synthesis
Regarding to the Protein synthesis, the effect of hyperthermia, in general, on whole body leads to temporary cessation of normal synthesis of it (German, 1984). Also, Edwards et al. (1974) mentioned that heat causes protein denaturation in newborn guinea pigs. Heat causes a transient depression in overall protein synthesis (Lindquist and Craig, 1988). Moreover, Edwards et al. (1997) said that there was a reduction in normal protein synthesis of mammalian brain as a result of heat shock. The degradation of protein increased in rat brain as a result of hyperthermia (Bongiovanni et al., 1999). According to the above results, it is worth mentioning that, the present overview may suggest the deleterious effect of temperature on the vital processes of the biological systems.
Metabolic Processes
The responses to heat are moderated by factors which influence on the metabolic heat production/ heat loss, including the severity and duration of heat stimuli, accompanying exercise, the magnitude of the metabolic response and individual characteristics such as body composition, age and gender. Kay and Marino (2000) said there were alterations in the metabolic process as a result of exercise heat stress. Moreover, Wilmore et al. (1975) observed that the higher environmental temperature decreased metabolic rate in patients with large thermal injuries in whom the decrement in dry heat loss produced by higher ambient temperature exceeded the increase of wet heat loss. In other instance, Nilsen (1984) recorded that the hyperthermia distorts the small vessels and also produces irregularities of the microvascular pattern. Therefore, the above results presumed that, the distortion in blood vessels may cause the impairment of the metabolic processes (carbohydrates genesis) and thus, may reflex some delaying in the growth of the cellular processes.
Thermal Tolerance and Thermal Sensitivity
There is no known mechanism for explaining sensitivity of cells to heat (Miller and Ziskin, 1989). A variety of effects is known, including enhanced sensitivity of cells during stages S, G2 and M, perturbations to the cytoskeletal system, modulation of polyamine release and changes in DNA synthesis in hyperthermia treated cells (Leeper, 1985).
In addition, cells appear to be capable of developing nonheritable tolerance to heat treatments. Maximal tolerance develops during 3 to 4 h of exposure of cells to temperature below 42.5°C followed by 8 to 10 h of exposure of cells to greater than 43°C and then a return to 37°C (Leeper, 1985). The thermotolerance has been observed to occur in nearly every living biological system, including protozoan, slime molds, fungi, molluscs, nematodes, insects, echinoderms, fish, amphibian, birds, mammals and plants (Mover and Sharf, 1984). A notable exception to this appears to be the very early embryo stages (where only 8 or so cells are involved); it is possible that such cells are thermotolerance capable, but because there are so few of them it is difficult to demonstrate (Heikkila et al., 1985). Later embryonic stages (e.g., blastula), which have many more cells, have been shown to be capable of induction of thermotolerance (Heikkila et al., 1985). Thus, thermotolerance increases the resistance of surviving cells by several orders of magnitude, particularly if the heat fractions are given daily (Miller and Ziskin, 1989).
Thermal Protection
Pertaining to a genetic basis, the thermotolerance can be found in the development of Heat Shock Proteins (HSPs) (Hahnel et al., 1986). Stressful stimuli activate the heat shock (stress) response in which a set of heat shock proteins is induced, which play roles in cellular repair and protective mechanisms (Bechtold and Brown, 2000). Also, the induction of stress response Heat Shock Proteins (HSPs) is a highly conserved response that protects many cell types from diverse physiological and environmental stressors (Kelly, 2002). There was an increase in the synthesis of a small set of proteins known as the heat shock proteins due to heat exposure (Lindquist and Craig, 1988). Also, the HSP72 synthesis significantly increased in the brain of the rats with hyperthermic treatment (Yang et al., 1994). Recovering embryos mounted a heat shock response as evidenced by the induction of a 71 kilodalton heat shock protein (Walsh et al., 1987). Activation of the heat shock response was not a teratogenic event in the developing embryo. Heat acclimation elevates the levels of HSP (Malyshev et al., 2000). Moreover, when mouse exposed to environmental stress, cell survival is supported by the upregulation of stress proteins such as heat shock proteins or Glucose Regulated Proteins (GRPs), which help prevent protein denaturation (Ostberg et al., 2002). Yang and Lin (1999) revealed that, the neuronal HSP72 increases survival in rats exposed to heat stroke by attenuating arterial hypotension. The susceptibility of large neurons to stress induced cell death could be due, in part, to their inability to synthesize rapidly HSP70 in sufficient amounts to protect these cells from the initial molecular consequences of stress (Morrison-Bogorad et al., 1994). Based on the above described results, the hyperthermia or heat may induce the secretion of the heat shock protein to make the protective mechanisms but this secretion depends not only on the duration of exposure but also on the type of tissue and the environmental conditions. Actually, it is not known if these proteins are truly protective, or if they are merely products of the process that provide the thermal tolerance (Miller and Ziskin, 1989).
Temperatures and Lethality
A general relationship between time and temperature for thermal death is given by Dickson and Calderwood (1980) in vivo, in vitro and some clinical studies. An important point in Dickson's presentation is that below about 40°C there was virtually no effect of temperature on organisms, but above that point less exposure duration is required with rise in temperature to cause an effect. Heat induced cell lethality has been shown to be dependent on both temperature and heating time (Haveman and Hart, 1989). Also, Dunn et al. (2004) reported that, in the zooxanthellae, both apoptosis-like and necrosis-like activity increased throughout the duration of the exposure to heat stress (6 days), dependent on temperature dose. Heat induced cell death by apoptosis is a feature of teratogenic damage to the developing brain (Edwards et al., 1997). Apoptosis could be a by-product of a damaging heat exposure because of a priority favoring induction of the heat shock response over the normal gene program for organogenesis, survival being achieved at the expense of normal development (Edwards et al., 1997). This effect depends on the age of the animals and their prior thermal experiences. In general, thresholds and exposure-response relationships vary between species and even between different strains of the same species, depending on genotypes. But, a thermal stress, which by itself is nonlethal (Kahraman and Thach, 2004).
Effect Temperature on the Balance Between Heat Gain and Heat Loss
The rise in deep body temperature of the rats during work is proportional to work intensity and that the enhanced heat production capacity can be compensated for by increasing the heat loss activities (Harri et al., 1982). Also, Shih et al. (1984) found at Ta = 40°C, heat gain exceeded heat loss and led to hyperthermia and heat stroke and the latency for the onset of heat stroke was found to be around 87 minutes. At the onset of heat stroke, the comatose animals showed higher levels of rectal temperature, ear skin blood flow, respiratory evaporative heat loss, metabolic rate, intracranial pressure (ICP) and cerebral water content as compared to those of control animals (kept at an ambient temperature of 24°C). Therefore, it can be inferred that, the temperature may cause some disturbance in the balance between the heat gain and heat loss. Furthermore, Hyperhydration or increasing body water content above normal (euhydration) level was thought to have some benefit during exercise heat-stress; however, attempts to overdrink have been minimized by a rapid diuretic response (Latzka and Sawka, 2000).
Other Effects of Hyperthermia
Normal temperature maintenance requires an intact autonomic nervous system (Landsberg and Young, 1983). The body has adaptive capabilities for maintaining homeothermic condition; when the body temperature begins to rise, heat dissipating mechanisms come into play the peripheral blood vessels dilate, cardiac pulse rate increases (Gross et al., 1986) and sweating occurs. When the body temperature begins to drop "below normal" then shivering occurs, a mechanism designed to increase heat generation. On the other hand, the environmental stress as hyperthermia is generally believed to modify autonomic function through well-known classical pathways involving alternation in the cardiovascular function (Black et al., 1976). These changes in the cardiovascular function vary in severity and direction depending upon the degree of temperature change as well as the duration of the exposure (Tveita et al., 1991). Elazar et al. (1981) found that, the hyperthermia reduced the activity of the enkephalinergic system in the brain of rat; this reduced due to acceleration of the activity of peptidases involved in its breakdown. Furthermore, Arieli et al. (2003) reported that, the long term of heat acclimation prolongs the time to the development of the oxygen toxicity in the Central Nervous System (CNS-OT). Furthermore, Mirochnitchenko et al. (1995) suggested that, the hyperthermia may increase oxidative stress in tissues to form the reactive oxygen species with harmful to cellular functions and Chang et al. (2004) found that, the heatstroke increased the hydroxyl radicals and striatal neuronal damage in the brain of rats.
Summary of the Biological Effects of Hyperthermia
The elevated temperatures described in this report produce a wide range of significant biological effects as shown below:
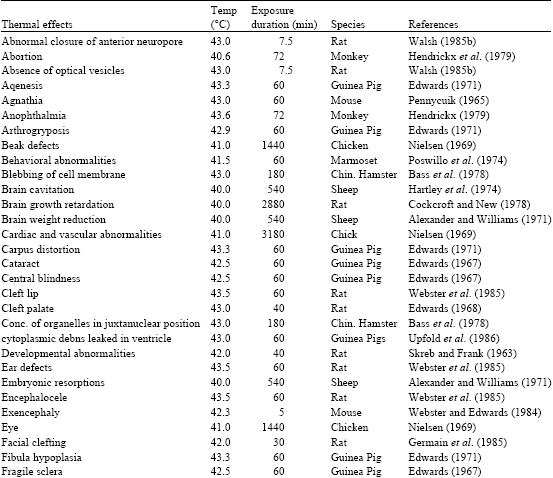 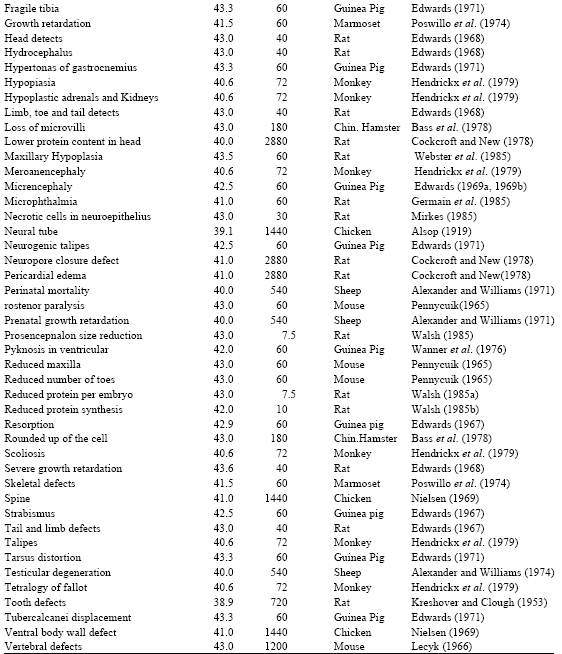 |
Finally, in view to the recent studies on the effects of heat and hyperthermia on the biological consequences, the current overview throw up important findings, the biological consequences are extremely sensitive to the heat and hyperthermia exposure; this may lead to some harmful effects on them.
REFERENCES
- Black, P.R., S.V. Devanter and L.H. Cohn, 1976. Effects of hypothermia on systemic and organ system metabolism and function. J. Surg. Res., 20: 49-63.
PubMed - Edwards, M.J., D.A. Walsh and Z. Li, 1997. Hyperthermia teratogenesis and the heat shock response in mammalian embryos in culture. Intl. J. Dev. Biol., 41: 345-358.
Direct Link - Edwards, M.J., R.D. Saunders and K. Shiota, 2003. Effects of heat on embryos and foetuses. Int. J. Hyperthermia, 19: 295-324.
Direct Link - Bass, H., J.L. Moore and W.T. Coakley, 1978. Lethality in mammalian cells due to hyperthermia under oxic and hypoxic conditions. Intl. J. Radiat. Biol., 33: 57-67.
PubMed - Fisher, N.L. and D.W. Smith, 1981. Occipital encephalocele and early gestational hyperthermia. Pediatrics, 68: 480-483.
Direct Link - Alexander, G. and D. Williams, 1971. Heat stress and development of the conceptus in domestic sheep. J. Agric. Sci. Camb., 76: 53-72.
CrossRef - Arora, K.L., B.J. Cohen and A.R. Beaudoin, 1979. Fetal and placental responses to artificially induced hyperthermia in rats. Teratology, 19: 251-259.
PubMed - Flanagan, S.W., A.J. Ryan, C.V. Gisolfi and P.L. Moseley, 1995. Tissue specific HSP70 response in animals undergoing heat stress. Am. J. Physiol., 268: 28-32.
Direct Link - Arieli, Y., M. Eynan, H. Gancz, R. Arieli and Y. Kashi, 2003. Heat acclimation prolongs the time to central nervous system oxygen toxicity in the rat possible involvement of HSP72. Brain Res., 962: 15-20.
CrossRef - Berman, E., D. House and H.B. Carter, 1990. Effect of ambient temperature and running wheel activity on the outcome of pregnancy in CD 1 mice. Teratog Carcinog Mutagen., 10: 11-20.
PubMed - Bongiovanni, G., S. Fissolo, H.S. Barra and M.E. Hallak, 1999. Posttranslational arginylation of soluble rat brain proteins after whole body hyperthermia. J. Neurosci. Res., 56: 85-92.
PubMed - Chou, Y.T., S.T. Lai, C.C. Lee and M.T. Lin, 2003. Hypothermia attenuates circulatory shock and cerebral ischaemia in experimental heatstroke. Shock, 19: 388-393.
PubMed - Bechtold, D.A. and I.R. Brown, 2000. Heat shock proteins Hsp27 and Hsp32 localize to synaptic sites in the rat cerebellum following hyperthermia. Mol. Brain Res., 75: 309-320.
CrossRef - Gross, D., A.R. Williams, C.W. Mann, F. McCord and D.L. Miller, 1986. Thermal and heart rate response to ultrasonic exposure in the second and third trimester dog fetus. J. Ultrasound Med., 5: 507-513.
Direct Link - Chang, C.K., W.T. Chiu, C.P. Chang and M.T. Lin, 2004. Effect of hypervolaemic haemodilution on cerebral glutamate, glycerol, lactate and free radicals in heatstroke rats. Clin. Sci., 106: 501-509.
Direct Link - Hendrickx, A.G., G.W. Stone, R.V. Hendrickson and K. Matayoshi, 1979. Teratogenic effects of hyperthermia in the bonnet monkey (macaca radiata). Teratology, 19: 177-182.
PubMed - Cockcroft, D.L. and D.A.T. New, 1978. Abnormalities induced in cultured rat embryos by hyperthermia. Teratology, 17: 277-283.
PubMed - Dickson, J.A. and S.K. Calderwood, 1980. Temperature Range and Selective Sensitivity of Tumors to Hyperthermia a Critical Review. In: Thermal Characteristic of Tumors Applications in Detection Treatment, Jain, R.K. and P.M. Gullino (Eds.). Annals of the New York Academy of Science, New York, pp: 180-205.
- Edwards, M.J., 1967. Congenital defects in guinea pigs following induced hyperthermia during gestation. Arch. Pathol., 84: 42-48.
PubMed - Harri, M., P. Kuusela and R.O. Rossi, 1982. Temperature responses of rats to treadmill exercise and the effect of thermoregulatory capacity. Acta Physiol Scand., 115: 79-84.
PubMed - Edwards, M.J., 1968. Congenital malformations in the rat following in duced hyperthermia during gestation.c Teratology, 1: 173-177.
PubMed - Hutchinson, R. and K. Bowler, 1984. The effect of hyperthermia in the development of the brain in the guinea pig. Brain Res., 14: 219-227.
PubMed - Edwards, M.J., 1969. Congenital defects in guinea pigs prenatal retardation of brain growth of guinea pigs following hyperthermia during gestation. Teratology, 2: 329-336.
PubMed - Hahnel, A.C., D.J. Gifford, J.J. Heikkila and G.A. Schultz, 1986. Expression of the major heat shock protein (hsp 70) family during early mouse embryo development. Teratog. Carcinog. Mutagen., 6: 493-510.
PubMed - Haveman, J. and A.A.M. Hart, 1989. The relationship between treatment duration and temperature for hyperthermia induced lethality of cultured murine cells. Eur. J. Cancer Clin. Onco., 25: 1629-1635.
Direct Link - Edwards, M.J., 1971. The experimental production of arthrogryposis multiplex congenita in guinea pigs by maternal hyperthermia during gestation. J. Pathol., 104: 221-229.
PubMed - Edwards, M.J., R.C. Mulley, S. Ring and R.A. Wanner, 1974. Mitotic cell death and delay of mitotic activity in guinea pig embryos following brief maternal hyperthermia. J. Embryol. Exp. Morph., 32: 593-602.
Direct Link - Elazar, Z., E. Motles and R. Simantov, 1981. Influence of body temperature on the epileptogenic effect of enkephalin microinjections into the hippocampus. Electroencephalogr. Clinical Neurophysiol., 52: 640-646.
PubMed - Johnson, K.M., J.G. Lyie, M.J. Edwards and R.H.C. Penny, 1976. Effect of prenatal heat stress on brain growth and serial discrimination reversal learning in the guinea pig. Brain Res. Bull., 1: 133-150.
PubMed - Kreshover, S.J. and O.W. Clough, 1953. Prenatal influences on tooth development I artificially induced fever in rats. J. Dent. Res., 32: 565-577.
CrossRef - Kay, D. and F.E. Marino, 2000. Fluid ingestion and exercise hyperthermia implications for performance thermoregulation metabolism and the development of fatigue. J. Sports Sci., 18: 71-82.
Direct Link - Kelly, K.J., 2002. Stress response proteins and renal ischemia. Minerva Urol Nefrol., 54: 81-91.
Direct Link - Kahraman, L. and B.T. Thach, 2004. Inhibitory effects of hyperthermia on mechanisms involved in autoresuscitation from hypoxic apnea in mice a model for thermal stress causing SIDS. J. Appl. Physiol., 97: 669-674.
Direct Link - Laurence, K.M., C.O. Carter and P.A. David, 1968. Major central nervous system malformations in South Wales II Pregnancy tactors, seasonal variation and social class effects. Br. J. Prev. Soc. Med., 22: 212-222.
CrossRef - Lepock, J.R. and J. Kruuv, 1980. Thermotolerance as a possible cause of the critical temperature at 43°C in mammalian cells. Cancer Res., 40: 4485-4488.
Direct Link - Latzka, W.A. and M.N. Sawka, 2000. Hyperhydration and glycerol thermoregulatory effects during exercise in hot climates. Can. J. Appl. Physiol., 25: 536-545.
PubMed - Lee, S.Y., S.H. Lee, K. Akuta, M. Uda and C.W. Song, 2000. Acute histological effects of interstitial hyperthermia on normal rat brain. Int. J. Hyperthermia, 16: 73-83.
Direct Link - Morishima, H.O., B. Glaser, W.H. Niemann and LS. James, 1975. Increased uterine activity and fetal deterioration during maternal hyperthermia. Am. J. Obstet. Gynecol., 121: 531-538.
PubMed - Mirkes, P.E., 1985. Effects of acute exposures of elevated temperatures on rat embryo growth and development in vitro. Teratology, 32: 259-266.
PubMed - Miller, M.W. and M.C. Ziskin, 1989. Biological consequence of hyperthermia. Ultrasound Med. Biol. 15: 707-722.
Direct Link - Milunsky, A., M. Ulcickas, K.J. Rothman, W. Willett, S.S. Jick and H. Jick, 1992. Maternal heat exposure and neural tube defects. JAMA, 268: 882-885.
Direct Link - Bogorad, M.M., S. Pardue, D.D. Mclntire and E.K. Miller, 1994. Cell size and the heat shock response in rat brain. J. Neurochem., 63: 857-867.
PubMed - Mirochnitchenko, O., U. Palnitkar, M. Philbert and M. Inouye, 1995. Thermosensitive phenotype of transgenic mice overproducing human glutathione peroxidases. Proc. Natl. Acad. Sci., 92: 8120-8124.
Direct Link - Malyshev, I. Yu, A.I. Bayda, N.P. Trifonov, L.D. Larionov and E.B. Vanin et al., 2000. Manukhina cross talk between nitric oxide and HSP70 in the antihypotensive effect of adaptation to heat. Physiol. Res., 49: 99-105.
Direct Link - Nilsen, N.Q., 1984. Endothelial changes and microvascular leakage due to hyperthermia in chick embryos. Virchows Arch. B Cell Pathol. Incl. Mol. Pathol., 46: 165-174.
PubMed - Ostberg, J.R., K.C. Kaplan and E.A. Repasky, 2002. Induction of stress proteins in a panel of mouse tissues by fever range whole body hyperthermia. Int. J. Hyperthermia., 18: 552-562.
Direct Link - Pennycuik, P.R., 1965. The effects of acute exposure to high temperatures on prenatal development in the mouse with particular reference to secondary vibrissae. Aust. J. Biol. Sci., 18: 97-113.
PubMed - Poswillo, D., H. Nunnerly D. Sopher and J. Keith, 1974. Hyperthermia as a teratogenic agent. Ann. R. Coll. Surg. Engl., 55: 171-174.
Direct Link - Radmilovich, M., A. Femandez and O. Trujillo-Cenoz, 2003. Environment temperature affects cell proliferation in the spinal cord and brain of juvenile turtles. J. Exp. Biol., 206: 3085-3093.
Direct Link - Shiota, K., 1982. Neural tube defects and maternal hyperthermia in early pregnancy epidemiology in a human embryo population. Am. J. Med. Genet., 12: 281-288.
PubMed - Shih, C.J., M.T. Lin and S.H. Tsai, 1984. Experimental study on the pathogenesis of heat stroke. J. Neurosurg., 60: 1246-1252.
PubMed - Sminia, P., D. Troost and J. Haveman, 1989. Histopathological changes in the spinal cord after 434 MHZ microwave hyperthermia in the cervical region of the rat. Int. J. Hyperthermia, 5: 85-98.
PubMed - Sharma, H.S. and J.C. Navarro, 1990. Brain oedema and cellular changes induced by acute heat stress in young rats. Acta Neurochir. Suppi. (Wien), 51: 383-386.
PubMed - Sharma, H.S., R. Kretzschmar, J.C. Navarro, A. Ermisch, H.J. Ruble and P.K. Dey, 1992. Agerelated pathophysiology of the blood brain barrier in heat stress. Prog. Brain Res., 91: 189-196.
PubMed - Sasaki, J., A. Yamaguchi, Y. Nabeshima, S. Shigemitsu, N. Mesaki and T. Kubo, 1995. Exercise at high temperature causes maternal hyperthermia and fetal anomalies in rats. Teratology, 51: 233-236.
Direct Link - Tveita, T., E. Mortensen, O. Hervroy, K. Ytrehus and H. Refsum, 1991. Hemodynamic and metabolic effects of hypothermia and rewarming. Article Med. Res., 50: 48-52.
Direct Link - Upfold, J.B., M.S.R. Smith and M.J. Edwards, 1989. Quantitative study of the effect of maternal hyperthermia on cell death and proliferation in the guinea pig brain on day 21 of pregnancy. Teratology, 39: 173-179.
Direct Link - Wanner, R.A., M.J. Edwards and R.G. Wright, 1976. The effect of hyperthermia on the neuroepithelium of the 21 day guinea pig foetus histologic and ultrastructural study. J. Pathol., 118: 235-244.
PubMedDirect Link - Wilmore, D.W., Jr. A.D. Mason, D.W. Johnson and Jr. B.A. Pruitt, 1975. Effect of ambient temperature on heat production and heat loss in burn patients. J. Appl. Physiol., 38: 593-597.
Direct Link - Webster, W.S. and M.J. Edwards, 1984. Hyperthermia and the induction of neural tube defects in mice. Teratology, 29: 417-425.
PubMed - Walsh, D.A., N.W. Klein, L.S. Hightower and M.J. Edwards, 1987. Heat shock and thermotolerance during early rat embryo development. Teratology, 36: 181-191.
PubMed - Yang, S.L., S.H. Jing, S.S. Chen, T.J. Chen and R.C. Yang, 1994. The effect of hyperthermic treatment on electroencephalographic recovery after interruption of respiration in rats. Exp. Brain Res., 99: 431-434.
PubMed - Asakura, H., A. Nakai, G.G. Power and T. Araki, 2002. Short-term effects of different thermal conditions during uteroplacental ischaemia on fetal growth of Sprague-Dawley rats. Reprod. Fertil. Dev., 14: 355-361.
CrossRefDirect Link