
O. H. Adekanmbi
Department of Botany and Microbiology, University of Lagos, Lagos, Nigeria
International Journal of Botany
Year: 2009 | Volume: 5 | Issue: 4 | Page No.: 295-300







ABSTRACT
Seven plant species belonging to 3 families were subjected to standard palynological sample preparation in other to provide additional tool in pollen identification. Taxa in this study include the families Asteraceae, Convolvulaceae and Malvaceae. Pollen grains belonging to the family Asteraceae are unique in being characteristically echinate. They can be differentiated from similar pollen in other families by the relatively small size of both the pollen and the spines and the irregular arrangement of the spines. Pollen of Ipomoea aquatica belonging to the family Convolvulaceae has spines like found in Asteraceae but can be distinguished in that those of Convolvulaceae are remarkedly bigger in size. Hibiscus rosasinensis a species in the family Malvaceae share the same echinate characteristics but peculiar in possessing isolated spines, which stand out conspicuously. The pollen grain is differentiated from Convolvulaceae pollen in being larger. Light micrographs, detailed descriptions of the species and where possible Scanning Electron Micrographs are provided. This study is expected to be useful in palaeoecology research, petroleum exploration, honey industry and in forensic investigations.
PDF Abstract XML References Citation
How to cite this article
O. H. Adekanmbi, 2009. Pollen Grains of Asteraceae and Analogous Echinate Grains. International Journal of Botany, 5: 295-300.
DOI: 10.3923/ijb.2009.295.300
URL: https://scialert.net/abstract/?doi=ijb.2009.295.300
DOI: 10.3923/ijb.2009.295.300
URL: https://scialert.net/abstract/?doi=ijb.2009.295.300
INTRODUCTION
Pollen grains have several morphological characters on the exine, which are of diagnostic importance (Edeoga et al., 1998; Edeoga and Ikem, 2002; Mbagwu and Edeoga, 2006; Mbagwu et al., 2008). They are more often than not, species specific. Pollens therefore are used to identify source plant during the analysis of palynological samples in fields such as biostratigraphy, climatology, medicine-alleviation of pollinosis (hay fever-allergenic disease), forensic studies, mellisopalynology, plant evolution, taxonomy and environmental restoration activities (Bryant et al., 1990; Agwu and Uwakwe, 1992; Bayer and Kubitzki, 2003; Perveen et al., 2004; Palazzesi et al., 2007; Adeonipekun and Ige, 2007; Ige, 2009). However, due to high species diversity especially in wet tropics, there occurs the inability of the palynologist to identify or differentiate some pollen forms, which could result in the omission of some important indicator species.
At present, only few descriptions of pollen grains from some Nigeria plants exist, which are scattered over several journals. Existing atlases are in-house work of oil companies and are considered strictly confidential and therefore are not available for public use except the work of Sowunmi (1973, 1995) on the compilation of pollen grains of Nigerian woody plants; the works of Legoux (1978), Takahashi and Ulrich (1989), Jan du Chene et al. (1978), Salard-Cheboldaeff and Dejax (1991) and Biffi and Grignani (1983) which are published.
Realising that a complete coverage of all echinate grains will take several years of teamwork, the decision was made to publish the major differences in the commonly recovered echinate grains belonging to the families Asteraceae, Convolvulaceae and Malvaceae.
MATERIALS AND METHODS
The polliniferous material (some flowers, buds, or even single stamens) collected from University of Lagos and environs was treated with hot water in order to make it soft. Anthers were picked out under a dissecting microscope and placed in a centrifuge tube. Sample preparation was carried out at the Palynological Unit, Botany and Microbiology Department, University of Lagos and repeated at the Jodrel Laboratory, Micromorphology Section, Royal Botanic Garden, Kew, UK following standard sample preparation method which involves acetolysis, repeated wash in distilled water and sieving as outlined by Erdtman (1969). Five plant species belonging to the family Asteraceae and one species each of the families Convolvulaceae and Malvaceae were prepared. Seven species were prepared in all and slides were made.
Photomicrography was carried out under Olympus light microscope with Motic MC2000 (2.0 Megapixel) camera. Specimens are illustrated at either 1000x or 400x. Scanning Electron Micrographs were taken using Hitachi S-4700 Scanning Electron Microscope (SEM) from prepared stubs coated with gold in the Emitech Sputter Coater K550. Pollen grains were described based on their morphological characteristics. Amb or overall grain shape is first defined, followed by shape classes as defined by Erdtman (1969). The shape classes are based on measurements of the polar axis (P) and equatorial diameter (E) and the resulting P/E ratio. Mean dimensions are provided, followed by minima and maxima in parentheses. The figures recorded for the various pollen grains dimensions were generally the average of measurements of 10 non-folded grains (polar and equatorial axis) measured in equatorial view at X400 magnification. Aperture and exine thickness were also measured. The measured values were directly rounded to the nearest micrometer unit (Moore and Webb, 1983; Moore et al., 1991).
RESULTS
Pollen morphological characters of the species studied is presented below by family. The descriptive photomicrographs are compiled in Fig. 1-4. The list of species included in the study, where they were collected from and collection date are showed in Table 1. Morphological characteristics of described palynomorphs are shown in Table 2.
Characteristic of pollen grains:
Asteraceae: Aspilia africana (Pers.) C.D. Adams haemorrhage plant (Fig. 1):
| • | Shape: Rounded triangular amb Prolate grain P/E (1.13) |
| • | Size: Polar axis 14.3 (12.2-15.7 μm); Equatorial axis 12.7 (8.7-14.0 μm) |
| • | Aperture: Tricolporate, aperture not very distinct |
| • | Exine: Tectate, spines wide at the base, with pointed tip, supported by stout bacules |
Chromolaena odorata Linn. (Fig. 1)
| • | Shape: Angular amb, Prolate grain P/E (1.38) |
| • | Size: Polar axis 15.5 (12.2-19.2 μm); Equatorial axis 11.2 (8.7-14.0 μm) |
| • | Aperture: Tricolporate to tetracolporate |
| • | Exine: Tectate. Tectum supported with bacules. Possess supratectal spines |
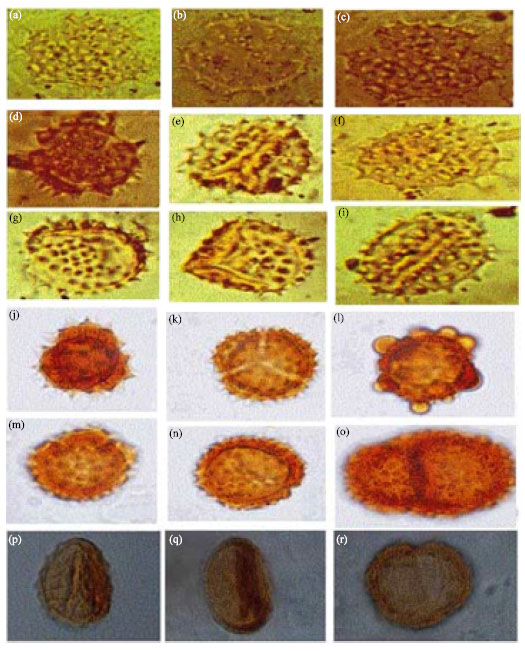
Helianthus sp. (Fig. 2)
| • | Shape: Rounded triangular amb, Prolate grain P/E(1.25) |
| • | Size: Polar axis 15.5 (14.0-17.5 μm); Equatorial axis 12.4 (9.6-14.0 μm) |
| • | Aperture: Tricolporate, aperture not very distinct |
| • | Exine: Tectate, baculate, provided with supratectal spines |
Tridax procumbens Linn. (Fig. 2)
| • | Shape: Rounded square amb, subspheroidal grain, isopolar, radially symmetrical. Prolate grain P/E(1.09) |
| • | Size: Polar axis 18.7 (15.7-21.0 μm); Equatorial axis 17.2 (15.7-19.2μm) |
| • | Aperture: Tri-tetracolporate; colpus long, pore lalongate |
| • | Exine: Tectate, wavy margin, homogenous, spinose; spines wide at the base, with blunt tip, supported by stout bacules |
| Table 1: | Collection information and common names of taxa included in pollen atlas |
 | |
| Table 2: | Morphological characteristics of described palynomorphs |
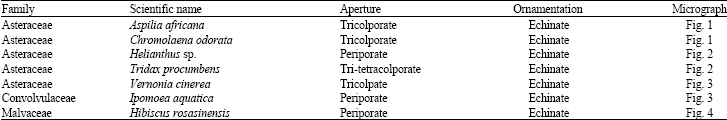 | |
 | |
| Fig. 1: | Asteraceae. (a-i) Aspilia africana (Pers), LM x600 and (j-r) Chromolaena odorata (Linn.) LM x 1000. Note echinate structure of the family Asteraceae |
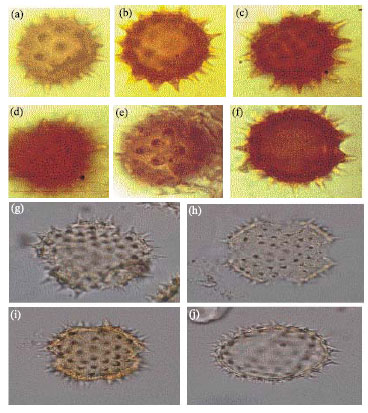
Vernonia cinerea Linn. (Fig. 3)
| • | Shape: Spherical to rounded triangular amb, radially symmetrical |
| • | Size: Maximum dimension 29.5 (27.3-32.5 μm) |
| • | Aperture: Lophate tricolpate; colpus long |
| • | Exine: Wavy margin, exine thickness: 3.5 (2.6-4.1); spinose; spines wide at the base, with sharp tips; spine height: 2.1 (1.6-2.5) μm |
Convolvulaceae: Ipomoea aquatica Forsk (Fig. 3):
| • | Shape: Spherical amb. Grains apolar, radially symmetrical |
| • | Size: 42.4 (39.0-46.0 μm); Equatorial axis 31.2 (27.0-33.0 μm) |
| • | Aperture: Pantotreme, periporate, elliptic and circular, 1.89 μm wide |
| • | Exine: Crassimarginate, differentiated into ectosexine and endosexine, 1.62 μm thick, spinose; spines 4.05 μm long, 2.43 μm wide at the base, blunt at the tip |
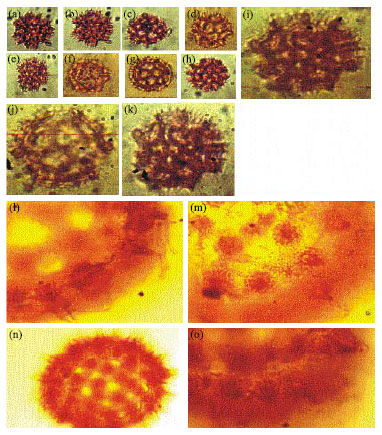
Malvaceae: Hibiscus rosasinensis (Linn.) (Fig. 4):
| • | Shape: Spherical amb. Grains apopolar, radially symmetrical |
| • | Size: Maximum dimension 79.3(72.0-83.5) μm |
 | |
| Fig. 2: | Asteraceae (a-f) showing conspicuous spines and compound aperture in Helianthus sp. (Mart), LM x1000 and (g-j) Tridax procumbens (Linn), LM x1000 |
 | |
| Fig. 3: | Asteraceae (a-k) showing lacunae, ridges and micro spines in Vernonia cinerea, LM, A-H x400. I-K x1000. Convolvulaceae. Ipomoea aquatica (Forsk). (l-o) Showing echinate exine structure in Ipomoea aquatica, LM. L, M, O x1000, N x40 |
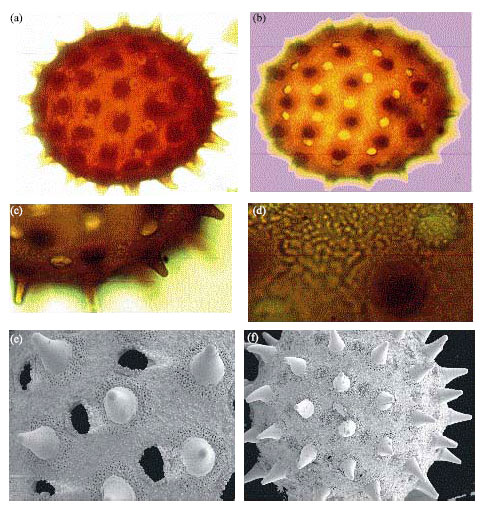 | |
| Fig. 4: | Malvaceae (a-f) showing prominent spines and evenly distributed pori in Hibiscus rosasinensis (Linn.) (a-d) LM, A-B x400, C-D x1000, E, f: SEM E x1000, F x600 |
| • | Aperture: Periporate, poly pantoporate (pores more than 20), uniform in distribution |
| • | Exine: Exine 2.7 μm thick, crassimarginate, spinose, spines 6.75 μm, 2.43 μm wide at the base, blunt apex, sometimes bent; baculate |
DISCUSSION
This study has thrown light on the morphological and differentiating characteristics of the pollens studied. Pollen grains in Asteraceae family are relatively stenopalynous with less variation exhibited in the pollen morphology. Asteraceae pollens are mostly rounded triangular to square amb, sometimes circular in polar view and prolate in equatorial view. Apertures are usually tricolporate to tricolpate sometimes tetracolpate or tetracolporate as found in T. procumbens. Pollen grains in this family are characteristically echinate in ornamentation. This agrees with morphological description of species of Asteraceae treated by Sowunmi (1973). They can be differentiated from similar echinate pollen in other families by the relatively small size of both the pollen and the spines, as well as the irregular arrangement of the spines. Although, Erdtman (1969) reported that in genus Artemisia and a few other anemophilous genera of the Asteraceae, the pollen grains are without obvious spines, Rowley et al. (1981) confirmed the presence of spines which are short and hard to see with light microscopy.
The three families studied have one thing in common and that is the echinate nature of the exine ornamentation. Ipomoea aquatica has spines like found in Asteraceae but remarkedly bigger in size. Aperture in this species is pantotreme or periporate in arrangement.
Hibiscus rosasinensis a species in the family Malvaceae share the same echinate characteristics but peculiar in possessing isolated spines, which stand out conspicuously. The echinate nature of species of Malvaceae was also reported by Perveen and Qaiser (2009). Hibiscus pollen is differentiated from Ipomoea aquatica in being larger in size. This size disparity was noted by Sowunmi (1973), who reported H. tiliaceus of 116.1 μm in size. Slight size disparity is however noted in some pollen grain species reported by various researchers. This may not be unconnected with the fact that pollen grain sizes are affected by acetolysis (Faegri and Iverson, 1989) and a few other chemicals used for grain isolation.
ACKNOWLEDGMENTS
Special thanks to the University of Lagos for the approval of research leave during this study and also for funding. A deep sense of gratitude also goes to The Keeper, Professor Mark Chase of Jodrell Laboratory and Dr. Carol Furness (my host) of The Royal Botanic Gardens, Kew, London for the provision of bench space and equipment. I am greatly indebted to Professor Oluwatoyin Ogundipe for his immense contribution to the successful completion of this research.
REFERENCES
- Biffi, U. and D. Grignani, 1983. Peridinioid of dinoflagellate cysts from the Oligocene of the Niger Delta, Nigeria. Micropaleontology, 29: 126-145.
Direct Link - Bryant, V.M. Jr., D.C. Mildenhall and J.G. Jones, 1990. Forensic palynology in the United States of America. Palynology, 14: 193-208.
Direct Link - Edeoga, H.O., H.N. Ugbo and P.I. Osawe, 1998. Palynomorphology of species of Commelina L. and Senna Tourn. ex Mill. from Nigeria. New Bot., 25: 1-10.
Direct Link - Edeoga, H.O. and I.C. Ikem, 2002. Structural morphology of the pollen grains in three Nigerian Species of Boerhavia L. New Bot., 29: 89-95.
Direct Link - Ige, O.E., 2009. A late tertiary pollen record from Niger Delta, Nigeria. Int. J. Bot., 5: 203-215.
CrossRefDirect Link - Chene, Jan du R.E., M.S. Onyike and M.A. Sowunmi, 1978. Some new Eocene pollen of the Ogwashi-Asaba Formation, South-Eastern Nigeria. Revista Espanola Micropaleontol., 10: 285-322.
Direct Link - Mbagwu, F.N. and H.O. Edeoga, 2006. Palynological studies on some Nigerian species of Vigna savi. J. Biol. Sci., 6: 1122-1125.
CrossRefDirect Link - Mbagwu, F.N., E.G. Chime and C.I.N. Unamba, 2008. Palynological studies on five species of Asteraceae. J. Plant Sci., 3: 126-129.
CrossRefDirect Link - Rowley, J.R., A.O. Dahl and J.S Rowley, 1981. Substructure in exines of Artemisia vulgaris (Asteraceae). Rev. Palaeobot. Palynol., 35: 1-38.
CrossRefDirect Link - Salard-Chebodaeff, M. and J. Dejax, 1991. Evidence of Cretaceous to Recent West Africa intertropical vegetation from continental sediment spore-pollen analysis. J. Afr. Earth Sci., 12: 353-361.
CrossRefDirect Link - Perveen, A., E. Grafstrom and G. El-Ghazaly, 2004. World pollen and spore flora 23. Malvaceae adams. P.p. Subfamilies: Grewioideae, tilioideae, brownlowioideae. Grana, 43: 129-155.
Direct Link