
Mohamed S.M. Abdel-Kareem
Department of Botany, Faculty of Science, Alexandria University, Egypt
International Journal of Botany
Year: 2008 | Volume: 4 | Issue: 4 | Page No.: 361-368







ABSTRACT
A total of 17 Saudi-Arabian algal species related to 4 classes (Cyanophyceae, Rhodophyceae, Phaeophyceae and Chlorophyceae) were screened for the presence of UV-absorbing pigments. Irradiation experiment was conducted on Cladophora glauescens to assess the response of this species to extradoses of UV-B radiation. All the investigated species contain UV-B absorbing pigments. Although it has been suggested that green algae commonly contain low levels of UV-absorbing pigments some investigated green species showed promising quantities of these pigments. The amount of UV-absorbing pigments of Cladophora glauescens varied greatly according to the time as well as the distance of irradiation. Chlorophyll a, Chlorophyll b and carotenoids contents of the investigated species were also assessed.
PDF Abstract XML References Citation
How to cite this article
Mohamed S.M. Abdel-Kareem, 2008. UV-Absorbing Pigments from Some Saudi-Arabian Algal Species. International Journal of Botany, 4: 361-368.
DOI: 10.3923/ijb.2008.361.368
URL: https://scialert.net/abstract/?doi=ijb.2008.361.368
DOI: 10.3923/ijb.2008.361.368
URL: https://scialert.net/abstract/?doi=ijb.2008.361.368
INTRODUCTION
Pigments absorbing ultraviolet light (UV) in algae are thought to be mycosporine-like amino acids (MAAs) with absorbance maxima from 310 to 360 nm (Carreto et al., 1990; Karentz et al., 1991a; Karsten et al., 2005; Volkmann and Gorbushina, 2006; Oren and Gunde-Cimeran, 2007). These pigments are found in all classes of algae (Sivalingam et al., 1974). Some other compounds may participate in protection from UV such as phenolic compounds and alginates, while the linkage between these two compounds preserve the UV absorption capability of phenolic compounds along time (Salgado et al., 2007). Meanwhile, except for very few studies (Garcia-Pichel and Castenholz, 1993; Garcia-Pichel et al., 1993; Post and Larkum, 1993; Sinha et al., 1996; Abdel-Kareem, 1999), the study of UV-absorbing pigments in algae received little attention. However, approximately 400 species of plants have been screened for the sensitivity to UV-B radiation and of these; about two-thirds were found to be sensitive in some parameters (Sullivan and Rozema, 1999).
Though representing only a fraction of total solar electromagnetic spectrum, UV-B (280-315) has disproportional large photobiological effect, due to its absorption by important macromolecules such as proteins and nucleic acids (Giese, 1976). Therefore, it is not surprising that both plant and animal life are greatly affected by UV-B radiation (Sullivan et al., 2003). However, UV radiation has a wide range of effects. They include DNA damage in most organisms (Harm, 1980; Karentz et al., 1991a, b), inhibition of growth and photosynthesis (Worrest, 1982; Post and Larkum, 1993; Larkum and Wood, 1993; Ekelund, 1994; Abdel-Kareem, 1999; Huoyinen et al., 2006), inhibition of photosynthetic primary productivity in both micro-organisms (Smith et al., 1980; Hader and Worrest, 1991; Cullen and Neale, 1994; Prezelin et al., 1994) and higher plants (Tevini and Teramura, 1989; Tivini, 1993), inhibition of nitrogenase activity and heterocyst formation in some cyanobacteria (Sinha et al., 1996), induction of skin cancer (Hwang et al., 2006) and diversity of other responses.
The aim of this research was to screen some Saudi Arabian algal species for the presence of absorbing pigments and to assess the response, including the synthesis of UV-absorbing pigments, of a selected species to extradoses of UV-B radiation.
MATERIALS AND METHODS
Algal materials: Otherwise specified, algal materials were collected from Dammam and Khobar coasts in February 2006. Numerous healthy plants were picked out, washed several times with sea water to remove sand particles, freeze and conveyed to the laboratory. Cladophora glauescens was collected from Sad Nemar at Riyadh, Oedogonim fragile and Mougeotia sp. were collected from a farm at Kharj city (about 100 km South Riyadh), Oscillatoria tenuis was collected from a small rain pond in Riyadh.
Pigments determination: For extraction of UV-absorbing pigments, 1 g fresh samples were extracted for 1 h at room temperature in 75:24:1 methanol:water:HCl (Tevini et al., 1991) by grinding in a mortar and pestle with acid-washed sand. The extract was then centrifuged (3000 g) for 15 min and the supernatant was used for absorbance measurements at 200-400 nm (Pharmacia LKB Ultrospec spectrophotometer). UV-absorbing pigments were expressed as absorbance value at the absorption maxima of the extract in the UV absorption spectra per gram fresh weight and extraction volume 10 mL.
Chlorophylls and carotenoids concentrations were calculated according to Jeffery and Humphrey (1975) and Jaspers (1965), respectively.
Culturing conditions and UV-irradiation: Cladophora glauescens was collected in late May and the thalli were rinsed and placed in shallow trays. Water used for culturing was collected from the sampling site. The trays were placed in an environmental cabinet at 30±2°C with 12 h day/night cycles. On irradiation, the samples were placed in petri-dishes (9 cm diameter) without covers and exposed directly to UV light. The ordinary light was adjusted to 3000 lux. The source of UV-B light was an 8000 mW cm–2 transilluminator supplied by Hoefer pharmacia Biotechnology, USA. Samples were irradiated for 10, 20 and 30 min daily for 5 days at 3 different distances, 10, 30 and 50 cm.
Statistical analysis: All experiments described were conducted independently twice with three replicates each, the mean values given in results representing averages of six assays. Before and after radiation treatments were analyzed using paired t-test and the significance was defined at 5% level of probability.
RESULTS AND DISCUSSION
All the investigated species contain UV-B absorbing pigments (Table 1). The highest absorbance value was recorded by Chondria cornuta (8.38) while the lowest once (1.42) was recorded by Enteromorpha clathrata. Meanwhile, Enteromorpha intestinalis showed the highest value of both chlorophyll a (chl. a, 11.92 mg g–1 fresh wt.) and carotenoids (car., 15.71 mg g–1 fresh wt.). The lowest values of chl. a (0.26 mg g–1 fresh wt.) and car. contents (0.24 mg g–1 fresh wt.) were recorded by Oscillatoria tenuis and Oedogonium fragile, respectively.
While it has been suggested that green algae commonly contain low levels of UV-absorbing pigments (Karentz et al., 1991a), this is clearly not the case for some investigated green species (e.g. Cladophoropsis membranacea, 6.3; Mougeotia sp., 4.47 and Enteromorpha intestinalis, 4.23), which exceed values recorded for some investigated brown, red and blue green species. These results were in consistence with that found in Prasiola crispa growing in Antarctica (Post and Larkum, 1993).
| Table 1: | Screening pigments, chlorophyll a and carotenoids contents of the investigated species |
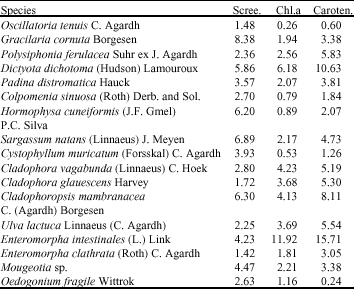 | |
Scree.: Screening pigments, absorbance value at the absorption maxima of the extract in the UV absorption spectra per gram fresh weight and extraction volume 10 mL. Chl.a: Chlorophyll a, mg g–1 fresh weight. Car.: Carotenoids, mg g–1 fresh weight | |
I chose Cladophora glauescens grown in Sad Nemar, Riyadh for the irradiation experiment because of two reasons: First, it contains low content of UV-absorbing pigments which permits good response to UV irradiation and good detection of any variation of the amount of these pigments. Second, this species is economically important, since it is used as bait for fish.
Although some evidence suggested that the concentrations of algal UV-absorbing compounds depend on the degree of exposure to UV light (Scherer et al., 1988; Carreto et al., 1990; Marchant et al., 1991), this is not the case of Cladophora glauescens. Approximately, for most irradiation periods (10, 20 and 30 min) and at most irradiation distances (10, 30 and 50 cm) the amount of UV-absorbing pigments increased gradually to the third day of irradiation and decreased in the fourth and the fifth days (Fig. 1). These increases were significant in more than 50% of treatments (8 out of 15, Table 2), most of these found in the second and the third days of irradiation. This increase in the amount of UV-absorbing pigments could be interpreted to provide an increased protection from UV-B damage. The maximum content of these pigments was achieved after the fourth day of irradiation at 30 cm and for 10 min irradiation time. Algal pigments absorbing UV light thought to be mycosporine-like amino acids (Carreto et al., 1990; Karentz et al.,1991a), functionally comparable to flavonoids of higher plants (Caldwell et al., 1983). The synthesis of these pigments depends not only on the fluence rate but also on the spectrum composition (Carreto et al., 1990). In plants, it is now well established that flavonoids are also be induced by UV-B (Mohle and Wellmann, 1982; Flint et al., 1985; Barnes et al., 1988; Tevini et al., 1991). However, tolerance of some algae such as Scenedesmus sp. and an Enallax sp. to UV-B radiation was correlated with cell wall concentration of sporopollenin (Xiong et al., 1997), a compound found in many algae (Guilford et al., 1988; Xiong et al., 1996, 1997). But this compound not appears to be induced specifically by UV radiation and may have a function in antimicrobial activity (Cockell and Knowland, 1999).
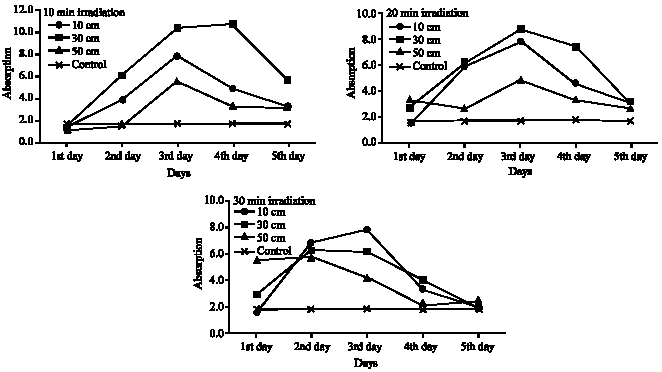 | |
| Fig. 1: | Screening pigments contents of Cladophora glauenscens after UV irradiation at different distances and for different time intervals (expressed as absorbance value at the UV absorption spectra per gram fresh weight and extraction volume 10 mL) |
| Table 2: | Paired samples t-test for screening pigments contents of Cladophora glauescens before and after UV irradiation |
 | |
| *Marked differences are significant at p<0.05, SD: Standard Deviation, SEM: Standard Error Mean | |
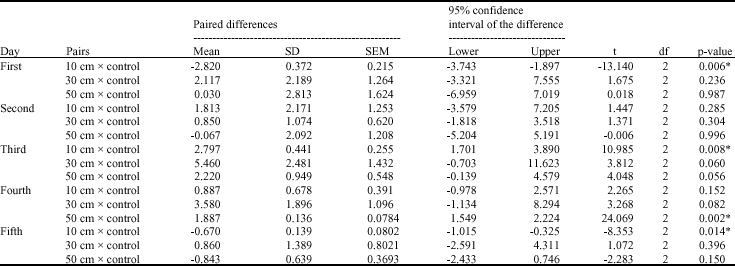
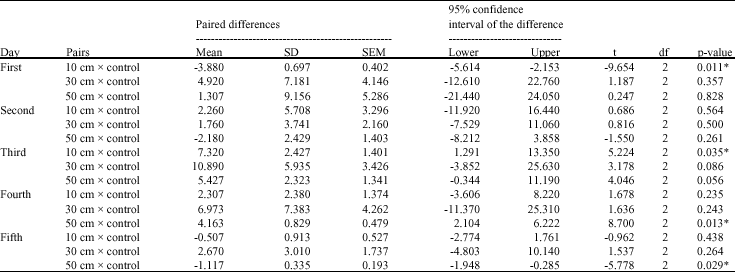
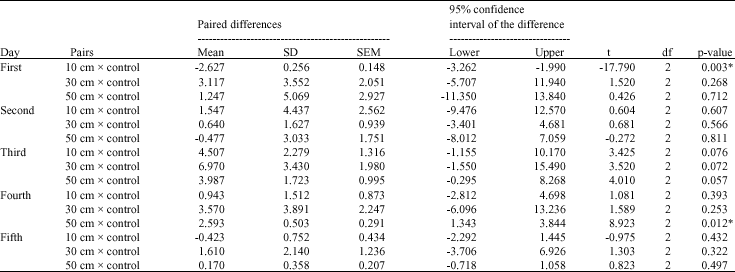
Approximately, the same trend was noticed in the corresponding treatments for the three investigated pigments- chlorophyll a, chlorophyll b and carotenoids (Fig. 2-4). Although there are no significant variations in most treatments on the mean level (Table 3-5), there are great increase in the quantity of these pigments after UV-B irradiation for certain time. In the case of 10 and 20 min irradiation time, the notable increase in the pigments quantity was recorded after the second and the third day of irradiation in most irradiation distances, then decreased quantities were recorded after the fourth and the fifth day of irradiation. At 30 min irradiation time, the pigments contents recorded the maximum increase comparing to control after one day irradiation, except for the 10 cm irradiation distance which recorded the maximum increase at the second or the third day.
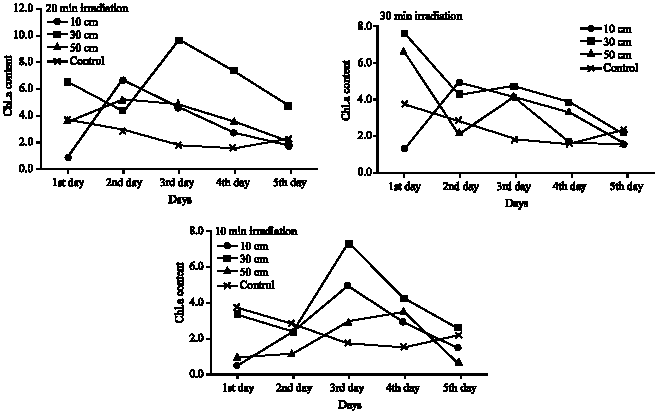 | |
| Fig. 2: | Chlorophyll a contents of Cladophora glauenscens after UV irradiation at different distances and for different time intervals (mg g–1 fresh wt.) |
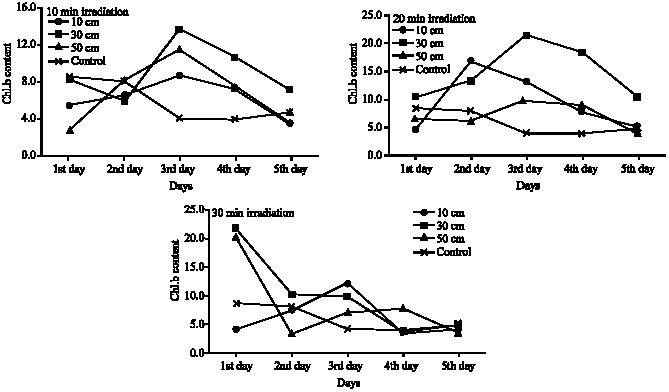 | |
| Fig. 3: | Chlorophyll b contents of Cladophora glauenscens after UV irradiation at different distances and for different time intervals (mg g–1 fresh wt.) |
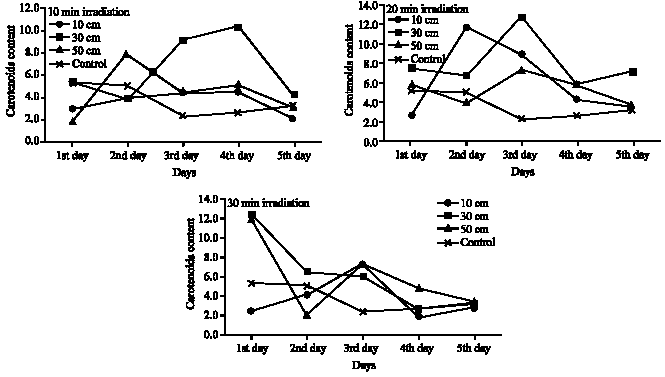 | |
| Fig. 4: | Carotenoids contents of Cladophora glauenscens after UV irradiation at different distances and for different time intervals (mg g–1 fresh wt.) |
| Table 3: | Paired samples t-test for chlorophyll a contents of Cladophora glauescens before and after UV irradiation |
 | |
| *Marked differences are significant at p<0.05, SD: Standard Deviation, SEM: Standard Error Mean | |
The promising feature of these results is the high quantity of carotenoids accumulating in Cladophora glauescens due to UV-B irradiation (Fig. 4). For instance, at 10 min irradiation time and 30 cm irradiation distance, carotenoids contents was about 5 times that of control, while it reached about 6 times of control at the same distance after 20 min irradiation in the fourth day.
For photosynthetic organisms, the protective role of carotenoids against high visible radiation is well known (Siefermann-Harms, 1987; Merzlyak et al., 2005) and a protective role of carotenoids in cyanobacteria against UV-A radiation was reported (Buckley and Houghton, 1976; Paerl, 1984; Ehling-Schulz et al., 1997). Only little is known about the role of carotenoids in photoprotection against UV-B radiation (Middleton and Teramura, 1993; Quesada et al., 1995; Cockell and Knowland, 1999).
| Table 4: | Paired samples t-test for chlorophyll b contents of Cladophora glauescens before and after UV irradiation |
 | |
| *Marked differences are significant at p<0.05, SD: Standard Deviation, SEM: Standard Error Mean | |
| Table 5: | Paired samples t-test for carotenoids contents of Cladophora glauescens before and after UV irradiation |
 | |
| *Marked differences are significant at p<0.05, SD: Standard Deviation, SEM: Standard Error Mean | |
The decrease in the Chl. a, Chl. b and car. contents of Cladophora glauescens with the continuation of UV-B irradiation at the fourth and the fifth day may be correlated with the effect of the UV-B irradiation on the species fitness, although more studies are needed before anything definitive could be postulated in this respect. Nevertheless, very few studies have been dealt with the effect of UV-radiation on algae and clearly further studies are needed. As studies on the response of plants to UV radiation continue to mature beyond the damage state it is likely that we will find that these shorter wavelengths of the ambient solar spectrum (UV-A and UV-B) may have more important roles that was imagined (Sullivan et al., 2003).
CONCLUSION
Promising quantities of UV-B absorbing pigments were found in many studied species. Meanwhile more related studies on such organisms are needed to reach the best conditions of growth that achieved the maximum UV-screening pigment content that may be used in trade and industry scale for the production of sun-screen skin cream. However, UV radiation studies should continue to mature beyond the damage state in both algae and plants to know the important roles of this radiation.
REFERENCES
- Abdel-Kareem, M.S.M., 1999. Influence of ultraviolet radiation on growth, photosynthetic pigments and protein profile of Dunaliella salina Teod. (Chlorophyceae). Egypt. J. Bot., 39: 77-95.
Direct Link - Barnes, P.W., P.W. Jordan, W.G. Gold, S.D. Flint and M.M. Caldwell, 1988. Competition, morphology and canopy structure in wheat (Triticum aestivum L.) and wild oat (Avena fatua L.) exposed to enhanced ultraviolet-B radiation. Funct. Ecol., 2: 319-330.
Direct Link - Buckley, C.E. and J.A. Houghton, 1976. A study of the effects of near UV radiation on pigmentation of the blue-green alga Gloeocapsa alpicola. Arch. Microbiol., 107: 93-97.
CrossRefDirect Link - Caldwell, M.M., R. Robberecht and S.D. Flint, 1983. Internal filters: Prospects for UV-acclimation in higher plants. Physiol. Plant., 58: 445-450.
CrossRefDirect Link - Carreto, J.I., M.O. Carignan, G. Daleo and S.G. De Marco, 1990. Occurrence of mycosporine-like amino acids in the red-tide dinoflagellate Alexandrium excavatum: UV-photoprotective compounds? J. Plant. Res., 12: 909-921.
CrossRefDirect Link - Cockell, C.S. and J. Knowland, 1999. Ultraviolet radiation screening compounds. Biol. Rev., 74: 311-345.
CrossRefDirect Link - Cullen, J.C. and P.J. Neale, 1994. Ultraviolet radiation, ozone depletion and marine photosynthesis. Photo. Res., 39: 303-320.
CrossRefDirect Link - Ehling-Shulz, M., W. Bilger and S. Scherer, 1997. UV-B induced synthesis of photoprotective pigments and extracellular polysaccharides in terrestrial cyanobacterium Nostoc commune. J. Bacteriol., 197: 1940-1945.
Direct Link - Ekelund, N.G.A., 1994. Influence of UV-B radiation on photosynthetic light-response curves, absorption spectra and motility of four phytoplankton species. Physiol. Planta, 91: 696-702.
CrossRefDirect Link - Flint, S.D., P.W. Jordan and M.M. Caldwell, 1985. Plant protective response to enhanced UV-B radiation under field conditions: Leaf optical properties and photosynthesis. Photochem. Photobiol., 41: 95-99.
CrossRefDirect Link - Garcia-Pichel, F. and R.W. Castenholz, 1993. Occurrence of UV-absorbing, mycosporine-like compounds among cyanobacterial isolates and an estimate of their screening capacity. Applied Environ. Microbiol., 59: 163-169.
Direct Link - Garcia-Pichel, F., C. Wingard and R.W. Castenholz, 1993. Evidence regarding the UV sunscreen role of a mycosporine-like compound in the cyanobacterium Gloeocapsa sp. Applied Environ. Microbiol., 59: 170-176.
Direct Link - Guilford, W.J., D.M. Sceneider, J. Labovitz and S.J. Opella, 1988. High-resolution solid-state 13C NMR spectroscopy of sporopollenins from different plant taxa. Plant. Physiol., 86: 134-136.
Direct Link - Hader, D.P. and R.C. Worrest, 1991. Effects of enhanced solar ultraviolet radiation on aquatic ecosystem. Photochem. Photobiol., 53: 717-725.
CrossRefDirect Link - Huovinen, P., I. Gomez and C. Lovengreen, 2006. A five year study of solar ultraviolet radiation in Southern Chile (39°S): Potential impact on physiology of costal marine algae. Photochem. Photobiol., 82: 515-522.
Direct Link - Hwang, H., T. Chen, R.G. Nines, H. Shin and G.D. Stoner, 2006. Photochemoprevention of UVB-induced skin carcinogenesis in SKH-1 mice by brown algae polyphenols. Int. J. Cancer, 119: 2742-2749.
Direct Link - Jaspers, E.M.W., 1965. Pigmentation of tobacco crown-gall tissues cultured in vitro in dependence of the composition medium. Physiol. Plant., 18: 933-940.
CrossRefDirect Link - Jeffery, S.W. and G.F. Humphrey, 1975. New spectroscopic equation for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton. Biochem. Physiol. Pflanzen., 167: 191-194.
Direct Link - Karentz, D., J.E. Cleaver and D.L. Mitchell, 1991. Cell survival characteristics and molecular responses of Antarctic phytoplankton to ultraviolet-B radiation. J. Phycol., 27: 326-341.
CrossRefDirect Link - Karentz, D., S. McEuen, M.C. Land and W.C. Land, 1991. Survey of mycosporine-like amino acid compounds in Antarctic marine organisms: Potential protection from ultraviolet exposure. Mar. Biol., 108: 157-166.
CrossRefDirect Link - Karsten, U., T. Friedl, R. Schumann, K. Hoyer and S. Lembcke, 2005. Mycosporine-like amino acids and phylogenies in green algae: Prasiola and its relatives from trebouxiphyceae (Chlorophyta). J. Phycol., 41: 557-566.
Direct Link - Larkum, A.W. and W.F. Wood, 1993. The effect of UV-B radiation on photosynthesis and respiration of phytoplankton, benthic macroalgae and seagrasses. Photo. Res., 36: 17-25.
CrossRefDirect Link - Marchant, H.J., A.T. Davidson and G.J. Kelly, 1991. UV-B protecting compounds in marine alga Phaeocystis pouchetii from Antarctica. Mar. Biol., 109: 391-395.
CrossRefDirect Link - Merzlyak, M.N., A.E. Solovchenko, A.I. Smagin and A.A. Gitelson, 2005. Apple flavonols during fruit adaptation to solar radiation: Spectral features and technique for non-destructive assessment. J. Plant Physiol., 162: 151-160.
CrossRefPubMedDirect Link - Middleton, E.M. and A.H. Teramura, 1993. The role of flavonol glycosides and carotenoids in protecting soybean from ultraviolet-B damage. Plant. Physiol., 103: 741-752.
CrossRefDirect Link - Mohle, B. and E. Wellmann, 1982. Induction of phenylpropanoid compounds by UV-B irradiation in roots of seedlings and cell cultures from Dill (Anethum graveolens L.). Plant Cell Rep., 1: 183-185.
CrossRefDirect Link - Oren, A. and N. Gunde-Cimeran, 2007. Mycosporine and mycosporine-like amino acids: UV protectants or multipurpose secondary metabolites. FEMS. Microbiol. Lett., 269: 1-10.
Direct Link - Paerl, H.W., 1984. Cyanobacterial carotenoids: Their roles in maintaining optimal photosynthetic production among aquatic bloom forming genera. Oecolagia, 61: 143-149.
Direct Link - Post, A. and A.W. Larkum, 1993. UV-absorbing pigments, photosynthesis and UV exposure in Antarctica: Comparison of terrestrial and marine algae. Aquat. Bot., 45: 231-243.
Direct Link - Quesada, A., J.L. Mouget and W.F. Vincent, 1995. Growth of Antarctic cyanobacteria under ultraviolet radiation: UV-B inhibition. J. Phycol., 31: 424-428.
Direct Link - Salgado, L.T., R. Tomazetto, L.P. Cinelli, M. Farina and G.M.A. Filho, 2007. The influence of brown algae alginates on phenolic compounds capability of ultraviolet radiation absorption in vitro. Braz. J. Oceanol., 55: 145-154.
CrossRefDirect Link - Scherer, S., T.W. Chen and P. Boger, 1988. A new UV-A/B protecting pigment in the terrestrial cyanobacterium Nostoc commune. Plant. Physiol., 88: 1055-1057.
Direct Link - Siefermann-Harms, D., 1987. The light-harvesting and protective functions of carotenoids in photosynthetic membranes. Physiol. Plant., 69: 561-568.
CrossRefDirect Link - Sinha, R.P., N. Singh, A. Kumar, H.D. Kumar, M. Hader and D.P. Hader, 1996. Effects of UV irradiation on certain physiological and biochemical processes in cyanobacteria. J. Photochem. Photobiol. B: Biol., 32: 107-113.
CrossRef - Sivalingam, P.M., T. Ikawa, Y. Yokohama and K. Nisizawa, 1974. Distribution of a 334 UV-absorbing-substance in algae, with special regard of its possible physiological roles. Bot. Mar., 17: 23-29.
Direct Link - Smith, R.C., K.S. Baker, O. Holm-Hansen and R. Olsen, 1980. Photoinhibition of photosynthesis in natural waters. Photochem. Photobiol., 31: 585-592.
CrossRefDirect Link - Sullivan, J.H. and J. Rozema, 1999. UV-B Effects on Terrestrial Plant Growth and Photosynthesis. In: The Effects of Enhanced UV-B Radiation on Terrestrial Ecosystems, Stratospheric, O.D. and J. Rozema, (Eds.). Backhuys Publishers, Leiden pp: 39-57.
Direct Link - Sullivan, J.H., D.C. Gitz, M.S. Peek and A.J. McElrone, 2003. Response of three Eastern tree species to supplemental UV-B radiation: Leaf chemistry and gas exchange. Agric. For. Meteorol., 120: 219-228.
Direct Link - Tevini, M. and A.H. Teramura, 1989. UV-B effects on terrestrial plants. Photochem. Photobiol., 50: 479-487.
CrossRefDirect Link - Tevini, M., J. Braun and G. Fieser, 1991. The protective function of the epidermal layer of rye seedlings against ultraviolet-B radiation. Photochem. Photobiol., 53: 329-333.
CrossRefDirect Link - Volkmann, M. and A. Gorbushina, 2006. A broadly applicable method for extraction and characterization of mycosporines and mycosporine-like amino acids of terrestrial, marine and freshwater origin. FEMS. Microbiol. Lett., 255: 286-295.
Direct Link - Xiong, F., F. Lederer, J. Lukavsky and L. Nedbal, 1996. Screening of freshwater algae (Chlorophyta, Chromophyta) for ultraviolet-B sensitivity of the photosynthetic apparatus. J. Plant Physiol., 148: 42-48.
Direct Link - Xiong, F., J. Komenda, J. Kopecky and L. Nedpal, 1997. Strategies of ultraviolet-B protection in microscopic algae. Physiol. Plant, 100: 378-388.
Direct Link