
C. Cosmas Ogbu
Department of Animal Science, University of Nigeria, Nsukka, Nigeria
Asian Journal of Poultry Science
Year: 2012 | Volume: 6 | Issue: 3 | Page No.: 89-96







ABSTRACT
The study was carried out to estimate phenotypic response in body weight for three generations using mass selection and additive genetic heritability of body weight at 12, 16, 20 and 39 weeks of age (WOA) in males of the Nigerian indigenous chicken. The study took place at the Department of Animal Science, University of Nigeria, Nsukka. Body weight at 39 WOA was the selection criterion. Results showed that body weight increased from 1372.66±16.46 g in G0 generation to 1656.58±27.45 and 1768.75±33.15 g in G1 and G2 generations, respectively. These implied phenotypic responses of 321.09, 373.42 and 231.25 g and cumulative responses of 321.09, 694.51 and 925.76 g for G0, G1 and G2 generations, respectively. There was moderate to high additive genetic heritability for body weight up to 20 WOA (range, 0.24±0.27 to 0.59±0.45) and low to moderate heritability for body weight at 39 WOA (range, 0.13±0.49 to 0.25±0.31). Selection intensity was 2.11 in G0, 1.75 in G1 and 1.16 in G2 generation. Total variance was 198.87 in the last generation indicating the presence of usable variation for further selection. From the results, it was concluded that growth performance in the indigenous chicken can be improved through mass selection especially within the growing period of 12 to 20 WOA.
PDF Abstract XML References Citation
Received: March 22, 2012;
Accepted: September 08, 2012;
Published: October 12, 2012
How to cite this article
C. Cosmas Ogbu, 2012. Phenotypic Response to Mass Selection in the Nigerian Indigenous Chickens. Asian Journal of Poultry Science, 6: 89-96.
DOI: 10.3923/ajpsaj.2012.89.96
URL: https://scialert.net/abstract/?doi=ajpsaj.2012.89.96
DOI: 10.3923/ajpsaj.2012.89.96
URL: https://scialert.net/abstract/?doi=ajpsaj.2012.89.96
INTRODUCTION
The genetic progress in performance traits such as growth and egg production in the exotic breeds has been tremendous. Meat type birds (broilers) attain live weights of 1.5 to 2.0 kg in about 6 to 8 weeks with feed conversion ratio of about 2:1 (Pym, 2010). Commercial egg stocks average 300+ eggs per annum under optimal conditions. Continuous selection is generally believed to be responsible for these improvements. Conversely, the Nigerian indigenous chicken has remained unimproved. Schemes suggested for the improvement of Indigenous Chickens (IC) in developing countries include: crossbreeding with improved exotic breeds, selection within the local stocks and a combination of these methods (Safalaoh, 2000; Kperegbeyi at al, 2009; Pym, 2010). Crossbreeding using exotic breeds has received the greatest attention in most developing countries (Safalaoh, 2000) because of the belief that crossing the indigenous chickens with the exotic will yield improved indigenous stocks. This scheme however failed (Besbes, 2008; Dana et al., 2010). No indigenous breed of commercial appeal exists in any of these countries. A number of reasons have been advanced for the dismal failure of this scheme among which was the lack of sustained supply of exotic replacement stocks (Safalaoh, 2000). This reason indicates that the products of the mating between the IC and exotic breeds were not meant to be breeders (grand parent stocks) but commercial or terminal products.
To develop indigenous poultry breeds capable of sustaining the indigenous poultry industry, a deliberate and sustained effort must be made to select from among the IC individuals having good genetic potentials for economically important traits (Safalaoh, 2000; Ogbu and Nwosu, 2011). These individuals can then be bred, further selected and established as specialised lines which could subsequently be used in complementary crossings involving proven exotic and indigenous chicken lines (Safalaoh, 2000). In spite of current emphasis on molecular markers and marker assisted selection, conventional selection techniques such as phenotypic selection are still considered very effective selection strategies (Robbins and Staub, 2005; Davis and Berzonsky, 2006) and will continue to be employed for the genetic improvement of livestock species especially in developing countries lacking requisite technology and technical competence in molecular selection techniques. The present study determines the phenotypic response to mass selection for body weight at 39 weeks of age (BWT39) and additive genetic heritability for body weight at 12, 16, 20 and 39 WOA in males of the Nigerian Heavy Local Chicken Ecotype (NHLCE).
MATERIALS AND METHODS
The study was carried out between 2005 and 2008 and covered three generations (G0, G1 and G2 generations). Five hundred 1-day chicks (sexes combined) which were F1 progenies of 5 sire families established from a random breeding population of NHLCE were used for the study. These birds formed the G0 generation with which the process of selection was commenced. The birds were wing-banded, brooded and reared according to sire families. They were fed on chicks mash (20% CP and 2800 kcal ME kg-1) from 0-8 weeks and growers mash (16% CP and 2670 kcal ME kg-1) from 8 to 20 weeks of age. Feed was provided ad libitum to the birds at the starter phase (0-8 weeks). During the grower phase (8-20 WOA) and mature phase (above 20 WOA) birds received feed above their maintenance requirement as determined previously by Ogbu and Omeje (2011) namely: 78 g bird day-1 (8-12 WOA), 88 g bird day-1 (12-16 WOA), 98, 108, 118 and 125 g bird day-1 between 16-20, 20-24, 24-30 and 30-39 WOA, respectively. The birds had unrestricted access to cool and clean water throughout the period of the experiment. At 12 WOA, males were separated from females. At 16 WOA, they were moved into individual cages to prevent cannibalism. Routine vaccination and other health management practices were kept optimal to allow each bird express its full genetic potentials.
Data collection and analysis: Birds were weighed every 4 weeks from 1 day-20 WOA in each generation to obtain their body weight (BWT) values. Final BWT for males was taken at 39 WOA at which their female counterparts were selected based on body weight and egg production performance. The values for BWT39 were used to rank males for mass selection. Selected males from the G0 generation were used as sires to produce the G1 generation which in turn yielded sires for the G2 generation. Data on BWT39 were subjected to analysis of variance (ANOVA) in a completely randomised design using the SPSS computer package. The statistical model is:
Xijk = μ+Si+Gj+eijk |
where, Xijk is observation on the kth progeny of the ith sire in the jth generation; μ is overall mean, Si is random effect of sire; Gj is fixed effect of jth generation and eijk is residual effects assumed to be independent and normally distributed with zero mean and variance of error. Significant means were separated using the Duncan New Multiple Range Test option of SPSS. Population mean comparison was done using the t-test option of SPSS computer programme. From sire component of variance, the additive genetic heritability of BWT from 12 to 20 and 39 weeks for each generation was calculated using the relationship:
| Where: | ||
| = | Sire variance component | |
| σ2w | = | Residual variance |
Phenotypic response (selection differential, ΔS): This was calculated as the mean difference between the selected and the entire population before selection. Thus,
where, ΔS is selection differential (phenotypic response); ![]() is mean of selected group and
is mean of selected group and ![]() is mean of whole population (mean of population before selection).
is mean of whole population (mean of population before selection).
Selection intensity: This was calculated using the expression:
where, ΔS is selection differential and σp is phenotypic standard deviation.
Cumulative selection differential: This is the sum of the selection differential in the current generation and the selection differential(s) in the previous generation(s).
That is:
RESULTS
Table 1 presents the descriptive statistics (mean±SE) for BWT performance of the experimental birds from 0-8 weeks (sexes combined) and from 12-20 weeks of age (males) for G0, G1 and G2 generations. The table shows significant (p≤0.05) differences between generations across the age periods. Body weight (BWT) at 0, 4 and 16 WOA was highest in G2 generation and least in G0 generation.
| Table 1: | Growth performance of NHLCE from 0-8 weeks (sexes combined) and 12-20 weeks (males) |
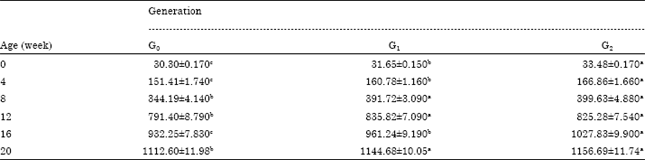 | |
| Values (Mean±SE) in the same row with different superscripts are significantly different at p≤0.05, G: Generations | |
| Table 2: | Population and generation comparison for BWT39 in a population of males of NHLCE under selection |
 | |
| Values (Mean±SE) in the same column with different superscripts are significantly different | |
| Table 3: | Selection intensity, phenotypic variance and phenotypic response (selection differential) for BWT39 in males of NHLCE under selection |
 | |
Mean values for BWT at 8, 12 and 20 WOA were similar in G1 and G2 generations but higher than the values in G0 generation. Across age periods, standard error of mean (SEM) increased while across generations values of SEM were generally similar.
The between population mean comparison (whole vs. selected) across generations and generation mean comparison for BWT39 across populations are presented in Table 2. From the table, selected populations were consistently superior (p≤0.01) to their whole (unselected) counterparts in BWT39 in each generation. Among generations, mean value for whole populations varied significantly (p≤0.01) with an upward trend. Thus BWT39 was highest in G2 generation, followed by G1 generation and least in G0 generation. For the selected population, mean BWT39 were similar in G1 and G2 generations but significantly (p≤0.01) surpassed that in G0 generation.
Table 3 presents the applied selection intensity (i), phenotypic standard deviation (σp), selection differential (Δs) and cumulative selection differential (cumΔs) for BWT39 across the generations of selection. The table shows that selection intensity ranged from 2.11 to 1.16 with a downward trend from G0 generation. Phenotypic standard deviation varied across generations but generally increased in value from the initial value of 152.05 in Go generation to 212.63 and 198.87 in G1 and G2 generations, respectively.
| Table 4: | Estimate of additive genetic heritability |
 | |
Table 3 also shows positive phenotypic selection response (positive selection differential) for BWT39 across the generations. Cumulative selection differential was hence positive and increased over generations from 321.09 g in G0 to its final value of 925.76 g in G2 generation.
Table 4 displays the estimate of additive genetic heritability for BWT at various age periods for unselected (whole) populations of the three generations. Heritability values were generally high (≥0.39±0.26) across generations at 12 weeks of age, moderate to high (0.24±0.27-0.51±0.31) at 16 and 20 weeks of age and low to moderate (0.13±0.49-0.25±0.31) at 39 weeks of age. For all age periods except 12th week, heritability values showed a downward trend from G0 to G2 generation while within each generation heritability values were least for BWT39.
DISCUSSION
The BWT performances as obtained in G0 generation at various ages (0-8 weeks for sexes combined and 12-20 weeks for males) were higher than those reported by Oluyemi (1979a), Nwosu and Asuquo (1985), Adetayo and Babafunso (2001) and Okpeku et al. (2003) for indigenous chickens from the derived savannah, Guinea savannah and rainforest zones of South-Eastern and Western Nigeria. These birds containing variable numbers of the heavy and light ecotypes were expectedly lower in BWT than the ‘pure’ heavy ecotype chickens that made up the G0 generation in the present study. The results were thus very similar to values reported for heavy ecotype local chickens by Momoh et al. (2010). The superiority of chickens belonging to G1 and G2 generations over those of G0 generation in BWT across the entire age periods was expected. G1 birds being progenies of selected parents from G0 generation manifested realized selection responses in body weight due to selection in the G0 generation of males superior in BWT39. These positive correlated responses, manifested as increased body weight at these age periods. Birds belonging to G2 generation also manifested positive correlated responses to selection at 39 weeks although to a lesser degree hence they were superior to their G1 counterparts only at 0, 4 and 16 weeks of age. The increasing values observed for SEM indicate increasing values of phenotypic variance with increases in body weight. It does appear that means and variances for growth are correlated. Gowe and Fairfull (1985) as well as Fairfull and Gowe (1990) had shown that environmental variation and thus phenotypic variation, experiences relatively large increases with age. The between generation similarity among SEM for BWT values at each age period indicate homogeneity of variances across generations. Generally, the generational differences in BWT within the self accelerating growth phase (0-20 weeks) both for sexes combined and for males were not very striking. The selection of females based on BWT (body weight at first egg) and egg production (egg number and egg weight) may have reduced the gain in body weight to below what it would have been had females been selected solely on BWT as their male counterparts. The marginal increases in BWT values from 0-20 weeks in G1 and G2 generations therefore, represent correlated responses to selection for BWT39 across the generations. Furthermore the selection of males based on BWT39 may have affected the selection response since much of the genetic potentials for growth may have been exhausted and additive genetic differences (variation) among individuals in each generation may have waned. Selection for body weight improvement in chickens would be most effective at juvenile age periods (4 to 8 weeks) in meat type birds (Marks, 1983) and between 4-20 weeks in local and egg-type chickens (Oluyemi, 1979b) because of higher additive genetic heritability-a reflection of additive genetic variance-at these periods. Marks (1983) reported an apparent plateau of body weight at 40 weeks of age in a population of meat type birds selected for 4 and 8 weeks body weights due to exhaustion of genetic variability. On the other hand, the results obtained in the present study indicate that growth performance in males can be improved (though marginally) while selecting females for egg production.
The highly significantly heavier BWT39 of selected sires compared to the unselected population was in response to the selection of superior individuals (phenotypes and genotypes) within each generation to become parents of the next generation. The consistently significant increases in BWT39 from G0 to G2 generations observed for the whole population indicate positive response to selection for BWT39. Marks (1983) reported such linear increases in BWT in a broiler population selected for 8 weeks BWT. Oluyemi (1979b) reported similar trends in a population of Nigerian indigenous fowl selected based on 12 weeks BWT. Also Nwagu et al. (2007) reported increases in 40 weeks BWT (in the male line) over four (4) out of five (5) generations of selection in a population of Rhode Island chickens selected for part period egg production.
The reported values of selection intensity show that on the average a selection intensity of 1.67 standard deviations was applied across the generations for the selection of sires. The low selection pressure after G0 generation resulted from the declining additive genetic variability among individuals of G1 and G2 generations in BWT39 as inferred from the declining values for additive genetic heritability. The selection intensity values reported in the present study are higher than the values reported by Marks (1983) who obtained average selection intensities of 0.70 and 0.91 in two populations of broilers selected for 8 weeks BWT in male and female lines for four (4) generations. The values are, however, in high accord with selection intensities calculated from data reported by Oluyemi (1979b) which ranged from 0.996 to 2.40 over seven (7) generations of selection for 12 weeks BWT in the Nigerian indigenous fowl. The phenotypic standard deviation indicate persistence of usable variation across the generations. The positive selection differentials (positive phenotypic response) observed, followed from the superiority of selected population over the unselected (whole) population in BWT39. The cumulative selection differentials were correspondingly positive and increased in value across generations.
The heritability estimates obtained in the present study for BWT from 12 to 20 weeks fall within the range commonly reported in literature (Omeje and Nwosu, 1983; Nwosu and Asuquo, 1985; Fairfull and Gowe, 1990; Ogbu and Nwosu, 2010). The moderate to high heritability values for BWT in the accelerating phase of growth (≤20 weeks) indicate the availability of high additive genetic variance in the heavy local chicken ecotype and that BWT at these age periods will respond favourably to selection. The low additive genetic heritability obtained in G2 for BWT39 indicate reduced additive genetic variance in BWT and that selection on BWT39 may not yield significant genetic progress for improved growth rate in the NHLCE as was observed.
CONCLUSION
Conventional selection strategies have been mostly responsible for the improved performance in virtually all exotic breeds of animals. Phenotypic selection response measures the superiority of selected individuals over the population from which they were selected and gives indication of expected genetic gain in the trait concerned. Response to selection relies heavily on the additive genetic variation among individuals of a population in the trait considered. This variation is measured by the value of additive genetic heritability. The positive phenotypic response for body weight obtained in the present study as well as the low to high heritability values indicate that mass selection for body weight in the indigenous chicken population will lead to genetic improvement in this trait especially within the growing age periods (12 to 20 weeks).
REFERENCES
- Adetayo, A.S. and S.E. Babafunso, 2001. Comparison of the performance of Nigerian indigenous chicken from three agro-ecological zones. Livest. Res. Rural Dev., 3: 1-6.
Direct Link - Dana, N., L.H. van der Waaij, T. Dessie and J.A.M. van Arendonk, 2010. Production objectives and trait preferences of village poultry producers of Ethiopia: Implications for designing breeding schemes utilizing indigenous chicken genetic resources. Trop. Anim. Health Prod., 42: 1519-1529.
CrossRefDirect Link - Kperegbeyi, J.I., J.A. Meye and E. Ogboi, 2009. Local chicken production: Strategy of house-hold poultry development in coastal regions of Niger Delta. Nigeria. Afr. J. General Agric., 5: 17-20.
Direct Link - Davis, J., W.A. Berzonsky and G.D. Leachm, 2006. A comparison of marker-assisted and phenotypic selection for high grain protein content in spring wheat. Euphytica, 152: 117-134.
CrossRef - Marks, H.L., 1983. Selection for body weight at eight weeks of age in Dwarf and Normal meat-type chickens originating from a common control population Background. Poult. Sci., 62: 227-234.
PubMed - Momoh, O.M., C.C. Nwosu and I.A. Adeyinka, 2010. Comparative evaluation of two nigerian local chicken ecotypes and their crosses for growth traits. Int. J. Poult. Sci., 9: 738-743.
CrossRefDirect Link - Nwagu, B.I., S.A.S. Olorunju, O.O. Oni, L.O. Eduvie, I.A. Adeyinka, A.A. Sekoni and F.O. Abeke, 2007. Response of egg number to selection in Rhode Island chickens selected for part period egg production. Int. J. Poult. Sci., 6: 18-22.
CrossRefDirect Link - Ogbu, C.C. and C.C. Nwosu, 2010. Characterization of the Nigerian indigenous chickens reared in Ebonyi State in the humid tropics 2: Variance components and additive genetic heritability of body weight, body weight gain and feed: Gain ratio. Proceedings of the 15th Annual Conference of Nigerian Society for Animal Production, September 13-16, 2010, Nigeria, pp: 54-57.
- Ogbu, C.C. and C.C. Nwosu, 2011. Phenotypic response to three generations of mass selection for body weight at 39 weeks in males of the Nigerian Heavy Local Chicken Ecotype (NHLCE). Proceedings of the 36th Annual Conference of Nigerian Society for Animal Production, March 13-16, 2011, University of Abuja, Nigeria, pp: 84-86.
- Ogbu, C.C and S.S.I. Omeje, 2011. Within population variation in performance traits in the Nigerian Indigenous Chicken (NIC). Int. J. Sci. Nat., 2: 192-197.
Direct Link - Safalaoh, A.C.L., 2000. Strenghts and Weaknesses of Small Holder Poultry Improvement Programmes in Malawi. In: Sustainable Animal Agriculture and Crisis Mitigation in Livestock Development Systems in Southern Africa, Banda, J.W., M.G.G. Chagunda, L.A. Kamwanja, R.K.D. Phoya and A.C.L. Safalaoh (Eds.). Malawi Institute of Management, Lilongwe, Malawi.